

Summary
Immunolabeling для анализа различных популяций микротрубочек в развивающемся мозге данио рерио описаны методы, которые широко применимы для других тканей. Первый протокол излагаются оптимизированный метод для immunolabeling стабильное и динамичное микротрубочек. Второй протокол предоставляет метод для изображений и количественно зарождающейся микротрубочек специально.
Abstract
Микротрубочки (МТС) являются динамичной и хрупких структур, которые являются сложные изображения в естественных условиях, особенно в позвоночных эмбрионы. Immunolabeling методы описаны здесь, чтобы проанализировать различные популяции МТС в развивающихся нервной трубки zebrafish эмбриона. В то время как основное внимание уделяется нервной ткани, эта методология широко применяется для других тканей. Процедурах оптимизированы для рано для середины somitogenesis этап эмбрионов (1 Сомит до 12 сегменты), однако они могут быть адаптированы к целому ряду других этапов с относительно незначительными изменениями. Первый протокол предоставляет метод для оценки пространственного распределения стабильное и динамичное МТС и выполнить количественный анализ этих групп населения с программным обеспечением обработки изображений. Этот подход дополняет существующие средства изображения микротрубочек динамики и распределения в режиме реального времени, используя трансгенных линий или переходных выражение тегами конструкций. Действительно такие инструменты очень полезны, однако они не легко различать динамичного и стабильного МТС. Способность изображения и анализировать эти собственный микротрубочек населения имеет важные последствия для понимания механизмов поляризации клеток и морфогенеза. Второй протокол описывает метод для анализа нарождающихся МТС специально. Это достигается путем захвата de novo роста свойства МТС со временем, после деполимеризацию микротрубочек с наркотиками Нокодазол и восстановительный период после вымывания наркотиков. Этот метод еще не был применен к изучению МТС в zebrafish эмбриона, но является ценным assay для расследования в естественных условиях функции белков, замешанных в сборку микротрубочек.
Introduction
Микротрубочки (МТС) являются полимерами из α - и β-тубулина, собрать в линейной protofilaments, некоторые из которых в совокупности образуют полая трубка1,2. МТС, поляризационные структуры, с быстро растущей плюс концы и медленнорастущие минус концы, которые крепятся на Центросома или другие организации микротрубочек центр (КТВМ)3. De novo Формирование MT инициируется нуклеации на кольцо γ-тубулина комплекс (γ-Турецкая), которая предоставляет шаблон для Ассамблеи МТ4. В любой заданной ячейки две популяции МТС можно выделить то поворот над разными темпами. Динамический МТС исследовать их клеточной среды, переключение фаз роста и усадки в процессе, известном как Динамическая нестабильность5. В отличие от динамических МТС стабильной МТС межвегетационный и имеют более длительный период полувыведения, чем динамические МТС6.
Десятилетия исследований в клеточной биологии предоставила сложный спектр инструментов для изучения MT структуры и функции и привели к большой объем знаний об этих цитоскелетных элементов. Например, МТС играют центральную роль в создании и поддержании клеток полярности, который объясняется не только их внутреннюю полярность, но и для дифференциальной субцеллюлярные распределения стабильной по сравнению с динамической МТС7, 8. напротив, гораздо меньше понимается о MT архитектуры и функции в более сложной трехмерной (3-D) средах, таких как позвоночных эмбриона, отчасти из-за проблемы визуализации цитоскелета MT с высоким разрешением. Несмотря на это ограничение, Последнее поколение GFP-выражения трансгенных линий лейбл МТС или переходных выражение дневно тегами MT маркеров увеличилась нашего понимания динамических изменений, которые претерпевают МТС и их сотовых и роль в процессе развития в zebrafish эмбриона. Вся сеть MT может отражаться в трансгенных линий в котором тубулин – либо непосредственно помечены9 или тубулин полимеров косвенно помечены с помощью MT-связанных белков Даблкортин как киназы (Dclk) или Ensconsin (EMTB)10, 11. Другие линии (и конструкции) были созданы которые включить оценку MT внутреннюю полярность, специально маркировки МТ плюс концы или Центросома якорь минус заканчивается11,12,13, 14. сила этих инструментов заключается в способности учиться жить, MT динамика в развивающихся организмов. Такие исследования показали, например, пространственных и динамическое распределение МТС в конкретных клеточных популяций, ориентацию митотическая шпинделей в тканях происходят морфогенез (индикатор плоскости деление клеток), полярность MT полимера как она относится к процессам таких ячеек удлинение и миграции и темпы роста MT, определяется кометы скорость9,,1315. Ограничение этих инструментов заключается в том, что они не в легко различать стабильное и динамичное население MT.
