1. Stomatal Conductance
- Measurements of stomatal conductance
NOTE: The authors made use of a simple type of steady state porometer (Decagon SC1). The porometer’s design has the advantage of small size, intuitive manual operation and high reliability. When measuring stomatal conductance in the field, make sure to minimize the distance between measured individuals for a repeated measurement cycle to be the most efficient possible. - Choose leaves of different species and individuals according to a reproducible pattern (same height, same exposure, same position within the plant, if possible only from the same node, and only from one category; sun or shade leaves, etc.).
- Only measure leaves in healthy, non-damaged and fully developed condition. Mark the leaves on the individuals (e.g., with cable ties or color tape), to ensure that the repeated measurements are done on the same leaf.
NOTE: Measurements of stomatal conductance should be made only on leaf surface whilst avoiding the midrib and strong leaf veins. - Start measurements in the early morning hours before sunrise taking five to ten repeated measurements until stomatal conductance values shows a clear decline at noon.
NOTE: A daily course of measurements will deliver good data for analyzing the relationships between vpd and stomatal conductance.
- Only measure leaves in healthy, non-damaged and fully developed condition. Mark the leaves on the individuals (e.g., with cable ties or color tape), to ensure that the repeated measurements are done on the same leaf.
- Vpd measurements
- With each gS measurement, record temperature and relative humidity preferably with portable loggers to directly measure the conditions at the position of the same leaf. For calculating the vapor pressure deficit use the August-Roche Magnus formula15.

es=saturation water vapor pressure [hPa]
T=temperature [°C]
- Model gS – vpd response
- Now plot species-wise all gS data against vpd, combining all daily courses of individual leaves into one analysis per species. Extract the maximum value observed from the stomatal conductance data by searching for the maximum value. To scale the model for species-wise comparability, divide the observed values through the maximum value observed for that species (gS/gSMAX).
- For each species, regress the logits of gS (gS/gSMAX) to vpd and the quadratic term of vpd using a generalized linear model with a binomial error distribution (a, b and c represent regression parameters):

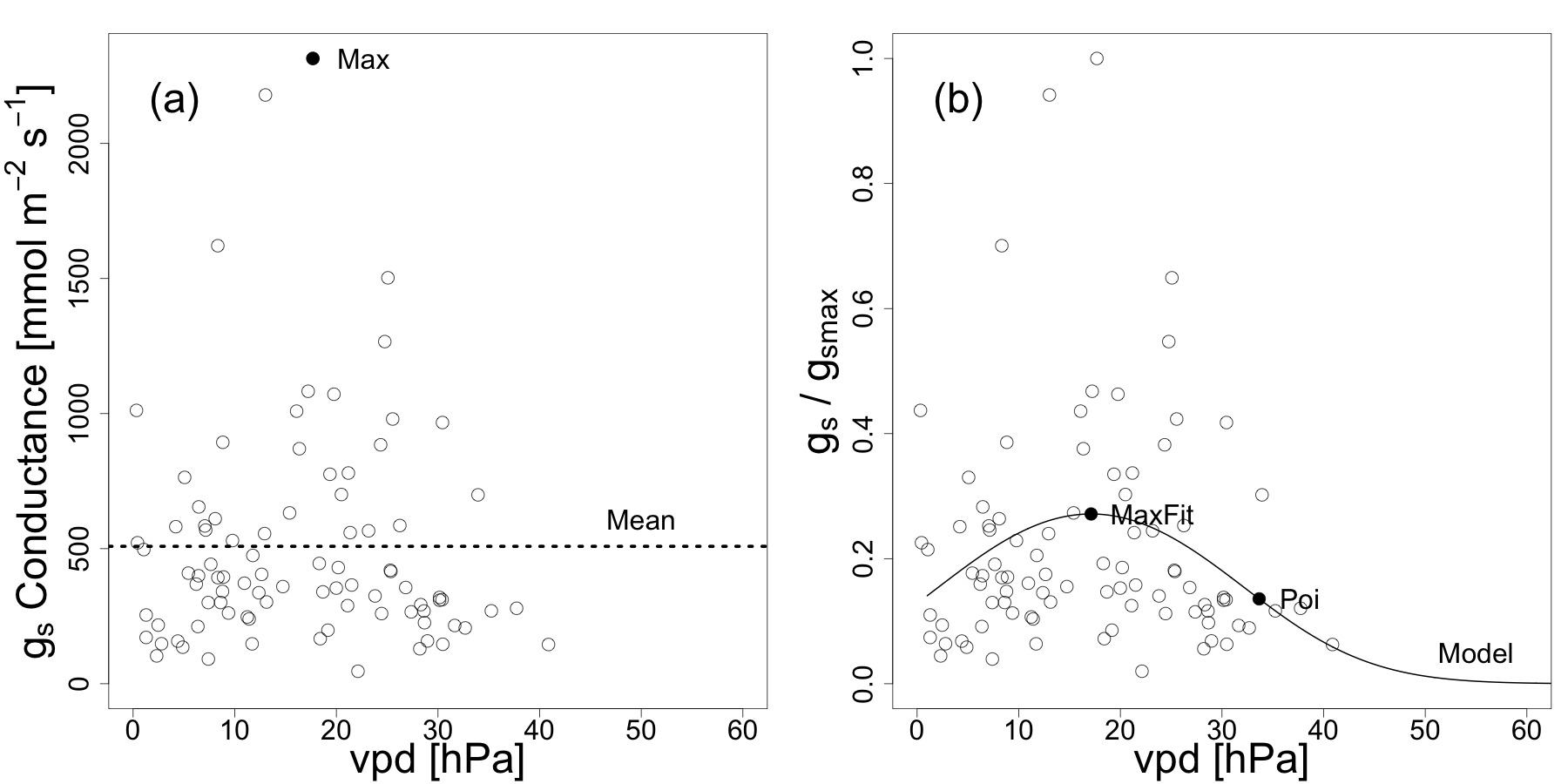
