

Summary
本明細書で我々は、小胞の調製、転写および翻訳機構のカプセル化、およびタンパク質の産生を監視するための簡単な方法を記載する。得られた無細胞系がますます複雑な細胞模倣物の構築の開始点として使用することができる。
Abstract
個々の分子から分子のシステムへの関心がシフトしたように、研究室が増えて、より良い携帯人生の複雑さを表す携帯ミミックボトムアップからビルドしようとしてきた。これまでに、油中水型エマルジョン、マイクロ流体デバイス、および小胞の利用を含む区画セルラ模倣物の構築を取ることができるパスの数が存在する。利用可能なオプションには、それぞれ固有の利点と欠点を有する。例えば、油中水型エマルジョンは、高い封入効率を与えるが、よく生きた細胞の透過障壁を模倣しない。本明細書に記載の方法の主な利点は、すべての実装が容易で安価であることである。転写·翻訳機構は、遠心エバポレーター、押出機などの一般的なインストゥルメンテーションを、悪用プロセスを通じてリン脂質小胞の内部に封入されている。反応を蛍光分光法によって監視されます。プロトコルSは、組換えタンパク質の発現、細胞の模倣の建設、携帯生命の最小要件の探査、または遺伝回路の組み立てに適合させることができる。
Introduction
無細胞は、in vitroで転写·翻訳反応と合成脂質からの小胞の生成は、新しいものではありません。しかし、模倣携帯に2を組み合わせること。E. challenging1-6かなり多くあるT7 RNAポリメラーゼの有無にかかわらず大腸菌細胞抽出物を転写-翻訳機構7,8の供給源として使用することができる。細胞抽出物は、タンパク質発現および折り畳みを容易にすることができる付加的な細胞成分の存在から恩恵を受ける。あるいは、個別にRNAと蛋白質分子の精製混合物が、9 PUREシステム、すなわち 、膀胱内タンパク質合成4,10-14を媒介するために使用することができる。 PUREシステムは完全に定義された細胞の模倣の建設を可能にし、細胞抽出物で見つかったヌクレアーゼ活性に悩まされることはありません。実際には、このことにより、低封入効率11とプロセスを容易にする、はるかに少ないDNAテンプレートが必要であることを意味する16-18報告されている。
転写翻訳反応を監視する最も簡単な方法は、遺伝的にコード要素の蛍光又はルミネセンスを測定することである。 インビトロ反応で頻繁に放射性標識によって測定されるが、典型的には、ホタルルシフェラーゼ19又はGFPが用いられる。蛍光検出は、さらにそれによって生物学のような過程の確率的性質にいくつかの洞察を提供し、サイトメトリーベースのメソッドを介して小胞20,21の個体群のモニタリングを可能にします。これらの監視方法は、iのと互換性のある蛍光タンパク質の集合を含むデザインルールとからビルドするのに部品のライブラリの小さなセットを定義するために使用されているN体外転写翻訳22、表現22、シグマ因子16の活動、および転写ターミネーター23の効率に遺伝子組織の影響。それにもかかわらず、 インビトロで予測可能な遺伝的にエンコードされた装置を構築する能力を高めるために行われる必要があり、その多くが残っている。
小胞を作るために利用できる多くの方法がある。最も一般的な方法は、水性solution24で再懸濁し、続いてガラス表面上に薄い脂質膜の生成に依存する。水溶液転写 - 翻訳機構が含まれている場合、例えば、次いで形成された小胞の割合は、タンパク質産生のために必要なコンポーネントを含むであろう。しかしながら、このような方法のカプセル化効率は、小胞のごく一部がアクティブであることを意味し、低い。はるかに高い封入EFFが特徴の代替メソッドの多くiciencyは、小胞の油中水型エマルジョン液滴の変換を利用する。そのような方法は、将来的には一般的であろうと思われているが、現在のところ、これらの方法は、特殊な装置を必要と苦しむ、変更された膜組成物25と小胞を得た。小胞のメソッドへの油中水型の明確な利点は、膜ラメラを制御するための電位である。本明細書に記載された方法は、追加の均質化工程を含むわずかな変更を加えて四方実験室11に記載の薄膜脂質膜プロトコルに基づいている。この方法は安価で、簡単で、うまく転写 - 翻訳機械のカプセル化に適した堅牢な小胞を与える。
Protocol
1。 DNAテンプレートの準備
- E.標準の実験室株からのプラスミド浄化例えば大腸菌などの大腸菌 、市販のキットで大腸菌 DH5αをまたはノヴァブルー。あるいは、線形PCR産物を、同様に市販のキットを用いて精製することができる。唯一のH 2 OでDNAを溶出。
- フェノール-クロロホルムはDNA溶液26を抽出します。
