A Lipid Extraction and Analysis Method for Characterizing Soil Microbes in Experiments with Many Samples
Summary
The article describes a method that increases throughput while balancing effort and accuracy for extraction of lipids from the cell membranes of microorganisms for use in characterizing both total lipids and the relative abundance of indicator lipids to determine soil microbial community structure in studies with many samples.
Abstract
Microbial communities are important drivers and regulators of ecosystem processes. To understand how management of ecosystems may affect microbial communities, a relatively precise but effort-intensive technique to assay microbial community composition is phospholipid fatty acid (PLFA) analysis. PLFA was developed to analyze phospholipid biomarkers, which can be used as indicators of microbial biomass and the composition of broad functional groups of fungi and bacteria. It has commonly been used to compare soils under alternative plant communities, ecology, and management regimes. The PLFA method has been shown to be sensitive to detecting shifts in microbial community composition.
An alternative method, fatty acid methyl ester extraction and analysis (MIDI-FA) was developed for rapid extraction of total lipids, without separation of the phospholipid fraction, from pure cultures as a microbial identification technique. This method is rapid but is less suited for soil samples because it lacks an initial step separating soil particles and begins instead with a saponification reaction that likely produces artifacts from the background organic matter in the soil.
This article describes a method that increases throughput while balancing effort and accuracy for extraction of lipids from the cell membranes of microorganisms for use in characterizing both total lipids and the relative abundance of indicator lipids to determine soil microbial community structure in studies with many samples. The method combines the accuracy achieved through PLFA profiling by extracting and concentrating soil lipids as a first step, and a reduction in effort by saponifying the organic material extracted and processing with the MIDI-FA method as a second step.
Introduction
Given the key role of microorganisms in nutrient cycling 1, modification of plant community composition 2, regulation of plant productivity 3, and decomposition of organic matter 4, understanding soil microbial communities is a vital to understanding terrestrial ecosystems.
Because of their relatively high abundance in soil, and their chemical signature, lipid biomarkers can be used to profile the dominant ecological groups comprising soil microbial communities 5. By quantifying lipid biomarkers that are characteristic of different microbial groups, we can estimate total lipids, and then separate these lipids into ecologically relevant groups such as Gram positive (Gm+) and Gram negative (Gm-) bacteria, arbuscular mycorrhizal (AM) and saprotrophic fungi, and actinomycetes 5,6,7,8.
There are many methods for characterizing aspects of microbial communities. The PLFA method is one commonly used to understand basic microbial community structure. It is an effective way to assess the relative abundance of microbial groups as well as total microbial biomass. Due to rapid lipid turnover, PLFA profiling also allows relatively fast detection of changes in the soil microbial community and gives information that allows comparison of ecosystem function, e.g., fungal:bacterial ratios to assess rates of nutrient cycling 1,9,10. However, while the PLFA extraction method is time honored and well respected, it is also time consuming and does not lend itself well to ecosystem-scale studies that require a large number of samples from field scale replicates11,12.
In contrast, the fatty acid methyl ester extraction method (MIDI-FA) has the potential to allow rapid throughput. In this method, samples are saponified, converted to FAMEs, extracted, and then analyzed. The MIDI-FA method is rapid but less discriminating than PLFA, which combines extraction of lipids with separation of different lipid classes13 (phospholipids, neutral lipids, and glycolipids).
In this protocol, we describe a method that combines elements of both PLFA and MIDI-FA lipid profiling. It employs extraction of lipids using the initial chloroform extraction steps of the modified Bligh and Dyer method, and then saponification and conversion to FAMEs. This provides a robust way to detect microbial community structure while excluding much of the background noise from non-microbial material 5,14. This method was developed to achieve a balance between both the PLFA and MIDI-FA protocols, i.e., retain most of the accuracy while increasing throughput to make it logistically and economically feasible to analyze lipids from large-scale studies with many samples15. By performing the initial extraction and isolating the organic-soluble components (e.g. lipids) prior to performing MIDI-FA, and completing this with a purification step, the protocol offers a balance between speed and precision.
Protocol
NOTE: Always wear appropriate personal protective equipment (PPE) throughout the procedure. To avoid potential sample contamination, do not touch glassware with bare hands. Wear appropriate gloves when running protocol steps that require handling of chloroform.
1. Preparations (2 Days for ~40 Samples)
- Collect soil into sterile bags and transport from the field in a cooler containing ice. If it is not possible to sieve fresh soil and freeze dry immediately, store the samples in a -80 °C freezer until ready to start the analysis.
- Prepare soil by homogenizing. Remove roots and stones and break up clods by coarse sieving (e.g. 2 mm).
- Prepare for freeze drying by putting subsamples in an appropriate container as per instructions of the freeze dryer manual and freeze dry as soon as possible. Once soils are freeze dried, store in a sealed container with desiccant until extraction. It is best to store freeze dried soil at a minimum of -20 °C but preferably at -80 °C.