Рисование от богатых ячейки биологии литературы, immunolabeling методы для изображения стабильное и динамичное МТС в zebrafish эмбриона описаны здесь, которые дополняют использования трансгенных линий. Широкое использование таких методов immunolabeling в zebrafish несколько сдерживается трудностями в сохранении целостности MT во время процедуры фиксации. Протокол 1 излагаются оптимизированный метод для immunolabeling всего, динамический, и стабильная МТС в сечения развивающихся данио рерио задний мозг. Кроме того простой метод, с помощью коммерчески доступное программное обеспечение описано для количественной оценки этих MT населения. Стабильные МТС отличаются от динамических МТС, основанные на несколько столб-поступательные изменения α-тубулина, например ацетилирования и detyrosination, которые накапливаются на стабильной МТС с течением времени16,17. В zebrafish эмбриона ацетилирования происходит на цилиарных и аксональной МТС, но не на стабильной межфазной МТС18, ограничивает полезность этого маркера к подмножеству стабилизированный МТС. В отличие от detyrosination, по-видимому, происходят на все стабильные МТС в zebrafish эмбриона18. Этот столб-поступательные изменения предоставляет карбоксильную терминал глутаминовой кислоты (detyrosinated тубулин) α-тубулина18 и могут быть обнаружены с помощью анти Glu тубулина19. Хотя detyrosination возникает преференциально на стабильной МТС, экспериментальные свидетельства указывает, что этот столб-поступательные изменения является результатом, а не причиной, MT стабильности16. Взаимные MT населения, состоящий из динамических МТС, отличается использованием антитела, анти-Tyr-тубулина, что конкретно признает tyrosinated форме α-тубулина19. После immunolabeling с этими маркеры и конфокальная томография количественный анализ МТС (длина, число и относительное изобилие) могут выполняться в определенных регионах развивающихся нервной трубки. Рациональный метод предоставляется здесь для проведения этого анализа с использованием трехмерной обработки изображений программное обеспечение. Этот метод может применяться для рассмотрения вопросов, касающихся морфогенеза и создание или созревания клеток полярности20. Действительно разработка поляризованные массивы стабильной МТС сопровождает многие развития событий, включая фоторецепторных морфогенеза21, эпителизация клеток в развивающихся нервной трубки18 и аксон формирования8.
2 протокол описывает в vivo адаптация assay биологии клетки для анализа МТС во время их Ассамблеи фазы (нуклеации/Анкоридж и роста)22,23. Нарождающейся МТС тому на Центросома и впоследствии привязан к subdistal придатков мать Центриоль23. Описан метод для анализа зарождающейся MT отрастание после деполимеризации. Этот протокол предоставляет сведения о Нокодазол лечения деполимеризуют МТС, процедура размыва наркотиков и период восстановления после лечения. MT re рост контролируется на регулярные промежутки временисмыв s пост immunolabeling с маркерами для всего МТС (анти-β-тубулина) наряду с маркерами для Центросома (анти-γ-тубулина) и ядра (4', 6-diamidino-2-phenylindole (DAPI)), в общем порядке, описанные в протокол 1. MT деполимеризации шаг настоящего Протокола имеет важное значение, поскольку позволяет оценки de novo MT роста, а не расширение существующих МТС. Поэтому этот метод отличается от других опубликованных процедур для измерения темпов роста MT (в отсутствии деполимеризации), используя маркер плюс чаевые как конец связывая протеин 3 сливается с зеленого флуоресцентного белка (EB3-GFP), как показано в Чан et al., 2012-11. Кроме того, этот assay особенно полезна для анализа эмбрионов дефектных в Ассамблее de novo MT, как сообщалось ранее мутантов NEDD1 , в которых вербовки γ-тубулина в Центросома нарушается, что приводит к неполной формирование нервной трубки и нейрональных дефекты24.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
этика заявление: процедуры описано ниже следуют Университет штата Мэриленд Балтимор округа животных ухода руководящих.
1. анализ стабильного и динамичного МТС с использованием Immunolabeling (протокол 1)
- ручной dechorionation эмбрионов до фиксации
- получить свежезаваренным породил эмбрионов, налив воды избыточные системы и затем собирая оставшиеся эмбрионы в пластиковой чашке Петри (см. Таблицу материалы).
- Удалить любой мусор из системы воды и переноса эмбрионов в новое блюдо заполнены с зародыш среднего (см. Таблицу материалов) для обеспечения в чистой среде эмбрионы.
- Позволяют эмбрионов для разработки на нужную сцену в контролируемой температурой инкубатор на 28,5 ° C.
- Место эмбрионов моложе, чем 24 часа после оплодотворения (hpf) в стеклянную посуду до dechorionation.
Примечание: Dechorionate эмбрионов до фиксации максимально быстрое проникновение фиксатором и сохранения целостности МТ. Использование эмбрионов среднего вместо воды системы для предоставления дополнительных Ca 2 + во время dechorionation. - Вручную удалить chorions из эмбрионов в чашке Петри, с помощью тонкой щипцы под микроскопом рассечения.
- Щепотка небольшой площади в круглые, прозрачные Хорио, которая окружает зародыш с парой щипцы и аккуратно вытащить щипцы врозь для создания разрыв мембраны.
- Увеличить открытия, деликатно посторонних на разорванные хориона, используя пинцет. Будьте осторожны, чтобы не коснуться эмбриона с щипцами, как он может разорваться.
- Фиксации поставил эмбрионов
- передачи поставил, dechorionated эмбрионов для 1.5 мл пробирок. Удалить столько среды эмбриона возможно с помощью стеклянной пипетки Пастера.