Figure 1. Example of the plotted values and the fitted model for gS – vpd. Stomatal conductance plotted as a function of vapor pressure deficit for the species Liquidambar formosana. Empty dots represent the observed values. (A) Maximum stomatal conductance, the vpd at maximum stomatal conductance and mean stomatal conductance was extracted from the absolute thus non-scaled stomatal conductance (gS) data. (B) Scaled stomatal conductance data (gS/gSMAX) were plotted to extract the relative parameters (shown as filled dots) 20. Re-print with permission from20. Please click here to view a larger version of this figure.

Figure 2. All fitted models for all species. Model graphs for the stomatal conductance data to vpd regression for all species. Evergreen species are represented by black lines, deciduous species by red lines 20. Re-print with permission from20.
NOTE: Using the logits instead of regressing gS directly to vpd, leads to the point that modeled maximum values do not exceed gSMAX and that gS approaches 0 at high vpd.
- Extract parameter of stomatal regulation for every species
- Calculate absolute modeled gSMAX values (MaxFit in Figure 1B). To do this, calculate the vpd at maximum stomatal conductance from setting the first derivative of 1.4.2 to zero, which gives vpdgsMaxFit = -b/2a. Insert vpdgsMaxFit into the formula of 1.4.2 and raise to the power of e to obtain MaxFit. Calculate the mean of all conductance measurements per species (see Figure 1A).
NOTE: Use statistical software R (http://www.r-project.org). - To calculate relative values, from the scaled model (gS/gSMAX), extract stomatal conductance and vpd values for the following two points: (1) stomatal conductance and vpd at the maximum of the model (MaxFit) and (2) vpd at the second point of inflection of the curve (see Figure 1B). Multiply these values by gSMAX to obtain absolute gS values for these points. See Figure 2 for a complete overlay of the single models of all 39 species analyzed.
- Calculate absolute modeled gSMAX values (MaxFit in Figure 1B). To do this, calculate the vpd at maximum stomatal conductance from setting the first derivative of 1.4.2 to zero, which gives vpdgsMaxFit = -b/2a. Insert vpdgsMaxFit into the formula of 1.4.2 and raise to the power of e to obtain MaxFit. Calculate the mean of all conductance measurements per species (see Figure 1A).
2. Measurements of Stomatal Traits
- Take samples preferably from exactly the same leaves that have been used for the measurements of stomatal conductance. If this is not possible, apply the same selection procedure that was applied to choose the leaves for the stomatal conductance measurements, preferably on the same individuals.
- Apply a thin layer of colorless, quickly drying nail polish (test different makes, some are more suitable than others) to a fresh sample. If the samples cannot be processed immediately, store them in 70% alcohol. After the nail polish has dried out, gently peel the impression of from the leaf and proceed for the microscopic analysis, as with a normal leaf sample.