- In preparation for extraction, remove freeze dried soils from storage and grind to a flour-like consistency. Methods for grinding include ball mill, ball beater, and mortar and pestle. After grinding soil, store in a freezer (see 1.3).
NOTE: The amount of sample used for extraction depends on its organic matter content. A general guideline is to use 0.5 to 1 g of a soil that is 12 to 18% carbon by weight, and 3 to 5 g for a soil 1 to 3% carbon by weight. - Pre-rinse 30 mL centrifuge tubes with hexane. Add approximately 2 to 3 mL of hexane to tubes and vortex for 5 sec. Decant hexane to another tube, and vortex. Hexane (2 to 3 mL) can be used to serially rinse six tubes. Store hexane-rinsed tubes inverted in the fume hood and dispose of used hexane in an appropriate waste container.
- Wrap the glassware in 2 to 3 layers of aluminum foil and place in the muffle furnace. Bake (muffle) glassware at 450 °C for 4.5 h.
- Prepare reagents.
- For the soil amount used, add chemicals in a 0.8:1:2 volume ratio for phosphate-buffer (P-buffer):CHCl3:MeOH.
- Prepare P-buffer solution: Phosphate-Buffer: 0.1 M, pH 7.0.
- Add 61 mL of 1M K2HPO4 stock, sterile (chemical should be ACS certified grade or better). Add 39 mL of 1 M KH2PO4 stock, sterile (chemical should be ACS certified grade or better). Fill to 1000 mL with Type 1 water. Adjust pH to 7.0 with NaOH or HCl. Store unused solution for up to 7 d at ambient temperature or 30 days in a refrigerator.
- Alternatively, weigh out 10.62 g of K2HPO4 and 5.31 g of KH2PO4; dilute to 1 L with Type 1 water. Check pH, adjust if necessary.
- Prepare Reagent 1, Saponification Reagent.
- Dispense 300 mL of Type 1 water. Add 300 mL of CH3OH (HPLC grade or higher). Add 90.00 g of NaOH (certified ACS or better). Add NaOH pellets to the solution while stirring. Stir until the pellets dissolve. It is recommended to store this solution no longer than 30 d.
- Prepare Reagent 2, Methylation Reagent.
- Dispense 275 mL of CH3OH (HPLC grade or higher). Add 325 mL of certified 6.00 N HCL while stirring. It is recommended to store this solution no longer than 30 d.
- Prepare Reagent 3, Extraction Reagent: 50 % C6H14 (Hexane), 50% C5H12O (MTBE).
- In the fume hood, combine 200 mL of C6H14 (HPLC grade or higher) and 200 mL of C5H12O (HPLC grade or higher). Mix well; cap tightly. Store in flammables cabinet or fume hood for no longer than 1 year.
- Prepare Reagent 4, Base wash Reagent.
- Dispense 900 mL of Type 1 water to beaker. Add 10.80 g of NaOH (Certified ACS or better) while stirring. Stir until the pellets dissolve. It is recommended to store this solution no longer than 30 d.
- Prepare stock solution of internal standard.
- Weigh out 100 mg of 19:0 EE standard. Add to 50 mL of hexane-rinsed volumetric flask. Add ~15 mL of Hexane-MTBE solution (reagent 3) to flask, swirl to dissolve. Top off to 50 mL with Hexane-MTBE solution (reagent 3). Cap and swirl to mix. Transfer to hexane-rinsed glass tubes with PTFE lined caps and store at -20 °C.
2. DAY 1 – Extraction of Fatty Acids from Soil (4 to 5 h for ~40 Samples)
- Prepare three 5 to 10 mL dispensing pipettes for P-buffer, chloroform, and methanol.
NOTE: Less hazardous alternatives to chloroform have been proposed as extractants. These may result in equivalent lipid recovery rates 16,17, but are not evaluated in this protocol. - Weigh freeze-dried soils into 30 mL centrifuge tubes and record soil mass.
- Make two blanks and include a check standard (a soil that has been extracted previously).
- In the fume hood, add the reagents to the soil in the centrifuge tube in the following order: P-buffer, CHCl3, and MeOH (Table 1). Allow the soils time to wet after the P-buffer addition before adding the CHCl3. Cap the centrifuge tubes tightly and cover to protect from light.
- Place them on the shaker horizontally making sure they are well-secured. With the speed setting on 280 rpm, shake for 1 h18.
- Prepare two 16 mm x 150 mm glass tubes for each sample as follows: Label the tube and add the same volume of CHCl3 and an equal volume of P-buffer.
- Remove the centrifuge tubes from the shaker and centrifuge for 10 min at 1,430 x g and 25 °C. Phase separation should be visible in the glass tube.
- In the fume hood, decant the supernatant from the centrifuge tube into one of tubes prepared in step 2.6. Repeat steps 2.4 through 2.5 and decant the supernatant into the second tube.
- Securely cap all the16 mm × 150 mm glass tubes with PTFE-lined caps and invert 10 x to mix.