Примечание: Выполнение фиксации и наркотиков лечения на эмбрионах Янг (середина somitogenesis), до формирования нервных центров, посреднические болевых ощущений, которые требуют не дополнительные процедуры, чтобы облегчить боль во время эвтаназии. Этапы развития, как это определено в Kimmel и др., 1995 25. 4-5 и 11-12 этапов Сомит были использованы для получения изображения на рисунках 2 и 3. - Подготовить 4% параформальдегида (PFA) / тн Ассамблеи буфера (МАБ) фиксатором (см. Таблицу материалов), объединяя 1 мл 8% ПФА в 1 мл 2 X МАБ и добавив 2 мкл, 100% Тритон X-100 на 1 мл общего объема.
Предупреждение: Надевайте перчатки при работе с растворами, содержащими PFA и Тритон X-100, которые являются раздражителями кожи. - Исправление эмбрионов в 1 мл 4% PFA/МАБ фиксатором для 5 мин на 28,5 ° C. аспирата фиксатором с пипеткой, заменить его с 1 мл свежего фиксатором и Инкубируйте 3 ч при комнатной температуре (RT) на рокер.
Примечание: Образцы должны быть исправлены быстро их биологических температуре (28,5 ° C данио рерио) для предотвращения температур зависимая MT деполимеризации.
- передачи поставил, dechorionated эмбрионов для 1.5 мл пробирок. Удалить столько среды эмбриона возможно с помощью стеклянной пипетки Пастера.
- Аспирационная фиксатором и добавьте 1 mL 1 x трис амортизированное saline с NP40 (TBS-NP40) буфера. Аккуратно агитируйте на RT на рокер три раза за 5 мин. Хранение эмбрионов при температуре 4 ° C в 1 мл свежего 1 X TBS-NP40 не более 7 дней.
Предупреждение: Надевайте перчатки при работе с растворами, содержащими NP-40, раздражающие кожу. - Эмбрионов Sectioning для immunolabeling
- тепла RT 4% низкую температуру плавления (LMP) агарозы встраивание средне в закрытом контейнере, до тех пор, пока решение становится ясно, с помощью горячей плиты, равным 50 ° C, расположены недалеко от рассечения микроскопа . Держите контейнер закрыт между образцами и нагревают на протяжении всего процесса внедрения (шаги 1.4.4-1.4.6).
- Переноса эмбрионов от 1,5 мл центрифуга трубы в чашке Петри с помощью пипетки стеклянные и заполнить его с 1 X TBS-NP40.
- Удалить большой желток клеток эмбрионов somitogenesis этап (4-5 и 11-12 сегменты) в чашку Петри с помощью тонкой щипцы под увеличением рассечения Микроскоп 26. Держите эмбриона бутон хвост с одной парой щипцов и удаляйте клеток желток с другой парой для того чтобы сохранить ткани задний мозг. Передача де yolked эмбрионов в район Петри бесплатно мусора желток.
Примечание: Внедрить эмбрионов в агарозы заполненные формы отдельно для того, чтобы предотвратить преждевременное затвердение LMP агарозы. - Заполнения одной 12 мм x 5 мм x 3 мм и секущей плесень с 200 мкл расплавленный LMP агарозы с помощью микропипеткой. Выполните шаги 1.4.5.-1.4.6. быстро (в течение 20 s заполнения формы) внедрять эмбриона, прежде чем LMP агарозы остывает в RT и затвердевает.
- Использовать тонкой щипцы для передачи де yolked эмбриона, tailbud от Петри агарозы заполненные формы к его остроконечный конец под микроскопом рассечения.
- Использование тонкой щипцы сориентироваться эмбриона в форму таким образом, что vibratome сокращений в плоскости желаемого. Создайте разделы поперечной ориентируя эмбриона, таким образом, что задний мозг ткани проходит параллельно длине плесень с его дорсальной поверхности перед края и его передней поверхности конической региона завершения облицовки. Повторите шаги 1.4.4-1.4.6 для оставшихся эмбрионов.
- Разрешить вложение агарозы закрепить за 5 мин на RT.
- Генерировать 40 мкм секции высоких оси агарозы встроенных эмбрионов (шаги 1.4.1-1.4.7) с помощью vibratome с секущей блюдо, заполнены с 1 x TBS-NP40. Передавать интересующие разделы 24-ну пластины в 500 мкл 1 x TBS-NP40 с помощью тонкой щипцами. Место разделы только одного эмбриона на колодец.
Примечание: Обратитесь к ссылка 18 для получения более подробной информации. Убедитесь, что разделы остаются гидратированных на все времена в по крайней мере 250 мкл буфера и рок на низкой скорости (10-25 об/мин) для оставшихся шагов для предотвращения отделения от агарозы встраивание. Моющие средства представляют в блокировке и вымойте решения следует уменьшить поверхностное натяжение жидкой среды и погружения секций. Проверьте, что разделы остаются в скважинах, во время и после всех манипуляций. Соблюдайте осторожность для предотвращения случайно отбрасывая секции во время смывки.
- Удалить буфер и 500 мкл преграждая разрешение. Рок для по крайней мере 1 час на RT.
Примечание: Используйте блокировки решения, содержащего 5% сыворотки от принимающей видов каждого вторичного антитела, которые будут использоваться (см. Таблицу материалы). - Инкубировать в 300 мкл первичного антитела разводят в блокирующем буфере для 36-72 ч при температуре 4 ° C на рокер. Мыть дважды в 600 мкл 1 x TBS-NP40 на рокер для 30 минут каждый, на RT.
