

Summary
Bu yazıda veziküller, transkripsiyon ve çeviri makine kapsülleme ve protein üretiminin izlenmesi hazırlanması için basit bir yöntem tarif eder. Elde edilen hücre-içermeyen sistemler gittikçe daha karmaşık hücresel taklit oluşturmak için bir başlangıç noktası olarak kullanılabilir.
Abstract
Bireysel moleküllerden moleküllerin sistemlerine ilgi vardiya olarak, laboratuvar giderek artan sayıda daha iyi hücresel yaşamın karmaşıklığı temsil hücresel taklit aşağıdan yukarıya doğru inşa etmek için çalışmışlardır. Su-içinde-yağ emülsiyonları, mikroakışkan cihazlar, ve kabarcıklar dahil olmak üzere, işleme bölümlere ayrılmış hücresel taklit oluşturmak için alınabilir yolları bir dizi vardır, bugüne kadar. Mevcut seçeneklerden her biri belirli avantajları ve dezavantajları vardır. Örneğin, su-içinde-yağ emülsiyonları yüksek bir kapsülleme etkinliği vermek değil de canlı hücre geçirgenlik bariyeri taklit ederler. Bu tarifnamede tarif edilen metotların en önemli avantajı, bunların hepsi kolay ve uygulamak için ucuz olmasıdır. Transkripsiyon-çeviri makine böyle bir santrifüj evaporatör ve bir ekstruder olarak ortak enstrümantasyon, sömüren bir süreç fosfolipid veziküller içinde kapsüllenir. Reaksiyonları floresans spektroskopisi ile izlenmektedir. Protokols rekombinant protein ekspresyonu, hücre taklit yapımı, hücresel yaşam için minimum gereksinimleri arama, ya da genetik devre montajı için adapte edilebilir.
Introduction
Hücre içermeyen, in vitro transkripsiyon yapıldı reaksiyonlar ve sentetik lipidlerden veziküllerin üretilmesi yeni bir şey değildir. Ancak, taklit bir cep içine iki birleştirerek challenging1-6 önemli ölçüde daha fazladır. E. T7 RNA polimerazı ile veya coli hücre ekstreleri transkripsiyon yapıldı makine 7,8 kaynağı olarak kullanılabilir. Protein ekspresyonu ve katlanmasını kolaylaştırmak ek hücre bileşenlerinin varlığı hücre ekstreleri yarar. Alternatif olarak, tek tek saflaştırılmış RNA ve protein moleküllerinin bir karışımı, saf bir sistem, yani 9 İntraveziküler protein sentezi 4,10-14 aracılık etmek için kullanılabilir. Saf bir sistem tam olarak tanımlanmış hücresel taklit yapımı için izin verir ve hücre ekstreleri bulunan nükleaz aktivitesi muzdarip değildir. Pratik olarak, bu sayede düşük kapsülleme etkinliği 11 ile süreçleri kolaylaştıran, daha az DNA şablonu gerekli olduğu anlamına gelir 16-18 bildirilmiştir.
Transkripsiyon-çeviri reaksiyonları izlemek için en basit yolu genetik olarak kodlanmış unsurların floresan veya lüminesans ölçmektir. In vitro reaksiyonlar sıklıkla radyo-etiketleme ile ölçülür, ancak tipik olarak, ateşböceği lusiferaz 19 veya GFP kullanılır. Floresan algılama ayrıca böylece biyolojik benzeri süreçlerin stokastik doğası içine bazı bilgiler sunan, sitometri tabanlı yöntemlerle veziküller 20,21 nüfusu en izlenmesi için izin verir. Bu izleme yöntemleri i ile uyumlu floresan proteinleri bir toplama dahil tasarım kurallarının küçük bir set ve hangi gelen oluşturmak için parçaları bir kütüphane, tanımlamak için kullanılmıştırN in vitro transkripsiyon ve çeviri-22, ifade 22 genetik kuruluşun etkisi, sigma faktörleri 16 aktivitesini ve transkripsiyonel terminatörler 23 verim. Bununla birlikte, çok in vitro öngörülebilir inşa yeteneği, genetik olarak kodlanmış cihazlar artırmak için yapılması gereken orada kalır.