- Allow the samples to stand undisturbed overnight to complete separation of the two phases. To do this, keep the samples in a dark cabinet/space or covered in aluminum foil at room temperature. It is acceptable to allow the extracts to separate over the weekend.
- Alternatively, if one desires to move directly to the next step, or if samples get disturbed the next day, centrifuge samples for 10 min at 1,000 x g and 25 °C.
NOTE: Subject all samples within a comparison group to the same phase separation standing time.
- Alternatively, if one desires to move directly to the next step, or if samples get disturbed the next day, centrifuge samples for 10 min at 1,000 x g and 25 °C.
3. DAY 2 – Isolation of Lipids (3 to 4 h for ~40 Samples)
- Turn on evaporation apparatus and solvent trap. Set evaporation apparatus to 33 °C and preheat.
- Set up a vacuum aspirator in the hood: a side-arm flask connected to a vacuum pump, a length of high purity tubing and a glass pipet.
- Aspirate the top layer and interface (approximately 2/3 of the way down) of the two glass tubes, retaining the aqueous layer. Repeat the process using a clean pipet for each sample.
- Combine the aqueous layers of extract from the second tube with that in the first tube by carefully decanting. Ensure that the tube being decanting is free of scratches or cracks.
- Once the CHCl3 extracts are combined, inspect the liquid – it should be clear. If not add MeOH until clear.
- Dry down all the samples using the vacuum evaporation apparatus. Start with settings of temperature 33 °C, vortex speed 26%, and pressure 400 mbar.
- Place the samples in the evaporation system and after 10 min slowly lower the pressure to 350 mbar.
- Monitor the progress and when the liquid level in the tube has reached the level of the heating block, decrease the pressure to 300 mbar.
- Continue to monitor the height of the remaining liquid and when it is approximately 1 cm in depth, decrease the pressure to 200 mbar.
- After samples have dried, remove samples and run the pump for 10 additional min to clear vapors.
- Cap the tubes tightly and store the lipids in a freezer at -80 °C.
- Dispose of the aspirated liquid in accordance with applicable chemical safety regulations.
4. DAY 3 – Saponification and Methylation (6 to 7 h for ~40 Samples)
- Turn on the water baths. Set bath 1 to 95 °C and bath 2 to 80 °C.
- Set the pipet dispensers and ensure they are dispensing the correct volume.
Reagent 1: 1 mL per sample
Reagent 2: 2 mL per sample
Reagent 3: 1.25 mL per sample
Reagent 4: 3 mL per sample
NOTE: The heating process will build pressure in the tube. Do not use scratched, cracked, or chipped tubes. - Using a pipet dispenser add 1.0 mL of Reagent 1 to the dried lipids. Cap tightly, vortex for 5 s and place in a rack.
- Place the rack of sample tubes into the 95 °C water bath for 5min. Remove the rack of tubes from the bath and check the tubes for leaks, indicated by bubbles rising in the tube. Retighten or replace the caps of leaking tubes, and check again for cracks or chips. Continue heating the tubes in the water bath for an additional 10 min.
- Remove samples from bath 1 and place in bath 2 (80 °C) and continue the incubation for another 15 min.
- Remove the tubes and cool the samples by placing the rack into a pan of tap water for cooling.
- Add 2 mL of Reagent 2 to each sample. Cap tightly and vortex for 5 to 10s.
- Place the rack into the 80 °C water bath and incubate for 10 min.
- Remove the rack of tubes from the water bath and put into a pan of tap water for cooling. Agitate the rack of tubes to accelerate the cooling process.
NOTE: Do not exceed time and temperature. Too much heating could degrade FAMEs. - Using the pipet dispenser add 1.25 mL of Reagent 3 to each sample tube to extract the fatty acid methyl esters. Cap tightly and put the tubes on the shaker for 10 min.
- After shaking, allow the rack of tubes sit for 10 min for the phases to separate. Transfer the organic phase (top layer) to a 16 mm x 100 mm glass test tube using a glass pipet.
- Repeat the extraction by again adding Reagent 3, shaking, allowing phases to separate, and transferring the top phase.
NOTE Do not transfer any of the bottom-phase. It is fine to leave a small amount of top phase in the tube. - Using the pipet dispenser, add 3 mL of Reagent 4 to the 16 mm x 100 mm tubes.
- Cap the test tubes tightly and vortex for 20-30 s.
- After vortexing, centrifuge for 3 min at 1,000 × g.
- Using a clean glass pipet, aspirate the top organic phase and transfer to a 4- mL amber vial.
- Turn on evaporation apparatus and solvent trap. Set evaporation apparatus to 30 °C, the vortex speed to 26%, the vacuum to 200 mbar, and press preheat.
- Monitor the progress. After samples have dried, remove samples and run the pump for 10 additional min to clear vapors.
- Remove pipet from the reagent bottles 1 and 4 and pump through some dilute acid (e.g. 1% HCl), followed by D.I. water. Rinse the pipet used for Reagent 2 by pumping through D.I. water. In the hood, drain the solvent from the pipet used for Reagent 3 into an appropriate container and keep it in the hood until the solvent residues have evaporated.