Примечание: Двойной метки разделов по инкубации в первичных антител против всего МТС (анти-β-тубулина, или анти-α-тубулина) и стабильной МТС (анти Glu тубулина) или динамические МТС (анти Tyr тубулин). Выберите первичного антитела, которые были подняты в различных принимающих видов при двойной маркировки для общей и post-translationally α-тубулина населения. Обратитесь к Таблице материалов для разбавления антитела. - Инкубировать в 300 мкл конъюгированных Флюорофор вторичных антител, разбавленных в блокирующем буфере на рокер для 16-24 ч, при температуре 4 ° C в темноте. Мыть дважды в 600 мкл 1 x TBS-NP40 на рокер для 30 минут каждый, на RT.
Примечание: Оберните несколькими хорошо блюдо, содержащие вторичные антитела в фольге с этого момента и после каждого манипуляции, чтобы предотвратить закалки. Выберите вторичные антитела, которые реагируют с принимающей иммуноглобулина первичного антитела. Выбор вторичного антитела флуорофоров, имеют отдельный, неперекрывающийся выбросов спектры. Обратитесь к Таблице материалов для разбавления антитела. - Инкубировать эмбрионов в 500 мкл раствора DAPI на рокер за 30 мин., на RT. мыть три раза в TBS-NP40 качания на RT на 5 мин.
Примечание: Ядерной маркировки обеспечивает сотовой связи для количественного определения МТ, выполненной на шаге 1.12. - Место падение среднего монтажа с анти затухания агентом по центру слайда, пыли. Используйте тонкой щипцы для передачи секции для монтажа средних капли. Место coverslip, пыли, поверх образца. Хранить в сухом, темном и прохладном месте, завернутый в фольгу, до тех пор, пока выполняется визуализация слайды.
Примечание: Кружили в разделах на задней части слайда с помощью острым концом перманентный маркер до изображений поможет определить секции при использовании микроскопом.
Конфокальный Imaging - горе разделы на Перевернутый лазерного сканирования Конфокальный микроскоп путем нанесения на слайд на сцену с coverslip, сталкивается с целью
- . Определить соответствующие оптика (цель, лазерной и настройки канала такие как получить и компенсировать) на слайде управления и держать их согласованность между образцы 27. Избегайте oversaturating пикселей для предотвращения потери данных.
- Захвата Z-стеки конфокальный изображения с использованием параметров канала для выбранной вторичное антитело флуорофоров и сохранить файлы изображений 27. Приобрести Z-стеки для каждого раздела.
Примечание: Реплицировать параметры, используемые для получения изображения на рисунках 2 и 3, используя следующие параметры приобретения: режим = XYZ; цель увеличения = 63 X объектив погружения нефти; Цель числовая апертура = 1.4; Z-step = 0.1 мкм; Z-глубина = 16.23 мкм. Используйте следующие настройки канала: DAPI возбуждения с 20% УФ диапазона лазер, диапазона фильтра выбросов = 430-480 Нм, фотоэлектронный умножитель (ПЛТ) прибыль = 525 V и смещение ПЛТ =-1.72%; 448 Нм Флюорофор (см. Таблицу материалов) возбуждения с 20% 488 нм лазер, диапазона фильтра выбросов = 493-573 Нм, получить ПЛТ = 689 V и смещение ПЛТ = – 0,2%; 594 Нм Флюорофор возбуждения с 32% 594 Нм лазер, диапазона фильтра выбросов = 608-706 Нм, получить ПЛТ = 768 V и смещение ПЛТ = -6,8%. - Сохранить файлы исходных данных с уникальными, описательные имена и создать копию для редактирования изображений программное обеспечение анализа.
- Сборник Z-стеки для отображения максимального прогнозы
- Открыть копию файла данных, с помощью общественного достояния 3-D изображение программного обеспечения для анализа (например, ImageJ). Убедитесь, что каждый канал отображается в виде последовательности отдельных изображений (Z-стеки).
- Разделение каналов изображения, используя следующую последовательность меню: “ изображения/цвет/разделить каналы ”.
- Создание объединенного изображения, наложение каналов интерес, используя следующую последовательность меню: " изображения/цвет/объединить каналы. " выберите 594 Нм, 488 нм и DAPI каналы, чтобы быть ложным окрашен в красный цвет, зеленый и синий, соответственно. Проверить " создать составной " и выберите " ОК " 28.
Примечание: Опустить канал DAPI передать более подробно конкретные МТС в максимальной проекции как показано на рисунке 2 и 3, по только выбор ложных цветов для других двух каналов. - Изучить Объединенные Z-стек и запишите начальную и конечную позиции внутренней лучший Z-самолетов для всех видимых каналов. Уволить внешней Z-самолеты, которые обычно имеют субоптимальные сигнала из-за неровной поверхности раздела. Обратитесь к ссылки 29 для деталей.
- Визуализировать Объединенные Z-стек как единый образ на 2-D, выполняя проекция максимальной интенсивности Z-стека, используя следующую последовательность меню анализ трехмерных изображений: " изображения/стек/Z-проект. " введите начальную и конечную позиции внутренний лучших Z-самолеты из шага 1.11.3 как " начала фрагмента " и " остановка срез, " соответственно. Выберите " Макс интенсивность " как тип проекции и нажмите " ОК ". Обратитесь к ссылки 28 для получения более подробной информации.