Veziküller yapmak için pek çok yöntem vardır. En sık kullanılan yöntem, sulu solution24 tabanda tarafından izlenen bir cam yüzeyi üzerinde ince bir lipid filmi üretimi bağlıdır. Sulu çözelti transkripsiyon yapıldı makine içeriyorsa, örneğin, daha sonra veziküllerin bir kısmını oluşturduğu protein üretimi için gerekli olan bileşenler içerebilir. Bununla birlikte, bu yöntemlerin kapsülleme etkinliği veziküllerin sadece küçük bir yüzdesi aktif olduğunu, yani düşüktür. Alternatif yöntemlerin çoğu, çok daha yüksek bir kapsülleme eff ile karakterizeiciency veziküller su-içinde-yağ emülsiyonu damlacıklarının dönüşüm kullanılması. Bu tür yöntemler ileride olağan olması muhtemeldir olsa da, şu anda bu yöntemlerin özel ekipman ihtiyacı muzdarip ve değişmiş membran bileşimler ile 25 veziküller verir. Vezikül yöntemlere su-içinde-yağ bir avantaj membran lameller kontrol etmek için potansiyeldir. Burada tarif edilen yöntem, ek bir homojenleştirme adımı da dahil olmak üzere hafif değişiklikler ile Yomo laboratuar 11 tarafından tarif edilen ince bir lipid filmi protokolüne dayanmaktadır. Bu yöntem, kolay, ucuz, ve de transkripsiyon yapıldı makine kapsüllenmesi için uygundur sağlam veziküller verir.
Protocol
1. DNA Şablon hazırlanması
- E. standart bir laboratuar gerilme plazmid arıtınız örneğin E. coli gibi, coli DH5a veya bir ticari kit ile Nova Mavi. Alternatif olarak, doğrusal bir PCR ürünü, benzer şekilde, ticari bir kit ile daha da arıtılabilir. Sadece H 2 O ile DNA Zehir.
- Fenol-kloroform DNA çözümü 26 ayıklayın.
- UV emme ya da diğer uygun yöntemlerle, örneğin DNA konsantrasyonu ve saflığı belirlemek. Bu, transkripsiyon, çeviri için son derece etkili bir saf DNA kullanılması önemlidir.
2. İnce Lipid Film hazırlanması
- Kuru lipid toz tartmak ve çözücü içinde çözülür. 1-palmitoil-2-oleoil-sn-glisero-3-fosfokolin (POPC) için, stok çözeltileri, 40 mg / ml 'lik bir konsantrasyonda kloroform içinde hazırlanır.
Not: Organik çözücü madde her zaman cam pipetler ve şişeler ile ele alınması gerekir. Diğer bir deyişle, plastik kesinlikle kullanılmamalıdır. Mağaza stercihen argon altında, -20 ° C 'de hava geçirmez, çözücü dayanıklı amber cam şişeler içinde tak çözümler. - 5 ml'lik bir yuvarlak tabanlı şişe içine kısım 12 mmol POPC (40 mg / ml stok çözeltisinden 220 ul).
- Ince bir lipid filmi elde etmek için bir döner buharlaştırıcı (Şekil 1) ile birlikte çözücü buharlaşır.
- Güvenli şekilde dairesel bir klip ile damıtma tüp yuvarlak tabanlı şişeye takmak ve şişenin dönüş başlar.
- Kondansatör bobin suyun dolaşımı başlatmak. Kloroform 61.2, düşük bir kaynama noktası ° C, normal atmosfer basıncında olduğu için bir ısıtma banyosu kullanarak, gerekli değildir.
- Sistem musluğunu açık olduğunu kontrol ederek atmosfere açık olduğundan emin olun. Vakum pompası açın ve yavaş yavaş sisteme vakum uygulamak için valfli yakın.
- Ince bir lipid filmi opak bir filmin oluşturulması için döner buharlaştırma sırasında yuvarlak tabanlı şişeye duvarlarında yatırılırBu gözle görülebilir. Döner buharlaşma 0.5-2 saat devam edelim. Süreci durdurmak için, yavaş yavaş, vakum basıncı serbest değiştirilmesini durdurmak ve yuvarlak tabanlı bir şişe çıkarın.
3. Lipid Tekrar Süspansiyonu ve veziküllü Homojenizasyon
- Direk tabanı yuvarlak bir şişeye ince lipid filmi için, 1 ml 2 M 18.2 H O ekleyin. Maksimum hızda şiddetle vorteks çözüm, yaklaşık 3.200 rpm, lipid filmi gözle gözlemlenebilir cam, ayrılana kadar.