- Store the pipets inverted, with the plungers removed to prevent sticking of the check valves.
- Use glass pipets once and then dispose in an appropriate container.
- In the hood, allow solvent residues on glassware to evaporate.
- Rinse glassware with clean water and detergent solution.
5. DAY 4 – Preparation of the Working Solution and Transfer of FAMEs to the GC Vials (2-3 h for ~40 Samples)
- Gather materials: 4 mL amber vials with dried samples; amber 2 mL GC vials; 400 µL flat bottom glass inserts and caps; 500 µL glass syringe; 50 mL volumetric flask; Stock solution – Ethyl nonadecanoate (19:0 EE internal standard) in 50/50 hexane/MTBE (Reagent 3).
- Using the glass syringe, add 500 µL of ethyl nonadecanoate (19:0 EE) stock solution to a 50-mL volumetric flask.
- Fill flask to volumetric score with Reagent 3.
- Cap flask and invert 5x to mix.
- Transfer 3 mL to a clean 4 mL vial for a working solution reservoir.
- Using the glass syringe, add 300 µL of the working solution to each of the 4 mL vials containing the dried fatty acid methyl esters and cap.
- Vortex the sample for 15 s and set aside to stand for 15 min.
- Using a glass Pasteur pipet, carefully transfer the suspended fatty acid methyl esters to a 2 mL GC vial containing the 400 µL insert.
- Store sealed GC vials in a -20 °C freezer prior to analysis.
- Submit samples for GC analysis.
NOTE: The analysis must be carried out using a specific GC column and conditions which are described in Supplementary File S1. It is best that the GC analysis be completed within 2 weeks of methylation.
Representative Results
The data tables from the reports can be collated into a spreadsheet or database. After adjusting for response factor (a correction factor that normalizes the response for different chain lengths), peak areas can be compared with the peak area of external or internal standards to arrive at a concentration in the extract. By dividing through by the mass of soil extracted, the data can be expressed as mass of FAME per gram of soil or, by using the molecular weight of each FAME, the more commonly reported nmol per gram of soil. The sum of microbial FAMEs is indicative of total microbial biomass, and can be compared among treatments (Figure 1). Certain FAMEs can also be associated with particular microbial groups such as Gram-positive or Gram-negative bacteria, actinomycetes, arbuscular mycorrhizal fungi, and saprotrophic fungi 19,20,21,22,23,24. Ratios of the mass of specific biomarkers can reflect the relative abundance of these groups (Figure 2). Overall FAME abundance patterns create community-level fingerprints, which allow comparison of microbial community dissimilarity through multivariate techniques such as ordination (Figure 3). In contrast with most DNA-based approaches, community-level lipid data can be analyzed either as relative or absolute abundances. If total biomass differs substantially among samples, these two approaches will give very different results; the ecological questions underlying the experiment should determine which approach is used.
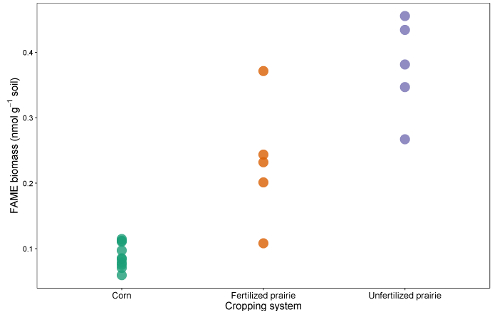
Figure 1: Total FAME biomass. Comparison of total FAME biomarker biomass (nmol g-1 soil) from continuous corn, fertilized prairie, and unfertilized prairie. Please click here to view a larger version of this figure.
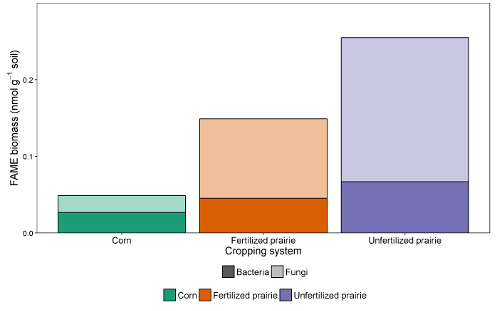
Figure 2: FAME biomass of bacteria and fungi. Treatment comparison of bacterial and fungal FAME biomarker biomass (nmol g-1 soil) from continuous corn, fertilized prairie, and unfertilized prairie.Fungal and bacterial mass from absolute abundance. Average (sum f/sum b). Please click here to view a larger version of this figure.