- Анализ MT маркировки
- открытое программное обеспечение анализа коммерческих трехмерного изображения. Выберите " создать библиотеку " и введите описательное имя для библиотеки изображений. Нажмите кнопку " создать. " перетащить файлы изображений в формате raw, созданные из конфокального микроскопа в библиотеку. Большие файлы требуют больше времени для передачи.
- Выбрать файл для анализа. Выбрать " расширенный фокус " от " вид " меню для отображения изображения объединены канала в главном окне.
- Настроить порог путем перетаскивания ползунка инструмент для каждого канала вправо или влево до тех пор, пока фонового сигнала сокращается и истинное сигнал является надежной. Отметить, что каждый канал показывает истинный сигнал для помечены молекулы (например, DAPI канал показывает продолговатые или митотическая ядер, но не auto флуоресценции в цитоплазме или агарозы).
- Выберите " Freehand региона интерес (ROI) " инструмент и наметить области интереса для анализа. Выберите " действия " вкладка последовали " культур к выбору " чтобы обрезать изображение. Сохраните файл под новым именем обрезанного изображения. Нажмите кнопку " измерения " вкладку, чтобы создать протокол для фильтрации определенных объектов, имеющих отношение к 3-D анализа.
- Сопротивления " найти объекты " в окне Протокол. Переименовать первый протокол " DAPI. " в в раскрывающемся меню выберите канал DAPI. Перетащите следующие параметры протокола DAPI и поместите их ниже " найти объекты " в следующем порядке (Таблица 1): " заполнить отверстия в ObjectsŔ " Отдельные прикосновение ObjectsŔ " Исключить объекты, SizeŔ " Исключить, не касаясь ROIs ".
Примечание: Цель параметров в таблице 1 должны сначала задать порог, который отбрасывает сигналов, распределение которых и размер несовместимы с размером анализируемого объектов. Например, устранить сигнал не достаточно большой, чтобы быть ядром при подсчете ядер. - Выполнение последовательности (шаг 1.12.9) собрать остальные фильтры для β-тубулина и другие маркеры, используя параметры в таблице 1.
- Выберите " меру " в нижней части каждого протокола. Выбрать " интенсивности и измерение объема " и " скелета длиной " для маркировки всех тубулин, но только бывший для сигнала DAPI.
- Нарисовать ROI вокруг области, чтобы быть измерены. Наблюдать за измерения под " Резюме " вкладке после того, как программное обеспечение обрабатывает региона. Скопируйте данные и сохранять их в реальные таблицы, как показано в таблице 2. Создайте резервную копию электронной таблицы для последующего анализа.
- Выберите измерения интереса (например, длина комплекта MT, количество MT связки/ядра, как показал с различные маркеры) в электронной таблице и проанализировать, чтобы определить средние значения для каждой группы,.
Примечание: Средняя длина комплекта MT = сумма ' означает скелетных длина для β-тубулина ' для каждого эмбриона, деленное на общее количество эмбрионов. Обратитесь к строке 20 Таблицы 2. Форматирование электронной таблицы таким образом, чтобы переменные и экспериментальных групп легко графике.
2. De Novo MT Ассамблеи Assay (протокол 2)
- конструкция и испытания нескольких хорошо потока через аппарат ( рис. 1) за 2 дня до эксперимента.
Примечание: Аппарат позволяет одновременное размыва нескольких экспериментальных групп, после Нокодазол лечения с помощью поставок из Таблицы материалов. Силиконовый герметик требует по крайней мере 24 часа сушки времени, прежде чем он представляет никакого риска токсичности для эмбрионов.- Разделить 50 мл пластиковых пробирок пополам вдоль, используя джиг или ленточнопильный.
- Cut 7 полукругов радиусом 3 см, из 70 мкм нейлон mesh и обрезать их приспосабливать плотно в одну половину Сплит центрифуга трубки. Клей полукругов в пластиковых пробирок параллельно маркировку градации 10 мл, используя герметик аквариум Сейф кремния. Разрешить устройству сухой в течение 2 дней и промойте путем замачивания в стакан воды для 2-3 ч.
- Линия (резьбовые) верхний конец резки пластиковых пробирок с пластилин, таким образом, что высота жидкости, сохранена в поток через устройство имеет глубину ¼ дюйма ( рис. 1, 2 и 3).
- Подготовить аппарат размыва, удалив поршень из 50 мл шприц и вставка 12 дюймов тонкой трубки в кончик. Нажмите трубы, насколько он будет идти и уплотнение вокруг сустава с помощью моделирования глины.
- Предварительно Намочите сетки с использованием эмбрионов среднего позволяют жидкости проходят через весь поток через устройство. Угол устройства на льду так что жидкость бассейнов во всех нишах, но по-прежнему впадает из передней, где находится глина ОПРАВЫ. С помощью кольца стенд, приостановить размыва аппарат выше потока через устройство на льду ( рис. 1).
- Chill 200 мл среды эмбриона на льду и налить достаточно в смыв аппарат для обеспечения что очищаются все пузырьки воздуха и скорость потока – приблизительно 7 мл/мин Отрегулируйте скорость потока, изменяя высоту шприца.