- 2 mL mikrosantrifüj tüpü içine lipid dispersiyon aktarın.
- Set-up bir halka bir 5 mm uçlu bir homojenizatör tutmak için stand. Güvenli bir şekilde tutmak için yağ dispersiyonu homojenleştirici altında bir mikrosantrifüj standı yerleştirin. 18.2 MQ H 2 O ucu çeker ve birkaç saniye çalıştırarak homojenizatör dağıtma elemanı durulayın.
- Th sağlanması lipid dispersiyon içine doğrudan dispersiyon eleman yerleştirinucunda mikrosantrifüj tüp alt dokunmaz. Güç seviyesi 4 veya 14.000 rpm'de 1 dakika homojenize.
4. Vezikül Ekstrüzyon ve Liyofilizasyon
- Bir dış kılıf oluşur mini-ekstruder (Şekil 2), ve iki teflon iç zar destekler, iki O-ring, ve bir Teflon yatak evler tutucu somun parçaları birleştirin.
- Iç membran desteklerin oluklar içine iki O-ring yerleştirin. Prewet iki filtre ve bir 400 nm membran. Filtreler aralarında membranı ile O-ring içinde teflon karşı yerleştirilir.
- Membran O-halkalar arasında iki filtre ile çevrili bir membran ile ekstrüder dış kasa içine destekler yerleştirin. Muhafaza içinde teflon yatak yerleştirin ve tutucu somunu takın ve elle sıkın.
- Iki şırınga durulayın ve 18.2 M su ile bir doldurun. Küçük s içine şırınga iğneleri kaydedinekstruder düzeneğinin her iki ucundaki Teflon oles. Iğneler kolayca slayt gerekir, iğne zorlamayın. Ekstruder konut ve sıkın içine şırınga ile ekstruder sabitleyin.
- Yavaş yavaş bir şırınga dışında ve diğer içine su iterek ekstrüzyon yoluyla su geçirin. Bu, bir pasaj temsil eder. Mevcut sızıntı vardır sağlamak üç pasajların toplam tekrarlayın. Su şırınga çıkarın ve su atmayın.
- Numune ile bir şırınga ile doldurun, ekstrüdere bağlayın ve yukarıdaki adım 4.3 'de açıklandığı gibi yavaş zarından vezikül çözeltisi geçmektedir. 11 pasajların toplam 10x tekrarlayın. Ekstrüzyon işlemi ilerledikçe, örnek daha az bulanık ve zar boyunca itmek için kolay olacaktır. Direncinde ani bir düşüş, ancak genellikle bir membran kırılması gösterir.
- Ekstrüde vezikül çözüm aktarın ve mikrosantrifüj tüpleri içine veziküllerin 40 ul hacimde olun.Flaş kuru buz ya ya da sıvı azot her kısım dondurma.
- 30 gecede bir santrifüj evaporatör ° C ile her kısım Lyophilize -20 ° C'de liyofilize boş kesecikler Mağaza
5. Transkripsiyon-çeviri Makine Kapsüllenen
- Transkripsiyon yapıldı reaksiyon bileşenleri karıştırmak ve RNaz inhibitörü, 20 adet ekleyin. Buz üzerinde inkübe edin.
- DNA şablonu ekleyin. Bir kontrol reaksiyonu için, 250 bir plazmid kodlayan mVenus ng veya bir T7 promotörü arkasına transkripsiyonel benzer bir floresan protein ve güçlü E. coli, ribozom bağlanma yeri tavsiye edilir.
- RNase içermeyen su ile nihai hacim 25 ul getirin.
- Adım 5.3 'de monte edilmiş, reaksiyon 10 ul ile liyofilize veziküllerin Hidrat bir kısım (adım 4.6' dan itibaren). Kısaca vorteks karışım veziküller yeniden süspanse kadar. Bu en az 30 saniye almalıdır.
- 3 boyunca buz üzerinde inkübe edilir, reaksiyon0 dakika veziküller şişmeye izin vermek.
- 50 mM Tris-HCI, 50 mM NaCI, pH 7.4 ile 27.0 ml ve 20.2 mg / ml Proteinaz K DNaz ve RNaz 1.5 ul içine veziküllerin 1.5 ul ekleyerek 30 ul son hacim olması için 20 kat vezikül seyreltin Proteinaz K alternatif extravesicular malzeme düşmesine aynı zamanda bu noktada ilave edilebilir.