- UV吸光度または他の適当な方法によって、 例えば DNA濃度と純度を決定します。転写翻訳効率的な高純度のDNAを用いることが重要である。
2。薄い脂質フィルムを準備する
- 乾燥脂質粉末を秤量し、溶媒中に溶解する。 1 -パルミトイル-2 -オレオイル-sn-グリセロ-3 -ホスホコリン(POPC)については、ストック溶液は、40 mg / mlの濃度のクロロホルム中で調製される。
注:有機溶剤は常にガラスピペットとボトルで処理しなければなりません。言い換えれば、プラスチックを使用してはいけません。店舗s好ましくは、アルゴン下、-20℃で気密、耐溶剤アンバーガラスボトルタックソリューション。 - アリコート12マイクロモルPOPC(40 mg / mlのストック溶液220μL)5 mlの丸底フラスコに。
- 薄い脂質フィルムを生成するためのロータリーエバポレーター( 図1)で溶媒を蒸発させる。
- しっかりと円形のクリップで蒸留管に丸底フラスコを取り付け、フラスコの回転を開始。
- 凝縮器コイルを通じて水の循環を開始します。クロロホルム61.2の低沸点℃で常圧でCを持っているので、熱浴を使用して、必要ではない。
- システムはコックが開いていることを確認することにより、大気に開放されていることを確認してください。真空ポンプの電源を入れ、ゆっくりとシステムに真空を適用するためのコックを閉じてください。
- 薄い脂質膜は遮光膜を形成し、回転蒸発中の丸底フラスコの壁面上に堆積されるそれは、目で見えるようになります。ロータリーエバポレーターは0.5-2時間続けてみましょう。プロセスを停止するには、徐々に、真空圧力を解放回転を停止し、丸底フラスコを除去する。
3。脂質再懸濁およびベシクル均質
- 直接丸底フラスコ内の薄い脂質膜に1ミリリットル18.2MΩH 2 Oを加える。最大速度で激しく渦ソリューション、約3,200 rpmで、脂質膜が目で観察されたガラスから切り離しまで。
- 2 mlのマイクロチューブに脂質分散を転送します。
- セットアップリングは、5mmチップでホモジナイザーを保持するために立っている。しっかりと脂質分散を保持するためにホモジナイザー下マイクロスタンドを置きます。 18.2MΩH 2 Oで先端を浸漬し、数秒間実行することにより、ホモジナイザーの分散要素をすすぐ。
- 番目を確保脂質分散に直接分散要素を配置先端にマイクロ遠心チューブの底に触れない。電力レベル4または14,000 rpmで1分間ホモジナイズする。
4。小胞の放出と凍結乾燥
- アウターケーシングや家2テフロン内部の膜がサポートするリテーナナット、2つのOリング、および1テフロン軸受けで構成されたミニ押出機( 図2)の部品を組み立てる。
- 内部膜支持体の溝に2個のO-リングを置きます。予め湿らせつのフィルタと1 400 nmの膜。フィルタは、それらの間の膜とO-リングの内側にテフロン(登録商標)に対して配置される。
- 膜がOリングの間に2つのフィルタに囲ま膜と押出アウターケーシングにサポートしています置きます。ケーシング内にテフロンベアリングを配置し、リテーナナットを取り付け、手で締めます。
- 2シリンジをすすぎ、18.2MΩ水を持つものを記入してください。小さな時間に注射針を挿入します押出機アセンブリの両端にテフロンでoles。針は簡単にスライドするはず;針を無理に押し込まないでください。押出機ハウジングと締めに注射器で押出機を固定します。
- ゆっくりと1注射器のうち、その他の中に水を押すことにより押出機を通して水を渡します。これは、1つの通路を表している。現在の漏れがないことを確実に3通路の合計に対して、この手順を繰り返します。水の注射器を取り出し、水を処分する。
- サンプルと1シリンジを埋める、押出機に取り付け、ステップ4.3で上記のように徐々に膜を通って小胞ソリューションを渡す。 11通路の合計10倍を繰り返します。押出プロセスが進むにつれて、試料は、膜を横切ってプッシュするために以下の混濁かつ容易になるであろう。抵抗の急激な減少が、しかしながら、通常、膜の破裂を示している。
- 押し出された小胞のソリューションを転送し、マイクロチューブに小胞の40μlのアリコートを行います。Flashには、どちらのドライアイスや液体窒素の各アリコートを凍結。
- 30で一晩遠心エバポレーター℃まで、各アリコートを凍結乾燥-20℃で凍結乾燥した空胞を保存
5。転写 - 翻訳機械をカプセル化
- 転写·翻訳反応の成分を混合し、RNase阻害の20ユニットを追加。氷の上でインキュベートする。