- Энзиматического dechorionate эмбрионов
- сделать рабочего раствора-неспецифический протеазы путем разбавления 10 мг/мл 1 мл неспецифической протеазы фондовая в 20 мл среды эмбриона.
- Химическая dechorionation выполнять на эмбрионах 1 ч до timepoint, когда они должны достичь желаемого стадии развития. Дайджест chorions, удалив зародыш среднего из 100 мм Петри содержащие устроили эмбрионов и добавления 20 мл раствора рабочей неспецифической протеазы.
- Инкубировать эмбрионов при 37 ° C за 5 мин
Примечание: Не превышает 5 мин или использовать более высокую концентрацию раствора неспецифической протеазы, как это приведет к эмбрионы падает врозь раз лечили Нокодазол. - Быстро Пипетка, неспецифичный протеазы и пополнить блюда с приблизительно 25 мл среды эмбриона. Повторить один раз.
- С помощью 1 мл стеклянной пипетки, переноса эмбрионов моложе 24 hpf стекло посуда защитить их от повреждения.
- Полная dechorionation удалив chorions с помощью пары тонкой щипцов, вручную, как описано в шаге 1.1.5.
- Место стекла Петри, содержащие dechorionated эмбрионов в инкубаторе 28,5 ° C не менее 30 мин, пока они не достигают желаемого стадии развития.
- Depolymerize существующих МТС
- подготовить рабочего раствора 5 мкг/мл Нокодазол, объединяя 50 мкл 1 мг/мл фондовых Нокодазол с 10 мл лед холодной зародыш среднего.
Предупреждение: Используйте перчатки при обращении с Нокодазол, раздражающие кожу. - Обмен эмбриона средней группе лечения Нокодазол с 10 мл холодной Нокодазол рабочего раствора. Место Петри на льду подходящее время для стадии развития (например, 1 h 4-5 Сомит эмбрионов). Отложите необработанных управления эмбрионов в чашке Петри на льду устанавливается вместе с размыва образцы в шаге 2.3.4.1.
- Переноса эмбрионов, с помощью пипетки полированного стекла 1 мл огонь для потока через аппарат, используя отдельные отсеки для каждой экспериментальной группы. Начало Нокодазол размыва средой поливая холодной эмбриона льда в верхней части 50 мл шприц.
Примечание: Используйте по крайней мере 30 эмбрионов в экспериментальной группе. Экспериментальных групп может состоять из управления эмбрионов или разнообразные Морфолино или РНК вводят эмбрионов. Смыв потребует в общей сложности около 150 мл среды эмбриона быть добавлены каждые 8-10 мин размыва Нокодазол продолжая ингибировать рост MT со льдом. Хранение эмбрионов на льду имеет важное значение для успеха этот assay, потому что МТС неустойчивы при низких температурах и холодной задержки развития в начале эмбрионов. - МТС позволяют вырастить после 20 мин размыва на RT путем переноса эмбрионов стекла Петри содержащие теплый среднего эмбриона (28,5 ° C) с помощью пипетки стеклянные полированной 1 мл огонь. Как только эмбрионы переносятся, запустите таймер.
- Исправить управления и размыва эмбрионов в 1 мин, 5 мин и 10 мин, закупорить приблизительно 10 эмбрионов в 1,5 мл центрифуга трубки, заполненные с 1 мл 4% PFA/МАБ исправить (28,5 ° C) и после направления в шаг 1.2.3.
- подготовить рабочего раствора 5 мкг/мл Нокодазол, объединяя 50 мкл 1 мг/мл фондовых Нокодазол с 10 мл лед холодной зародыш среднего.
- Подготовить образцы для immunolabeling, как описано в разделах 1.3-1.5.
- Immunolabel плавающей секции и изображения эмбрионов как описано в разделах 1.6-1.10 с следующие изменения спецификации основного антитела: использование 1: 500 кролик анти γ-тубулина и 1: 200 мыши анти β-тубулина.
- Процесс и анализ изображений с помощью программного обеспечения для анализа трехмерных изображений, как описано в шаге 1.12.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Анализ стабильное и динамичное МТС с использованием immunolabeling
В протокол 1распределение MT субпопуляциях во время ранних (нейронных Киль) и поздних стадиях развития нервной трубки (нейронных стержень) выявлено, используя Glu тубулина и Tyr тубулина как маркеры для стабильного и динамичного МТС, соответственно. Динамический МТС преобладают в задний мозг на стадии нейронных Киля (4-5 миобластах) (рис. 2A-D). Как Киль превращается в нейронных стержня (11-12 сегменты), стадии расширенной эпителизации, качественно меньше МТС иммунореактивных с Tyr-тубулина антитела (Рисунок 2E-H), особенно в брюшной части стержня. В отличие от Glu тубулина разрозненными и пунктата всей нейронных Киля (рис. 3А-D), но обогащается в брюшной нервной стержень вдоль MT участки (Рисунок 3E-H). Стрелки указывают на конкретные MT расслоений или структур где маркировки увеличивается.