- 37, en az 2.5 saat boyunca inkübe ° C.
6. Mikroskopla inceleme
- Farklı zaman noktalarında veziküller ve floresan protein üretim ilerleme inceleyin. Veziküller 400 nm vezikül ekstrüzyonu için kullanılan membranın gözenek boyutundan daha büyük bir çapa sahip olacaktır.
- Standart bir mikroskop lamı üzerine bir 20 x 5 mm silikon ayırıcı koyarak bir örnek odası hazırlayın. Örnek odasına veziküllerin Pipet 10 ul. Odanın içinde bir silikonlu cam kapak kayma yerleştirin.
- Bir 63X yağ dağılımı o ile veziküller dikkatsömürülen floresan protein için uygun filtre seti kullanılarak parlak bir alan ve floresan mikroskobu ile r benzer amaçlı.
Representative Results
Flüoresans mikroskobu extravesicular malzeme enzimatik olarak bozulmuş olduğu için bu floresans sadece (Şekil 3), kesecikler içinde görülmektedir ortaya koymaktadır. MVenus ifadesi için, İntraveziküler floresan 37 ° C'de 1.5 saat sonra, görülmeye başlar ve 6 saat içinde maksimum floresans yoğunluğunu ulaşır. Uygun sıcaklık ve inkübasyon süresi, kullanılan özel yapıları bağlı olarak değişebilir. Örneğin, farklı floresan proteinleri bir sıcaklıkta bağımlı bir şekilde çok farklı olgunlaşır. Diğer bir deyişle, protein üretim gözlem protein sentezi ve katlama üzerine değil, aynı zamanda kromofor oluşumu sadece bağımlı değildir. Genel olarak protein sentezi transkripsiyon ve çeviri için gerekli olan 27 azaltılmış ürün akışı için izin vermek için membran proteini içeren gözenekler artırılabilir.
Bu benzer bir transkripsiyon-translasyon yürütmek için tavsiye ediliryararlanan genetik yapı işlevsel olduğundan emin olmak için vezikül yokluğunda yon reaksiyonu. Bu kontrol Reaksiyon daha kolay fluoresans spektroskopisi yerine mikroskopisi tarafından izlenir. Şekil 4, bir yapı kodlama mVenus in vitro transkripsiyon, çeviri reaksiyonunda bir göstermektedir. Kesilmemiş reaksiyonlar benzer İntraveziküler reaksiyonlar çok daha yüksek toplam floresan yoğunlukları verir. Toplam hacmi İntraveziküler extravesicular hacmi (yani seyreltme etkisi) 'den daha az olduğu için bu kapsülleme etkinliği nedeniyle ve.

Şekil 1. Döner buharlaştırıcı ve vakum pompası. Büyük resim görmek için buraya tıklayın .
Şekil 2.. Konut ve ekstruder parçaları ayrı ayrı gösterilir. Soldan sağa, şırınga, tutucu somun, teflon rulman, iç membran birbirine bakan siyah O-ring, ekstruder dış kaplama, ve ikinci şırınga ile destekler. Daha büyük resim için tıklayınız .
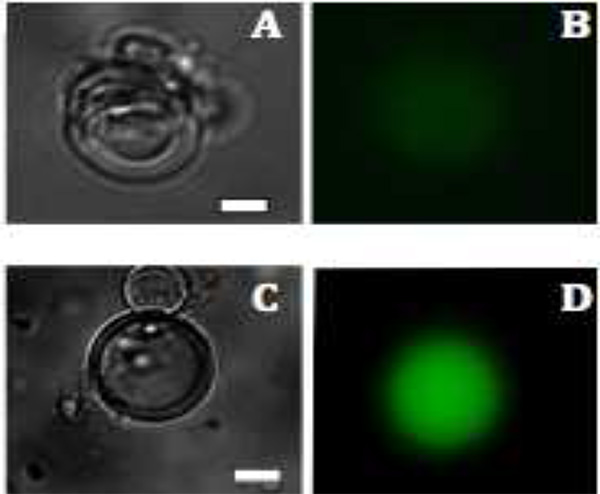
. Lipozomlarda mVenus protein üretimi Şekil 3 Floresan görüntüleri A ve C:.. Çok katmanlı veziküller Aydınlık alan görüntüleri 1.5 saat ve 2.5 saat sonra B ve &# 160; D: mVenus üretimi floresan (yeşil renkli) 1.5 ve 2.5 saat, sonra sırasıyla görüntülenmiştir. Ölçek çubuğu 20 mikron. Daha büyük resim için tıklayınız .