NOTE: In the case of leaves with high trichome density a preceding step of applying sodium hydroxide or a 1:1 solution of acetic acid and oxygen peroxide can lead to better results. - Optical microscope analytics
- Connect a camera to an optical microscope capable of magnifications between 40X to 400X. After the camera is connected to the microscope, match the pictures taken to the optical magnification and resolution of the picture, e.g., with the help of a scale.
- Employing open source image processing software like ImageJ 16 analyze these pictures.
- Draw a shape with the shape tool from the image analysis tool on the image in an area with no dirt, thumb prints, damaged areas, or large leaf veins. Count stomata in this area and in total of at least 50,000 µm2 per sample.
- Measure stomatal guard cell length and pore length. Calculate the number of stomata per mm2. Calculate stomatal index as the ratio of stomatal density to epidermal cell number per mm2.
3. Assessment of Leaf Vein Traits
NOTE: For the assessment of leaf vein traits, a modified a protocol from Sack & Scoffoni 17 was employed.
- Sample preparation
NOTE: To optimize the visibility of the leaf veins, the leaves are first bleached and then stained with safranine and malachite green.- To bleach the leaves, leave them at least 72 hr in 50% solution of decolorizer (test different makes, again some might work better). Alternatively, use 5% NaOH solution or 10% KOH solution or 25% H2O2 solution.
- Heat the solution up to 30° C or combine the different solutions from step 3.1.1 for better effects. Rinse several times in water afterwards. Adapt the bleaching process for the specific species, depending on their leaf characteristics.
NOTE: Thicker leaves may need longer periods of solution bath and or the more aggressive solutions. Thinner and more tender leaves may be bleached to a satisfactorily degree in less than 72 hr. - To color the leaves place them in 100% ethanol. Color them for 2 – 30 min in 1% safranine solution. To enhance coloring, add an additional treatment with 1% malachite green solution only for several seconds. Adapt the protocol to every species in terms of timing and intensity to achieve optimal results. Rinse several times in water afterwards. If the leaves are to deeply stained, some time in ethanol or decolorizer may help.
- Sample analysis
- Scan the leaves with a backlight scanner at a resolution of ca. 1,200 dpi. Match the scans taken to the resolution of the picture, e.g., with the help of a scale, to ensure that pixel length can be traced back to absolute length measures of the scanned leaf.
- Measure area, circumference, length and width of the leaves. Calculate several indices, e.g., length/width and circumference2/area. Cut a 1 by 1 cm rectangle out of the middle of the picture. Measure diameter of the veins of first and second order (do not include the main middle vein). Measure the length of all veins of first order in this quadrat (vein density).
4. Assessment of Other Leaf Traits
NOTE: Asses typical leaf traits such as specific leaf area (SLA), leaf dry matter content (LDMC), leaf area, element contents, leaf habit, leaf pinnation, leaf compound type, leaf margin type etc. following established protocols 18,19.
- Observational leaf traits
- Assess leaf pinnation, leaf compound type, leaf margin type and the presence of extrafloral nectaries18,19 through observation in the field.
- Analytical leaf traits
- Collect fresh leaf samples, preferably on the same individuals as used for the other measurements, for the determination of specific leaf area (SLA), leaf area and leaf dry matter content (LDMC) 18,19. After 48 hr in the drying oven at 80 °C, measure element contents and ratios, preferably on the same leaves.
Many parameters of stomatal conductance and stomatal regulation were found to be related to morphological, anatomical and chemical leaf traits. In the following, the focus will be on the links to vpd at the point of inflection, which decreased with stomatal density (p=0.04) and stomatal index (p=0.03) and increased with leaf carbon content (p=0.02, See Figure 3). The results show that with decreasing vpd at the point of inflection there was a decrease in stomatal density and stomatal index. In contrast, no parameter of stomatal conductance showed a clear relationship to leaf habit. The high variation within the two groups of leaf habit shows that different regulatory mechanisms exist both within the group of evergreen and deciduous leaf habits.

Figure 3. Result figure for the links between patterns of stomatal conductance regulation and leaf traits. Vapor pressure deficit (vpd) at the point of inflection of the gS – vpd curve (VpdPoi) as a function of (A) stomata density, (B) stomata index, and (C) leaf carbon content 20. Re-print with permission from20. Please click here to view a larger version of this figure.