Хотя антитела анти Glu тубулина и Tyr-тубулина были произведены в того же вида хост (недопущение двойной маркировки эксперимент), эти результаты показывают, что стабильный и динамичный MT маркеры редко пересекаются в zebrafish задний мозг. Во-первых брюшной нервной стержень имеет более стабильным (рис. 3F) чем динамический (Рисунок 2F) МТС. Эта тенденция обращена вспять в спинной нейронных стержень, в соответствии с моделью данио рерио Нейруляция, в котором спинной ткани остается динамических пока нервной трубки будет сформирован20. Во-вторых хотя митотического веретена полностью помечены с антителом Tyr тубулина в нейронных Киля (Рисунок 2D, наконечники стрел), только база шпинделя, совпадающих с Центросома, помечается маркером стабильности (Glu тубулина Рисунок 3 D, стрелок). Β-тубулина иммунофлюоресценции, общие для обоих анализов, сообщает экспериментатор распределения всех МТС и обеспечивает основу для уволить неспецифичный Этикетировочный.
Измерение объектов с помощью программного обеспечения для анализа 3-D изображения приводит к большой объем данных, которые могут быть организованы в удобную таблицу (Таблица 2). Чтобы сделать длина, количество и площадь измерений, мы используем только подмножество данных, доступных для анализа. Один из компонентов данных, мы не дальнейшего анализа является количество объектов определены. Это число используется в качестве внутреннего контроля качества, как число должно не различаются между как секции и соотношение ядер для МТС должна оставаться же в состоянии единственного метода лечения. Останец является показателем, что либо анализ необходимо перезапустить с скорректированным фильтры или что изображение слишком плохо помечены для анализа. Таким образом все останец изображения следует недоцератопс с скорректированных настроек. В разделе останец должны рассматриваться признаков плохого маркировки или физический ущерб, который может привести к пунктам необычный объект. После завершения анализа и контроль качества, полезная информация может быть восстановлена из необработанных данных таких, как средняя продолжительность общий МТС и стабильная МТС или соотношение стабильной МТС всего МТС (Таблица 3). Помимо этих измерений многие другие метрики можно получить с помощью программного обеспечения анализ трехмерных изображений, который может быть использован для сделать выводы о МТС или их отношения к другим клеточных структур (ядро, Центросома и т.д.).
De novo MT Ассамблеи пробирного
Лечение Нокодазол depolymerizes МТС, что приводит к диффузным маркировки (рис. 4A, 4 D и 4 G). Как вырастить МТС, они простираются от Центросома (Рисунок 4B, 4E и 4 H), однако, это не может быть очевидным в одной плоскости из-за их неплоских траектории (рис. 4C, 4F и 4I). Тем не менее некоторое программное обеспечение анализа изображения способны измерения длины в 3-D, позволяя оценки роста MT после Нокодазол подложка (Таблица 4). Важные наблюдения, которые могут быть получены из набора данных в таблице 4 является, что средняя длина МТС, по-видимому, возрастать с течением времени после вымывания Нокодазол во всех регионах нервной трубки проанализированы. Как упоминалось выше, другие виды метрик, полученные от анализа программного обеспечения 3-D изображение может обеспечить сотовой связи для интерпретации данных МТ (к примеру, коэффициент МТС на ядро).
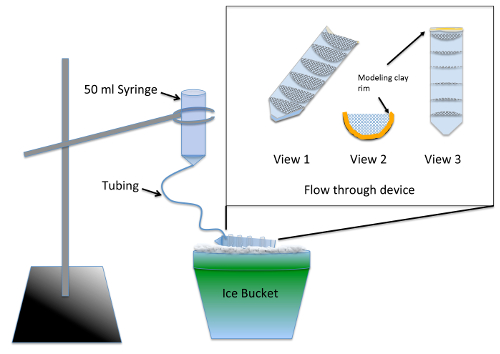
Рисунок 1 : Иллюстрация размыва аппарат для de novo MT Ассамблеи пробирного. Врезные является крупный поток через устройства из сетки склеиваются в 50-мл пластиковых пробирок, разрезать вдоль пополам. Сетка compartmentalizes потока через устройство таким образом, что несколько экспериментальных групп могут быть обработаны одновременно. Во время использования зародыш среднего добавляется в шприц и медленно протекает через трубки для заполнения потока через устройство, обеспечивая постоянное полоскание для всех экспериментальных групп. Пожалуйста, нажмите здесь, чтобы посмотреть большую версию этой фигуры.

Рисунок 2: Использование immunolabeling для изображения динамических МТС. Dechorionated эмбрионы были зафиксированы на соответствующих этапах (4-5 A-D и 12-13 сегменты в E-H), поперечно секционного через задний мозг и полученных с антителами против β-тубулина (зеленый в A и E) чтобы пометить все МТС и tyrosinated α-тубулина (красный в B и F) выявить динамический MT населения. Высокодинамичный МТС можно увидеть в Объединенном изображения (C, G) и их более высоких увеличениях (D, H) как районы, где желтая метка является видимым (наконечников в D, H). Масштаб баров = 25 мкм (A-C и E-G) и 10 мкм (D и H). Пожалуйста, нажмите здесь, чтобы посмотреть большую версию этой фигуры.

Figure 3: использование immunolabeling для изображения стабильной МТС Dechorionated эмбрионы были исправлены, секционного через задний мозг и полученных на соответствующих этапах (4-5 миобластах в A -D и 12-13 сегменты в E-H). Стабильные МТС помечены антител против detyrosinated форме α-тубулина (Glu тубулина) (красный в B и F) в то время как общая МТС были визуализированы с общей β-тубулина антитела (зеленый в A и E). Красные и желтые сигналы в Объединенном изображения (C, G) и их более высоких увеличениях (D, H) представляют области высокой стабильности MT (наконечников в D, H). Масштаб баров = 25 мкм (A-C и E-G) и 10 мкм (D и H). Пожалуйста, нажмите здесь, чтобы посмотреть большую версию этой фигуры.