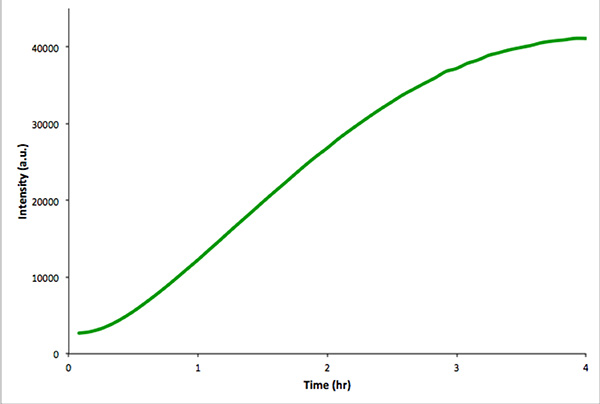
Şekil 4,. MVenus in vitro transkripsiyon ve çeviri kapsüllenmemiş in vitro kontrol reaksiyonu. Flüoresan yoğunluğu, 4 saat boyunca her 5 dakikada bir ölçüldü. Veriler bir Real-Time PCR cihazı ile elde edildi. Daha büyük resim için tıklayınız .
Discussion
Hücre içermeyen sentetik biyoloji henüz emekleme aşamasında olmasına rağmen, gelişmeler giderek daha karmaşık hücre-benzeri sistemler yapılabilir hangi bir temel atılmıştır var. Veziküller 28 tam olarak tanımlanmış bileşenler 9 içeriden transkripsiyon-çeviri makine sulandırma çevreye duyarlı yapay cells17, 18 inşa daha sonra çabaları kolaylaştırmak özellikle anlamlıydı. Benzer şekilde, yapay hücre çalışmaları evrimsel süreçleri 4,29,30 soruşturma için kullanılmıştır, mekanik RNA ayrıntıları ve protein sentezi 22,31, metabolik load32, 33 etkiler ve viral parçacıkların 34 montaj. Önemli olarak, yeterli bilgiye şimdi temel hücresel işlev, bu daha önceki raporlar ve burada tarif edilen protokollere yapılan aşağıdaki laboratuar vezikül içinde yeniden tesis edilebilir bulunmaktadır.
, Anlatılan kapsülleme pr kolay olmasının yanı sıra,ocedure birçok faydası vardır. Örneğin, çok sayıda boş, liyofilize edilmiş bir vezikül alikotları önceden yapılabilir ve daha sonra kullanılmak üzere -20 ° C'de saklanır. Protokol organik çözücüler, ciddi sıcaklık değişiklikleri, ya da diyaliz uzun süre tabi olmayan biyolojik moleküllerin yok. Biz gerektiği gibi prosedürün yumuşaklığı ek bileşenlerin dahil kolaylaştıracaktır bekliyoruz. Ayrıca kapsülleme veya transkripsiyon-çeviri verimlilik membran lipid bileşimi değişen yan etkiler gözlenen değil. Bu nedenle, membran proteinleri, belirli morfolojileri veya görselleştirme ile birleşmesiyle daha yumuşak lipidler makul yararlanılabilir.
Anlatılan yöntemin başlıca sınırlaması edilen veziküllerin boyutu veya lameller, homojen olmayan olmasıdır. Birçok uygulama için, bu zorlukların verilerin yorumlanması karışmaz. Bununla birlikte gerekirse, ek adımlar dar inci dahil edilebilirE boyut dağılımı ve kapsülleme sonra böyle ekstrüzyon daha fazla mermi olarak membran katmanları, azaltmak, donma-çözülme, ya da diyaliz 35. Bu ve diğer sorunları aşmak Kuşkusuz daha iyi yöntemler geliştirilecektir. O zamana kadar, biz protokol de hücresel taklit yapımı için uygun olması için burada açıklanan bulabilirsiniz.
Disclosures
Yazarlar ifşa hiçbir şey yok.