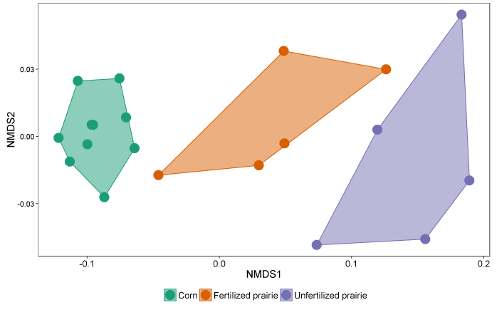
Figure 3: Non-metric Dimensional Scaling of FAME lipid biomarkers. Comparison of overall microbial community using absolute abundance profiles of all microbial FAME lipid biomarkers. In this example, continuous corn and unfertilized prairie communities separated and are very far apart while some fertilized prairie samples have microbial communities that resemble those from corn, and others resemble the unfertilized prairie. Please click here to view a larger version of this figure.
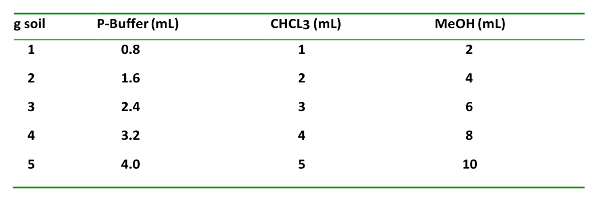
Table 1: The solvent type, proportions added g-1 soil, and their order of addition. This is important for proper extraction and separation of the organic and aqueous phases.
Supplementary File S1: Please click here to download.
Discussion
For the examination of multiple samples from experiments with many replicates and/or experimental units, researchers may find phospholipid fatty-acid analysis (PLFA) to be prohibitive in terms of time and materials25. With the PLFA method, cell membrane phospholipids are extracted, purified, and identified using the modified Bligh and Dyer26 two-phase aqueous-organic extraction. This is followed by solid-phase silica chromatography to separate lipids by polarity, and an alkaline methylation of phospholipid fatty acids into fatty acid methyl esters. In PLFA profiling lipid yield can be low but of a very high purity. Microbial ID introduced an alternate method, the fatty acid methyl-ester procedure (MIDI-FA). In the MIDI-FA method, all lipids are extracted directly from pure cultures or soil/sediment samples11,12,27 through saponification. This method has lower lipid loss and is rapid because it has none of the concentration or purification steps of the PLFA method. However, while the MIDI-FA method is quick and less expensive, because it was originally designed for identifying organisms in pure culture, there are no initial extraction or purification steps. Thus, it can include lipid-like compounds co-extracted from soil organic matter that distort the community signature27,28,29. Because this inclusion can also distort biomass measures, MIDI-FA has typically been used only to coarsely qualitatively describe soil lipids13.
The procedure we describe here combines the best of the two separate extraction procedures: 1) an extraction and concentration of lipids using the modified Bligh and Dyer26 method, and 2) the fatty acid methyl ester saponification, methylation, extraction, and base wash procedure developed commercially. This method was developed to achieve the benefits of both of these protocols while minimizing the disadvantages15. By performing the initial extraction and isolating the organic-soluble components (e.g. lipids) prior to performing MIDI-FA, and completing this with a purification step, this protocol offers a balance between speed and precision. Although this method may not be suitable when high purity is required (i.e., for 13C PLFA analysis) or when analyzing phospholipids and neutral lipids separately, in many cases it allows detection of microbial community responses to environmental conditions with greater sensitivity than DNA-based methods30,31,32,33. Membrane lipids decompose rapidly after cell death allowing them to reflect the living microbial community at the time of sampling5,7, in contrast with environmental DNA in which much of the information comes from dead or inactive organisms34. Given the high rate of dormancy observed among soil microorganisms35, the characterization of live biomass can be used to understand temporal plant-microbe interactions at a relatively fine temporal scale and lipid biomarkers can be used to assay the physiological status of the microbial community7. It has been shown that high throughput methods are required to assess microbial response in large field settings25, and while the method we propose here does not replicate the accuracy of PLFA biomarker profiling, it increases throughput while minimizing the variability realized with the MIDI-FA procedure. The method has proven to be an effective tool in addressing questions related to microbial community dynamics on a wide range of soils in large-scale agricultural and ecosystem studies36,37,38,42,43,44,45,46,47,48,49,50.
Lipid classes are combined with this method and there may be a loss of the information contained in those separate classes22,39, but combining the lipid classes may strengthen the power to detect the arbuscular mycorrhizal fungi origin of the 16:1 ω5c from both phospho- and neutral-lipids40. In addition, while the number of unknown fatty acids (which could be derived from non-living organic matter) may be higher with this method, it was shown to be lower than MIDI-FA and allows for treatment comparison of lipid profiles from studies with many samples where sample throughput capacity is a concern15. The neutral fraction is generally thought to derive primarily from storage lipids produced by fungi, though there may also be some smaller contribution from soil fauna41. In light of this, the method described here may yield results showing a greater contribution of the fungal lipids 18:2 ω 6,9c and 18:1 ω 9c than PLFA. The other lipids that tend to show up in the neutral fraction include some of the saturated fatty acids, e.g. 16:0, 18:0, 20:0.
