The strategies to generate murine mTERT-ER and LSL-mTERT knock-in alleles were described in Figure 1A and 1B. Specifically, a LoxP–triple Stopper–Neo–LoxP fragment was inserted between exon 1 and exon 2 of mTERT locus to generate LSL-mTERT allele. To generate mTERT-ER allele, ERT2-LBD domain in frame with the mTERT gene was inserted into the N-terminus of exon 1. We showed that when telomerase was transiently reactivated in telomere dysfunction mice by treating them with 4-OHT pellets for 4 weeks, the degenerative phenotype could be ameliorated in multiple organs such as brain (Figure 2A) and testes (Figure 2B). In order to determine the impact of telomerase reactivation on cells cultured in vitro, we isolated neural stem cells (NSCs) from late generation G4 LSL-mTERT and G4 mTERT-ER mice. We showed that when telomerase was reactivated in telomere dysfunctional NSCs, the self-renewal capability of NSCs was greatly increased (Figure 3A), and in vitro neurogenesis was also significantly enhanced (Figure 3B). In order to determine the impact of telomerase reactivation on tumorigenesis in the context of telomere dysfunction, we generated PB-Cre4 PtenL/L p53L/L LSL-mTERTL/L (prostate tumor model) and Atm-/- mTERTER/ER (thymic T-cell lymphoma model) late generation cohorts. When telomerase was reactivated by tamoxifen treatment in these mice with injection (Figure 4A) or 4-OHT pellets for 8 weeks (Figure 4B), the tumorigenesis was greatly enhanced in both prostate tumor model (Figure 4A) and thymic T-cell lymphoma model (Figure 4B). Lastly, we reported the protocols of performing telomere FISH on FFPE mouse brain tissues (Figure 5) and metaphase chromosomes (Figure 5, inset).

Figure 1: (A) mTERT-ER knock-in strategy. TV, targeting vector; WT, wild type allele; KI, knock-in allele; LA, left arm; RA, right arm; E1, exon 1; E2, exon 2; neo, pgk promoter-driven neomycin resistant gene; ER, modified estrogen receptor (ERT2) ligand binding domain; DT, dyphteria toxin gene. (B) LSL-mTERT knock-in strategy. TV, targeting vector; WT, wild type allele; KI, knock-in allele; LA, left arm; RA, right arm; E1, exon 1; E2, exon 2; neo, pgk promoter-driven neomycin resistant gene; STOPPER, three repetitive transcriptional stop sequences; DT, dyphteria toxin gene.

Figure 2: Representative data to show telomerase reactivation ameliorates degenerative phenotypes of organs. Representative images of brains (left) and testes (right) of G0 and G4 mTERT-ER mice treated with placebo or 4-OHT for 4 weeks. Scale bars indicate 50 µm. Re-print with permission from 20. Please click here to view a larger version of this figure.
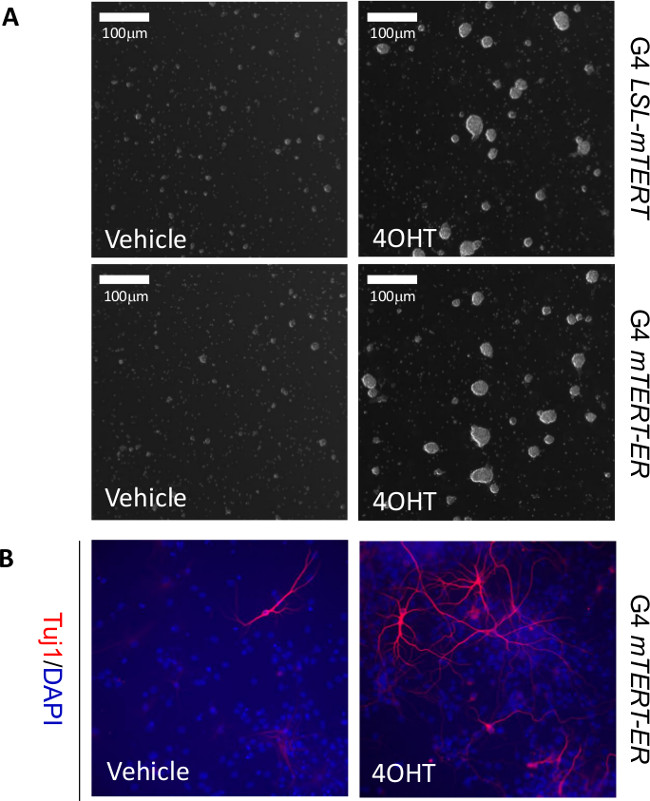
Figure 3: Representative data to show telomerase reactivation enhances self-renewal and neurogenesis of neural stem cells in vitro. (A) Representative images of neural stem cells isolated from G4 LSL-mTERT (upper panel) and G4 mTERT-ER (lower panel) growing in stem cell medium treated with vehicle or 4-OHT. (B) Representative images of in vitro neurogenesis (Tuj1+) of neural stem cells isolated from G4 mTERT-ER cultured in differentiation medium (with 1% FBS) treated with vehicle or 4-OHT. Scale bars indicate 100 µm.