- DNAテンプレートを追加します。コントロール反応のために、250をコードするプラスミドのmVenusのNGまたはT7転写プロモーターの後ろに類似の蛍光タンパク質と強いE.大腸菌リボソーム結合部位をお勧めします。
- RNaseフリー水で最終体積を25μlをもたらす。
- ステップ5.3で組み立て反応液10μlと凍結乾燥胞の水和物アリコート(ステップ4.6から)。簡潔にボルテックス混合ベシクルを再懸濁されるまで。これは、以下の30秒を取るべきである。
- 3のために氷の上での反応をインキュベート胞を膨潤できるように0分。
- 50mMのトリス-HCl、50mMのNaCl、pH7.4のの27.0μL及び20.2 mg / mlのプロテイナーゼKのDNaseとRNaseを1.5μlのに小胞の1.5μLを加えることにより、30μlの最終容量に20倍胞混合物を希釈また胞外物質を分解するプロテイナーゼKの代替として、この時点で添加することができる。
- 37少なくとも2.5時間インキュベート℃、
6。顕微鏡検査
- 小胞と異なる時点で蛍光タンパク質生産の進捗状況を調べる。小胞は400nmの小胞の放出のために使用される膜の孔径よりも大きな直径を有する。
- 標準的な顕微鏡スライド上に20×5ミリメートルシリコンスペーサを配置することによって、試料室を準備します。試料室への小胞のピペットを10μl。室以上シリコン処理ガラスカバースリップを置きます。
- 63X油分散Oで小胞を観察する搾取蛍光タンパク質の適切なフィルターセットを用いて明視野と蛍光顕微鏡法によるRと同様の目的。
Representative Results
蛍光顕微鏡は、胞外物質が酵素的に分解されるため、その蛍光のみ( 図3)、小胞の内側に観測される明らかにする。 mVenusの発現のために、胞内蛍光は37℃で1.5時間後に観察され始め、6時間以内に最大蛍光強度に到達する。最適な温度およびインキュベーション時間は、使用される特定の構築物に応じて変化し得る。例えば、異なる蛍光タンパク質は、温度依存的にまったく異なる成熟。換言すれば、タンパク質生産の観察は、タンパク質合成および折り畳み時だけでなく、発色団の形成にもっぱら依存しない。全体的なタンパク質合成は、転写および翻訳のために必要な27枯渇成分の流入を可能にするために、膜タンパク質の細孔を組み込むことによって高めることができる。
これは、類似した転写並進を行うことをお勧めします搾取遺伝子構築物が機能していることを確認するために小胞が存在しない場合に化反応。このコントロール反応をより簡単に蛍光分光法ではなく、顕微鏡によって監視される。 図4は、構築物、符号化mVenusのインビトロ転写翻訳反応で示している。カプセル化されていない反応が似胞内反応よりもはるかに高い総蛍光強度を与える。これは、カプセル化効率によるものであり、総胞内量は胞外体積( すなわち、希釈効果)よりもはるかに少ないからである。

図1。ロータリーエバポレーターと真空ポンプは 大きな画像を表示するには、ここをクリックしてください 。
図2ハウジングおよび押出機の部品を別々に示されている 。左から右に、注射器、リテーナナット、テフロンベアリング、内部の膜が互いに直面黒Oリング、押出アウターケーシング、及び第2のシリンジでサポートされています大きな画像を表示するには、ここをクリックしてください 。
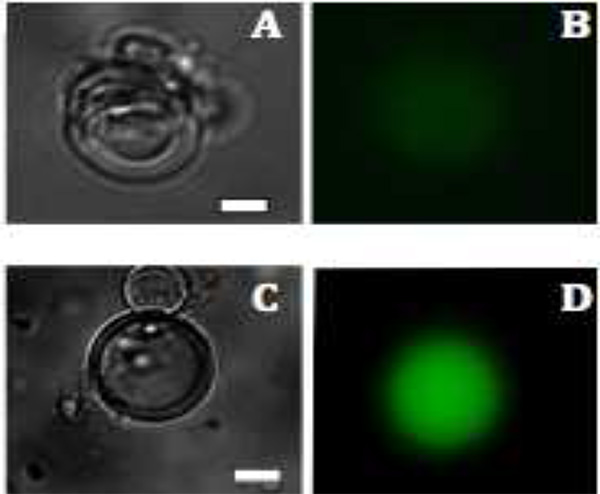
図3のリポソーム中mVenusタンパク質生産の蛍光画像とC:。。。多層小胞の明るいフィールド画像1.5時間と2.5時間後にBと  D:mVenusの生産は、それぞれ1.5と2.5時間後(緑色)蛍光によって可視化される。スケールバーは20μmである。 大きな画像を表示するには、ここをクリックしてください 。
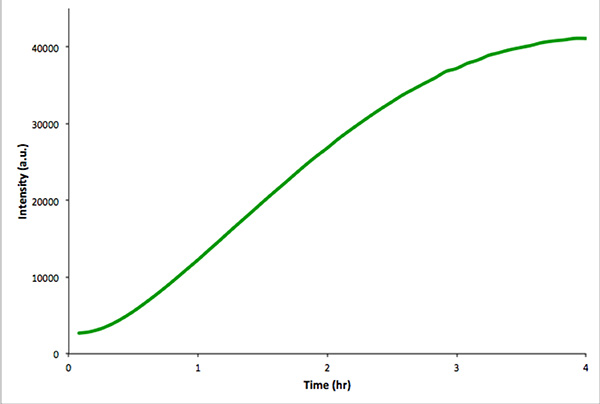
(4)in vitro転写とmVenusの翻訳でカプセル化されていないのin vitroでのコントロール反応の図 。蛍光強度を4時間かけて5分毎に測定した。データはリアルタイムPCR装置で取得した。 画像を拡大表示するには、ここをクリック 。
Discussion
無細胞合成生物学は、まだ始まったばかりであるが、進歩は、ますます複雑化する細胞のようなシステムを作ることができ、そこから基礎を築いた。完全に定義されたコンポーネントからの転写·翻訳機構の再構成胞28の9内部は環境対応人工cells17、18を構築する後の努力を促進する上で特に顕著であった。同様に、人工細胞研究は、進化の過程4,29,30、機械的なRNAの詳細およびタンパク質合成22,31、代謝load32、33の影響、およびウイルス粒子の集合体34をプローブするために使用されている。重要なのは、十分な知識は今、基本的な細胞機能が存在するこれらの以前の報告とここに記載されているプロトコルは、次の研究室での小胞の内側に再構成することができる。