There are different ways in which lipid data can be expressed and analyzed. The most common representations are abundance (nmol g-1 soil), mole fraction (nmol individual lipid nmol-1 total lipid), and mole percent (mole fraction*100). Normalized by the total lipids in a sample, mole fraction and mole percent are measures of the relative abundance of a given lipid. After an appropriate transformation, e.g., arcsine square-root, mole fraction is appropriate for use in analysis by principal components or redundancy analysis ordination. Abundance is the absolute amount of a given lipid extracted per gram of soil. Because the quantity of lipid per cell is reasonably constant, and the lipid extraction is highly efficient and comprehensive, total abundance is a good estimate of total lipids and the abundance of key indicators reflects the biomass of the ecological group it represents17. Finally, a good way to look at microbial community composition is to use multivariate analysis methods16, e.g., ordination methods such as nonmetric multidimensional scaling (NMDS – which does not need data transformation) or principal components analysis (PCA), can be useful for comparing the relative abundance of all lipid biomarkers.
Disclosures
The authors have nothing to disclose.
Acknowledgements
This work was funded in part by the DOE Great Lakes Bioenergy Research Center (DOE BER Office of Science DE-FC02-07ER64494) and DOE OBP Office of Energy Efficiency and Renewable Energy (DE-AC05-76RL01830). The authors would like to acknowledge Anders Gurda for his patient and skillful contribution to videography and editing.
Materials
| RapidVap | Labconco | 7900000 | Evaporative system |
| RapidVap | Labconco | 7491400 | Sample block |
| Chem-resistant vacuum pump | Labconco | 7584000 | Vacuum pump |
| Chemical trap cannister | Labconco | 7815300 | Trap cannister |
| Solvent insert | Labconco | 7515200 | Solvent trap insert |
| Diaphragm pump | Welch | 7398000 | DryFast vacuum pump |
| Water bath | Fisher | 15-462-15Q | 2 well water bath |
| Gas chromatograph | Various | N/A | For sample analysis |
| Centrifuge | Various | N/A | For sample separation |
| Freeze Dryer | Various | N/A | For sample lyophilization |
| Repipet (6) | BrandTech | 4720440 | For dispensing reagents |
| Vortex | Fisher | 02-215-365 | Analog vortex mixer |
| Teflon centrifuge tubes | Thermo/Nalgene | 3114-0030 | Teflon sample tubes |
| Caps | Thermo/Nalgene | DS3131-0020 | Caps for teflon tubes |
| Test tube | Corning | 9825-16 | 16x100mm tubes |
| Test tube | Corning | 9825-16xx | 16x150mm tubes |
| Caps | Corning | 9998-15 | 15-415 thread black phenolic caps w/PTFE liner |
| Pasteur pipets | Fisher | 13-678-20B | 14.6cm |
| 500 uL glass syringe | Fisher | 13684106LC | Hamilton 81217 |
| Amber vials | Agilent | 5182-0716 | 4mL Amber vials |
| Caps | Agilent | 5182-0717 | Blue screw caps |
| Inserts | Agilent | 5181-3377 | 400uL flat bottom glass inserts |
| Standards | |||
| 19:0 ethyl nonadecanoate (Ethyl nonadecanoate) | VWR | TCN0459-5G | Analytical standard |
| Chemicals | |||
| Dipotassium phosphate (K2HPO4 – ACS grade or better) | Various | N/A | For making Phosphate-Buffer |
| Potassium phosphate monobasic (KH2PO4 – ACS grade or better) | Various | N/A | For making Phosphate-Buffer |
| Methanol (CH3OH – HPLC grade or better) | Various | N/A | For making Reagents 1 & 2 |
| Sodium hydroxide (NaOH – ACS grade or better) | Various | N/A | For making Reagents 1 & 4 |
| Hydrochloric acid (6N HCL) | Various | N/A | For making Reagent 2 |
| Hexane (HPLC grade or better) | Various | N/A | For making Reagent 3 |
| MTBE (Methyl tert-butyl ether – HPLC grade or better) | Various | N/A | For making Reagent 3 |
References
- Van Der Heijden, M. G. a., Bardgett, R. D., Van Straalen, N. M. The unseen majority: Soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems. Ecol Lett. 11 (3), 296-310 (2008).
- Bever, J. D., et al. Rooting theories of plant community ecology in microbial interactions. Trends Ecol Evol. 25 (8), 468-478 (2010).
- Suleiman, A. K. A., Manoeli, L., Boldo, J. T., Pereira, M. G., Roesch, L. F. W. Shifts in soil bacterial community after eight years of land-use change. Syst Appl Microbiol. 36 (2), 137-144 (2013).
- Sayer, E. J., et al. Grassland management influences spatial patterns of soil microbial communities. Soil Biol Biochem. 61, 61-68 (2013).
- Fernandes, M. F., Saxena, J., Dick, R. P. Comparison of Whole-Cell Fatty Acid (MIDI) or Phospholipid Fatty Acid (PLFA) Extractants as Biomarkers to Profile Soil Microbial Communities. Microbial Ecol. 66 (1), 145-157 (2013).