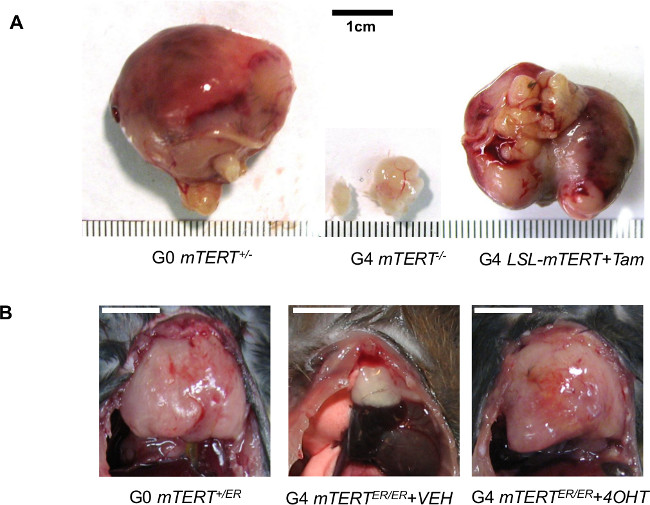
Figure 4: Representative data to show telomerase reactivation enhances tumorigenesis against the backdrop of genomic instability. (A) Representative images of prostate tumors dissected from G0 PB-Cre4 PtenL/L p53L/L LSL-mTERT+/-, G4 PB-Cre4 PtenL/L p53L/L LSL-mTERT-/-, and G4 PB-Cre4 PtenL/L p53L/L LSL-mTERTL/L treated with tamoxifen. Re-print with permission from 22. (B) Representative images of thymic T-cell lymphomas dissected from G0 Atm-/- mTERT+/ER, G4 Atm-/- mTERTER/ER treated with vehicle, and G4 Atm-/- mTERTER/ER treated with 4-OHT for 8 weeks. Scale bars indicate 1 cm.
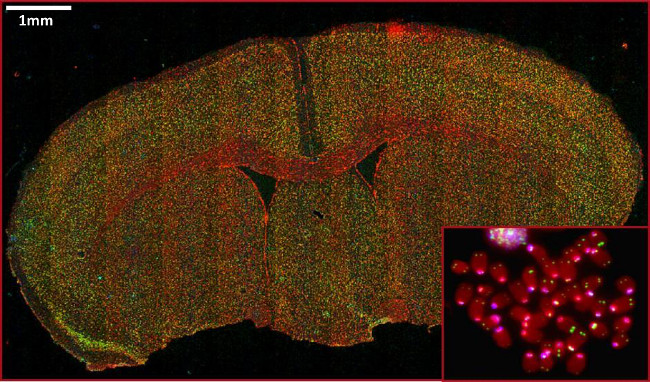
Figure 5: Representative images of telomeric and centromeric FISH on FFPE mouse brain tissue and metaphase (inset). Red indicates DNA, green indicates telomere staining, and cyan indicates centromere staining. Scale bars indicate 1 mm.