、説明されたカプセルは、PRに簡単であることに加えてocedureは、いくつかの利点を持っています。例えば、多くの空、凍結乾燥された小胞のアリコートを事前に行うことができ、後の使用のために-20℃で保存した。プロトコルは、有機溶剤、急激な温度変化、または透析の長い期間に受けない生体分子を行います。我々は、必要に応じて手順の優しさは、追加コンポーネントの取り込みを促進することを期待しています。我々はまた、カプセル化または転写 - 翻訳効率に膜の脂質組成の変化に悪影響を観察していない。従って、膜タンパク質、特定の形態、または可視化の取り込みに、より適した脂質は多分悪用される可能性があります。
記載されている方法の主な制限は、結果として小胞のサイズまたはラメラで均質でないことである。多くのアプリケーションでは、これらの問題は、データの解釈に干渉しない。ただし、必要に応じて、追加のステップが狭い番目に組み込むことができ電子サイズ分布及びカプセル化の後、押出のさらなるラウンドのような膜の層を、減少させるが、凍結融解、又は透析35。間違いなく、これらおよびその他の問題を回避するより良い方法が開発される。それまで、私たちは、プロトコルがよく携帯模倣の建設のために適していることが、ここで説明を見つける。
Disclosures
著者らは、開示することは何もありません。
Acknowledgments
著者はArmenise-ハーバード財団、マリー·キュリートレンCOFUND(ACS)、トレント(ECOMM)自治州、および資金調達のためにCIBIOを認める。
Materials
| Name | Company | Catalog Number | Comments |
| Quick Spin Mini-prep kit | Qiagen | 27104 | |
| Spectrometer | NanoDrop 1000 | NDB767ND | |
| POPC | Avanti Polar Lipids | 770557 | MW 760 g/mol Transition Temp -2 °C CAS# 26853-31-6 |
| Ethanol | Sigma Aldrich | 459836 | Anhydrous, >99.5% |
| Phenol-choloroform-isoamyl alcohol 25:24:1, for molecular biology use | Sigma Aldrich | P3803-100mL | Saturated with 10 mM Tris, pH 8.0, 1 mM EDTA |
| Chloroform | Biotech Grade Fluka | 496189-1L | Contain ethanol at 0.5-1.0% v/v as stabilizer |
| Brown amber glass bottles | VWR | 89043-518 | 55X 48 mm |
| Rotary evaporator | Buchi Rotovapor R-210/Sigma | Z563846EU-1EA | With jack and water bath, 29/32 joint 240 V |
| Analog vortex mixer | VWR | 945300 | Speed 1,000-3,200 rpm |
| Homogenizer | IKA T10 Basic Ultra-Turbax | 3420000 | |
| Mini-extruder | Avanti Polar Lipids | 610020 | |
| Extruder filters | Whatman | 610014 | drain disc 10 mm |
| Extruder polycarbonate membrane 400 nm | Whatman | 61007 | nuclepore polycarbonate |
| Speed vacuum | Labconco | 7970011 | Centritrap DNA concentrator |
| PURExpress kit | New England Biolabs | NRM #E6800S | |
| RNAse inhibitor (40,000 U/ml) | New England Biolabs | #M0307S | |