- van der Heijden, M. G. A., Wagg, C. Soil microbial diversity and agro-ecosystem functioning. Plant Soil. 363 (1-2), 1-5 (2013).
- Allison, V. J., Miller, R. M., Jastrow, J. D., Matamala, R., Zak, D. R. Changes in soil microbial community structure in a tallgrass prairie chronosequence. Soil Sci Soc Am J. 69 (5), 1412-1421 (2005).
- Kowalchuk, G. A., Buma, D. S., de Boer, W., Klinkhamer, P. G. L., van Veen, J. A. Effects of above-ground plant species composition and diversity on the diversity of soil-borne microorganisms. A van Leeuw. 81 (1-4), 509-520 (2002).
- Bardgett, R. D., Hobbs, P. J., Frostegard, A. Changes in soil fungal:bacterial biomass ratios following reductions in the intensity of management of an upland grassland. Biol Fert Soils. 22 (3), 261-264 (1996).
- Frostegård, &. #. 1. 9. 7. ;., Tunlid, A., Bååth, E. Use and misuse of PLFA measurements in soils. Soil Biol Biochem. 43 (8), 1621-1625 (2011).
- Haack, S. K., Garchow, I. H., Odelson, D. A., Forney, L. J., Klug, M. J. Accuracy , Reproducibility , and Interpretation of Fatty Acid Methyl Ester Profiles of Model Bacterial Communitiest. Appl Environ Microbiol. 60 (7), 2483-2493 (1994).
- Drenovsky, R. E., Elliott, G. N., Graham, K. J., Scow, K. M. Comparison of phospholipid fatty acid ( PLFA ) and total soil fatty acid methyl esters ( TSFAME ) for characterizing soil microbial communities. Soil Biol Biochem. 36, 1793-1800 (2004).
- Cavigelli, M. A., Robertson, G. P., Klug, M. Fatty acid methyl ester ( FAME ) profiles as measures of soil microbial community structure. Plant Soil. 170, 99-113 (1995).
- Determination of microbial community structure using phospholipid fatty acid profiles. Molecular microbial ecology manual Available from: https://www.researchgate.net/profile/Robert_Findlay2/publication/281208148_Determination_of_microbial_community_structure_using_phospholipid_fatty_acid_profiles/links/55db4f0b08aec156b9afe776.pdf (2004)
- Balser, T. C., Liang, C., Gutknecht, J. L. Linking microbial community analysis and ecosystem studies: A rapid lipd analysis protocol for high throughput. Biol Fert Soils. , (2017).
- Findlay, R. H., King, G. M., Watling, L., Watling, L. E. S. Efficacy of Phospholipid Analysis in Determining Microbial Biomass in Sediments Efficacy of Phospholipid Analysis in Determining Microbial Biomass in Sedimentst. Appl Environ Microbiol. 55 (11), 2888-2893 (1989).
- Willers, C., Jansen van Rensburg, P. J., Claassens, S. Microbial signature lipid biomarker analysis – an approach that is still preferred, even amid various method modifications. J Appl Microbiol. 118 (6), 1251-1263 (2015).
- Zelles, L., Bai, Q. Y., Beck, T., Beese, F. Signature fatty-acids in phospholipids and lipopolysachharides as indicators of microbial biomass and community structure in agricultural soils. Soil Biol Biochem. 24 (4), 317-323 (1992).
- Frostegard, A., Tunlid, A., Baath, E. Phospholipid fatty-acid composition, biomass, and activity of microbial communities from 2 soil types experimentally exposed to different heavy-metals. Appl Environ Microbiol. 59 (11), 3605-3617 (1993).
- Federle, T. W., Dobbins, D. C., Thorntonmanning, J. R., Jones, D. D. Microbial biomass, activity, and community structure in subsurface soils. Ground Water. 24 (3), 365-374 (1986).
- Vestal, J. R., White, D. C. Lipid analysis in microbial ecology – quantitative approaches to the study of microbial communities. BioScience. 39 (8), 535-541 (1989).
- Wilkinson, S. G. Gram negative bacteria. Microbial Lipids. , 299-488 (1988).
- Balser, T. C., Treseder, K., Ekenler, M. Using lipid analysis and hyphal length to quantify AM and saprotrophic fungal abundance along a soil chronosequence. Soil Biol Biochem. 37 (3), 601-604 (2005).
- Buyer, J. S., Sasser, M. High throughput phospholipid fatty acid analysis of soils. Appl Soil Ecol. 61, 127-130 (2012).
- Bligh, E. G., Dyer, W. J. A rapid method of total lipid extraction and purification. Can J Biochem Phys. (37), 911-917 (1959).
- Schutter, M. E., Dick, R. P. Comparison of Fatty Acid Methyl Ester (FAME) Methods for Characterizing Microbial Communities. Soil Sci Soc Am J. 64 (5), 1659-1668 (2000).