Acknowledgments
Yazarlar Armenise-Harvard Vakfı, Marie-Curie Trentino COFUND (ACS), Trento (Ecomm) Özerk Eyaleti ve finansman için CIBIO kabul.
Materials
| Name | Company | Catalog Number | Comments |
| Quick Spin Mini-prep kit | Qiagen | 27104 | |
| Spectrometer | NanoDrop 1000 | NDB767ND | |
| POPC | Avanti Polar Lipids | 770557 | MW 760 g/mol Transition Temp -2 °C CAS# 26853-31-6 |
| Ethanol | Sigma Aldrich | 459836 | Anhydrous, >99.5% |
| Phenol-choloroform-isoamyl alcohol 25:24:1, for molecular biology use | Sigma Aldrich | P3803-100mL | Saturated with 10 mM Tris, pH 8.0, 1 mM EDTA |
| Chloroform | Biotech Grade Fluka | 496189-1L | Contain ethanol at 0.5-1.0% v/v as stabilizer |
| Brown amber glass bottles | VWR | 89043-518 | 55X 48 mm |
| Rotary evaporator | Buchi Rotovapor R-210/Sigma | Z563846EU-1EA | With jack and water bath, 29/32 joint 240 V |
| Analog vortex mixer | VWR | 945300 | Speed 1,000-3,200 rpm |
| Homogenizer | IKA T10 Basic Ultra-Turbax | 3420000 | |
| Mini-extruder | Avanti Polar Lipids | 610020 | |
| Extruder filters | Whatman | 610014 | drain disc 10 mm |
| Extruder polycarbonate membrane 400 nm | Whatman | 61007 | nuclepore polycarbonate |
| Speed vacuum | Labconco | 7970011 | Centritrap DNA concentrator |
| PURExpress kit | New England Biolabs | NRM #E6800S | |
| RNAse inhibitor (40,000 U/ml) | New England Biolabs | #M0307S | |
| Proteinase K (20.2 mg/ml) | Fermentas | #EO0491 | |
| Microscope | Zeiss Observer Z1with a AxioCam MRm camera | ||
| RealTime | CFX96 Real time PCR Detection System (Biorad) | ||
| Silicon press to seal -Molecular Probe | Life Technologies | P18174 | Resistant from -25-30 °C |
| Siliconized glass circle cover slides | Hampton Research | HR3-231 | Diameter= 22 mm |
| ImageJ | NIH |
References
- Forster, A. C., Church, G. M. Towards synthesis of a minimal cell. Mol. Syst. Biol. 2, 1-10 (2006).
- Noireaux, V., Maeda, Y. T., Libchaber, A. Development of an artificial cell, from self-organization to computation and self-reproduction. Proc. Natl. Acad. Sci. U.S.A. 108, 3473-3480 (2011).
- Harris, D. C., Jewett, M. C. Cell-free biology: Exploiting the interface between synthetic biology and synthetic chemistry. Curr. Opin. Biotech. 23, (2012).
- Nishikawa, T., Sunami, T., Matsuura, T., Yomo, T. Directed Evolution of Proteins through In Vitro Protein Synthesis in Liposomes. J. Nucleic Acids. 2012, 1-11 (2012).
- Forlin, M., Lentini, R., Mansy, S. S.
- Chiarabelli, C., Stano, P., Anella, F., Carrara, P., Luisi, P. L. Approaches to chemical synthetic biology. FEBS Lett. 586, 2138-2145 (2012).
- Noireaux, V., Shin, J. Efficient cell-free expression with the endogenous E. Coli RNA polymerase and sigma factor 70. J. Biol. Eng. 4, 1-9 (2010).
- Fujiwara, K., Nomura, S. -iM. Condensation of an Additive-Free Cell Extract to Mimic the Conditions of Live Cells. PLoS ONE. 8, e54155 (2013).
- Shimizu, Y., et al. Cell-free translation reconstituted with purified components. Nat. Biotechnol. 19, 751-755 (2001).
- Hosoda, K., et al. Quantitative Study of the Structure of Multilamellar Giant Liposomes As a Container of Protein Synthesis Reaction. Langmuir. 24, 13540-13548 (2008).
- Sunami, T., Matsuura, T., Suzuki, H., Yomo, T.
- Murtas, G., Kuruma, Y., Bianchini, P., Diaspro, A., Luisi, P. L. Protein synthesis in liposomes with a minimal set of enzymes. Biochem. Biophys. Res. Commun. 363, 12-17 (2007).