| Proteinase K (20.2 mg/ml) | Fermentas | #EO0491 | |
| Microscope | Zeiss Observer Z1with a AxioCam MRm camera | ||
| RealTime | CFX96 Real time PCR Detection System (Biorad) | ||
| Silicon press to seal -Molecular Probe | Life Technologies | P18174 | Resistant from -25-30 °C |
| Siliconized glass circle cover slides | Hampton Research | HR3-231 | Diameter= 22 mm |
| ImageJ | NIH |
References
- Forster, A. C., Church, G. M. Towards synthesis of a minimal cell. Mol. Syst. Biol. 2, 1-10 (2006).
- Noireaux, V., Maeda, Y. T., Libchaber, A. Development of an artificial cell, from self-organization to computation and self-reproduction. Proc. Natl. Acad. Sci. U.S.A. 108, 3473-3480 (2011).
- Harris, D. C., Jewett, M. C. Cell-free biology: Exploiting the interface between synthetic biology and synthetic chemistry. Curr. Opin. Biotech. 23, (2012).
- Nishikawa, T., Sunami, T., Matsuura, T., Yomo, T. Directed Evolution of Proteins through In Vitro Protein Synthesis in Liposomes. J. Nucleic Acids. 2012, 1-11 (2012).
- Forlin, M., Lentini, R., Mansy, S. S.
- Chiarabelli, C., Stano, P., Anella, F., Carrara, P., Luisi, P. L. Approaches to chemical synthetic biology. FEBS Lett. 586, 2138-2145 (2012).
- Noireaux, V., Shin, J. Efficient cell-free expression with the endogenous E. Coli RNA polymerase and sigma factor 70. J. Biol. Eng. 4, 1-9 (2010).
- Fujiwara, K., Nomura, S. -iM. Condensation of an Additive-Free Cell Extract to Mimic the Conditions of Live Cells. PLoS ONE. 8, e54155 (2013).
- Shimizu, Y., et al. Cell-free translation reconstituted with purified components. Nat. Biotechnol. 19, 751-755 (2001).
- Hosoda, K., et al. Quantitative Study of the Structure of Multilamellar Giant Liposomes As a Container of Protein Synthesis Reaction. Langmuir. 24, 13540-13548 (2008).
- Sunami, T., Matsuura, T., Suzuki, H., Yomo, T.
- Murtas, G., Kuruma, Y., Bianchini, P., Diaspro, A., Luisi, P. L. Protein synthesis in liposomes with a minimal set of enzymes. Biochem. Biophys. Res. Commun. 363, 12-17 (2007).