- Jandl, G., Leinweber, P., Schulten, H., Ekschmitt, K. Contribution of primary organic matter to the fatty acid pool in agricultural soils. Soil Biol Biochem. 37, 1033-1041 (2005).
- Nielsen, P., Petersen, S. O. Ester-linked polar lipid fatty acid profiles of soil microbial communities: a comparison of extraction methods and evaluation of interference from humic acids. Soil Biol Biochem. 32, 1241-1249 (2000).
- Duncan, D. S., Jewell, K. A., Suen, G., Jackson, R. D. Detection of short-term cropping system-induced changes to soil bacterial communities differs among four molecular characterization methods. Soil Biol Biochem. 96, 160-168 (2016).
- Liang, C., et al. Switchgrass rhizospheres stimulate microbial biomass but deplete microbial necromass in agricultural soils of the upper Midwest, USA. Soil Biol Biochem. 94, 173-180 (2016).
- Jangid, K., et al. Land-use history has a stronger impact on soil microbial community composition than aboveground vegetation and soil properties. Soil Biol Biochem. 43 (10), 2184-2193 (2011).
- Ritz, K., et al. Spatial structure in soil chemical and microbiological properties in an upland grassland. FEMS Microbiol Ecol. 49 (2), 191-205 (2004).
- Carini, P., et al. Relic DNA is abundant in soil and obscures estimates of soil microbial diversity. J Chem Inf Model. 53 (9), 1689-1699 (2013).
- Lennon, J. T., Jones, S. E. Microbial seed banks: The ecological and evolutionary implications of dormancy. Nat Rev Microbiol. 9 (2), 119-130 (2011).
- Gutknecht, J. L. M., Field, C. B., Balser, T. C. Microbial communities and their responses to simulated global change fluctuate greatly over multiple years. Glob Change Biol. 18, 225-269 (2012).
- Pei, Z., et al. Soil and tree species traits both shape soil microbial communities during early growth of Chinese subtropical forests. Soil Biol Biochem. 96, 180-190 (2016).
- Oates, L. G., Duncan, D. S., Sanford, G. R., Liang, C., Jackson, R. D. Bioenergy cropping systems that incorporate native grasses stimulate growth of plant-associated soil microbes in the absence of nitrogen fertilization. Ag Ecosys Environ. 233, 396-403 (2016).
- Bååth, E. The use of neutral lipid fatty acids to indicate the physiological conditions of soil fungi. Microbial Ecol. 45 (4), 373-383 (2003).
- Ngosong, C., Gabriel, E., Ruess, L. Use of the signature Fatty Acid 16:1ω5 as a tool to determine the distribution of arbuscular mycorrhizal fungi in soil. J Lipids. 2012, 236807 (2012).
- Sharma, M. P., Buyer, J. S. Comparison of biochemical and microscopic methods for quantification of arbuscular mycorrhizal fungi in soil and roots. Appl Soil Ecol. 95, 86-89 (2015).
- Oates, L. G., Balser, T. C., Jackson, R. D. Subhumid pasture soil microbial communities affected by presence of grazing, but not grazing management. Appl Soil Ecol. 59, 20-28 (2012).
- Liang, C. Potential legacy effects of biofuel cropping systems on soil microbial communities in southern Wisconsin, USA. Ag Sci. 2 (2), 131-137 (2011).
- Herzberger, A. J., Duncan, D. S., Jackson, R. D. Bouncing Back Plant-Associated Soil Microbes Respond Rapidly to Prairie Establishment. PloS One. 9 (12), 1-14 (2014).
- Fraterrigo, J. M., Balser, T. C., Turner, M. G. Microbial community variation and its relationship with nitrogen mineralization in historically altered forests. Ecology. 87 (3), 570-579 (2006).
- Kao-Kniffin, J., Balser, T. C. Elevated CO2 differentially alters belowground plant and soil microbial community structure in reed canary grass-invaded experimental wetlands. Soil Biol Biochem. 39 (2), 517-525 (2007).
- Mentzer, J. L., Goodman, R. M., Balser, T. C. Microbial response over time to hydrologic and fertilization treatments in a simulated wet prairie. Plant Soil. 284 (1-2), 85-100 (2006).
- Ushio, M., Wagai, R., Balser, T. C., Kitayama, K. Variations in the soil microbial community composition of a tropical montane forest ecosystem: Does tree species matter. Soil Biol Biochem. 40 (10), 2699-2702 (2008).
- Bartelt-Ryser, J., Joshi, J., Schmid, B., Brandl, H., Balser, T. Soil feedbacks of plant diversity on soil microbial communities and subsequent plant growth. Perspect Plant Ecol. 7 (1), 27-49 (2005).
- Fichtner, A., von Oheimb, G., Härdtle, W., Wilken, C., Gutknecht, J. L. M. Effects of anthropogenic disturbances on soil microbial communities in oak forests persist for more than 100 years. Soil Biol Biochem. 70, 79-87 (2014).