- Pereira de Souza, T., Stano, P., Luisi, P. L. The Minimal Size of Liposome-Based Model Cells Brings about a Remarkably Enhanced Entrapment and Protein Synthesis. ChemBioChem. 10, 1056-1063 (2009).
- Caschera, F., et al. Programmed Vesicle Fusion Triggers Gene Expression. Langmuir. 27, 13082-13090 (2011).
- Noireaux, V., Bar-Ziv, R., Godefroy, J., Salman, H., Libchaber, A. Toward an artificial cell based on gene expression in vesicles. Phys. Biol. 2, P1-P8 (2005).
- Shin, J., Noireaux, V. An E. coli Cell-Free Expression Toolbox: Application to Synthetic Gene Circuits and Artificial Cells. ACS Synth. Biol. 1, 29-41 (2012).
- Kobori, S., Ichihashi, N., Kazuta, Y., Yomo, T. A controllable gene expression system in liposomes that includes a postive feedback loop. Mol. Syst. Biol. 9, 1282-1285 (2013).
- Martini, L., Mansy, S. S. Cell-like Systems with Riboswitch Controlled Gene Expression. Chem. Commun. 47, 10734-10736 (2011).
- Noireaux, V., Bar-Ziv, R., Libchaber, A. Principles of cell-free genetic circuit assembly. Proc. Natl. Acad. Sci. U.S.A. 100, 12672-12677 (2003).
- Sunami, T., et al. Detection of Association and Fusion of Giant Vesicles Using a Fluorescence-Activated Cell Sorter. Langmuir. 26, 15098-15103 (2010).
- Saito, H., et al. Time-Resolved Tracking of a Minimum Gene Expression System Reconstituted in Giant Liposomes. ChemBioChem. 10, 1640-1643 (2009).
- Lentini, R., et al. Fluorescent Proteins and in Vitro Genetic Organization for Cell-Free Synthetic Biology. ACS Synth. Biol. , (2013).
- Du, L., Villarreal, S., Forster, A. C. Multigene Expression In Vivo: Supremacy of Large Versus Small Terminators for T7 RNA Polymerase. Biotechnol. Bioeng. 109, 1043-1050 (2011).
- Trochilin, V. P., Weissig, V. Liposomes: A Practical Approach. , 2nd edn, Oxford University Press. (2003).
- Walde, P., Cosentino, K., Engel, H., Stano, P. Giant vesicles: preparations and applications. ChemBioChem. 11, 848-865 (2010).
- Sambrook, J., Russell, D. W. Molecular Cloning. , 3rd edn, Cold Spring Harbor Laboratory Press. (2001).
- Noireaux, V., Libchaber, A. A vesicle bioreactor as a step toward an artificial cell assembly. Proc. Natl. Acad. Sci. U.S.A. 101, 17669-17674 (2004).
- Yu, W., et al.
- Caschera, F., et al. Stable vesicles composed of monocarboxylic or dicarboxylic fatty acids and trimethylammonium amphiphiles. Langmuir. 27, 14078-14090 (2011).
- Pereira de Souza, T., Steiniger, F., Stano, P., Fahr, A., Luisi, P. L. Spontaneous crowding of ribosomes and proteins inside vesicles: a possible mechanism for the origin of cell metabolism. ChemBioChem. 12, 2325-2330 (2011).
- Niederholtmeyer, H., Xu, L., Maerkl, S. J. Real-Time mRNA Measurement during an in Vitro Transcription and Translation Reaction Using Binary Probes. ACS Synth. Biol. 10, (2012).
- Stögbauer, T., Windhager, L., Zimmer, R., Rädler, J. Experiment and mathematical modeling of gene expression dynamics in a cell-free system. Integr. Biol. 4, 494-501 (2012).
- Lazzerini-Ospri, L., Stano, P., Luisi, P., Marangoni, R. Characterization of the emergent properties of a synthetic quasi-cellular system. BMC Bioinformatics. 13, 1-10 (2011).
- Shin, J., Jardine, P., Noireaux, V. Genome Replication, Synthesis, and Assembly of the Bacteriophage T7 in a Single Cell-Free Reaction. ACS Synth. Biol. 1, 408-413 (2012).
- Zhu, T. F., Szostak, J. W.