- Pereira de Souza, T., Stano, P., Luisi, P. L. The Minimal Size of Liposome-Based Model Cells Brings about a Remarkably Enhanced Entrapment and Protein Synthesis. ChemBioChem. 10, 1056-1063 (2009).
- Caschera, F., et al. Programmed Vesicle Fusion Triggers Gene Expression. Langmuir. 27, 13082-13090 (2011).
- Noireaux, V., Bar-Ziv, R., Godefroy, J., Salman, H., Libchaber, A. Toward an artificial cell based on gene expression in vesicles. Phys. Biol. 2, P1-P8 (2005).
- Shin, J., Noireaux, V. An E. coli Cell-Free Expression Toolbox: Application to Synthetic Gene Circuits and Artificial Cells. ACS Synth. Biol. 1, 29-41 (2012).
- Kobori, S., Ichihashi, N., Kazuta, Y., Yomo, T. A controllable gene expression system in liposomes that includes a postive feedback loop. Mol. Syst. Biol. 9, 1282-1285 (2013).
- Martini, L., Mansy, S. S. Cell-like Systems with Riboswitch Controlled Gene Expression. Chem. Commun. 47, 10734-10736 (2011).
- Noireaux, V., Bar-Ziv, R., Libchaber, A. Principles of cell-free genetic circuit assembly. Proc. Natl. Acad. Sci. U.S.A. 100, 12672-12677 (2003).
- Sunami, T., et al. Detection of Association and Fusion of Giant Vesicles Using a Fluorescence-Activated Cell Sorter. Langmuir. 26, 15098-15103 (2010).
- Saito, H., et al. Time-Resolved Tracking of a Minimum Gene Expression System Reconstituted in Giant Liposomes. ChemBioChem. 10, 1640-1643 (2009).
- Lentini, R., et al. Fluorescent Proteins and in Vitro Genetic Organization for Cell-Free Synthetic Biology. ACS Synth. Biol. , (2013).
- Du, L., Villarreal, S., Forster, A. C. Multigene Expression In Vivo: Supremacy of Large Versus Small Terminators for T7 RNA Polymerase. Biotechnol. Bioeng. 109, 1043-1050 (2011).
- Trochilin, V. P., Weissig, V. Liposomes: A Practical Approach. , 2nd edn, Oxford University Press. (2003).
- Walde, P., Cosentino, K., Engel, H., Stano, P. Giant vesicles: preparations and applications. ChemBioChem. 11, 848-865 (2010).
- Sambrook, J., Russell, D. W. Molecular Cloning. , 3rd edn, Cold Spring Harbor Laboratory Press. (2001).
- Noireaux, V., Libchaber, A. A vesicle bioreactor as a step toward an artificial cell assembly. Proc. Natl. Acad. Sci. U.S.A. 101, 17669-17674 (2004).
- Yu, W., et al.
- Caschera, F., et al. Stable vesicles composed of monocarboxylic or dicarboxylic fatty acids and trimethylammonium amphiphiles. Langmuir. 27, 14078-14090 (2011).
- Pereira de Souza, T., Steiniger, F., Stano, P., Fahr, A., Luisi, P. L. Spontaneous crowding of ribosomes and proteins inside vesicles: a possible mechanism for the origin of cell metabolism. ChemBioChem. 12, 2325-2330 (2011).
- Niederholtmeyer, H., Xu, L., Maerkl, S. J. Real-Time mRNA Measurement during an in Vitro Transcription and Translation Reaction Using Binary Probes. ACS Synth. Biol. 10, (2012).
- Stögbauer, T., Windhager, L., Zimmer, R., Rädler, J. Experiment and mathematical modeling of gene expression dynamics in a cell-free system. Integr. Biol. 4, 494-501 (2012).
- Lazzerini-Ospri, L., Stano, P., Luisi, P., Marangoni, R. Characterization of the emergent properties of a synthetic quasi-cellular system. BMC Bioinformatics. 13, 1-10 (2011).
- Shin, J., Jardine, P., Noireaux, V. Genome Replication, Synthesis, and Assembly of the Bacteriophage T7 in a Single Cell-Free Reaction. ACS Synth. Biol. 1, 408-413 (2012).
- Zhu, T. F., Szostak, J. W.