Рисунок 4: Использование immunolabeling для изображения зарождающейся МТС. Dechorionated эмбрионы были зафиксировано на 4-5 миобластах и поперечно секционного через задний мозг. Разделы были полученных с β-тубулина (D, E и F) в ознаменование растущих МТС и γ-тубулина (A, Bи C) для обозначения точки/Центросома зародышеобразования. Спинной части нервной трубки упаковывается в (A, D; B, E и C-F) и на увеличение (G, H, я, соответственно) раскрыть ядер (DAPI, голубой), centrioles (γ-тубулина, красный) и общей МТС (β-тубулина, зеленый). Белые стрелки: colocalization МТС и centrioles; Желтые стрелки: второй Центриоль ячейка видима. Масштаб баров = 25 мкм (A-F) и 10 мкм (G-I). Пожалуйста, нажмите здесь, чтобы посмотреть большую версию этой фигуры.

Таблица 1: Параметры по умолчанию для фильтрации объектов в 3-D изображение анализа программного обеспечения.

Таблица 2: представитель сырой набор данных, полученных с помощью анализа 3-D изображений программное обеспечение для анализа стабильные МТС. Каждый столбец представляет измерения из одного раздела. Мин: маленький измерения; Макс: крупнейших измерения; SD: стандартное отклонение; SE: Стандартная ошибка.

Таблица 3: Примеры наборов данных, которые могут быть получены от 3-D изображений программное обеспечение для анализа для количественного определения стабильных МТС. Выберите измерения средней длины общей (β-тубулина) и стабильная МТС (Glu тубулина) рассчитывается путем принятия в среднем средняя скелетных длины для соответствующей метки от всех образцов (см. Таблицу2) и соотношение стабильной для всего (МТС Glu-tubulin полосы на β-тубулина полосы) рассчитывается путем принятия число средняя β-тубулина, деленное на среднее количество Glu тубулина.

Таблица 4: Примеры наборов данных, которые могут быть получены из трехмерного изображения анализ программного обеспечения для анализа de novo MT Ассамблеи. Представитель результаты от de novo MT Ассамблеи эксперимент, сравнивая наборы данных, полученные для трех точек восстановления время (1, 5 и 10 мин) после Нокодазол смыв. Для каждой точки время измерения, полученные для ядерной граф, centrioles (γ-тубулина puncta), количество всего МТС (β-тубулина полосы), показаны для отдельных регионов образа проанализированы (поперечное сечение развивающихся нервной трубки).
Subscription Required. Please recommend JoVE to your librarian.
Discussion
В настоящее время существует множество методов для визуализации динамики MT в начале развития данио рерио, начиная от живых изображений тегами молекул immunolabeling из фиксированной ткани11,12,,1314. Хотя МТС в одной ячейке может существовать в динамический или стабильного государства, эпителизация — это процесс, в котором МТС, со временем постепенно стабилизируются. С помощью маркеров для стабильного и динамичного МТС предлагает способ для визуализации этого явления. Представленные здесь метод использует возможности 3-D изображений программное обеспечение для количественного определения переход от динамических стабильный MT населению в сечении данио рерио эмбриональных тканей. В протоколе 2метод используется для обозначения различных населения зарождающейся МТС и следовать их зарождения и роста сверхурочных.
МТС сложно изображения в их родном государстве из-за их склонности к деполимеризуют. Таким образом ключевым компонентом этого метода является быстрой фиксации МТС на протяжении всего эмбриона. Это достигается путем запуска фиксации физиологических температурах и с использованием буфера, которая стабилизирует МТС и увеличивает проницаемость эмбриона. Время фиксации также важна, как сократить фиксации не арест МТС при чрезмерной фиксации может маскировать epitopes, мешая антитела связывая. Предлагаемые фиксации времени 3-4 h работает с эмбрионов, которые в середине гаструляция до 24 ч после оплодотворения. Эмбрионы в маленьких конце шкалы времени должна быть исправлена для ближе к 3 ч в то время как старые эмбрионов может понадобиться всего 4 h. Даже с надлежащей фиксации МТС будет деполимеризуют со временем, поэтому секционирование и immunolabeling должна происходить в течение одной недели фиксации.
Как только ткань должным образом исправлена, могут возникнуть проблемы с immunolabeling. Наиболее распространенные проблемы было плохое проникновение через центр ткани, особенно, если слишком много разделов инкубируют в том же хорошо. Повышение концентрации первичного антитела и инкубации время для первичных и вторичных антител, в сочетании с увеличением моющих средств для улучшения permeabilization эмбрионы будут улучшить большинство проблем immunolabeling. Если антитела обозначая сбой из-за фиксации или immunolabeling проблем, можно определить причину, просмотрев антитела обозначая шаблон. Плохой фиксации приведет к интенсивным маркировки в мембране и диффузный маркировки в цитоплазме, чрезмерной фиксации приведет к слабой маркировки, которая сохраняет MT архитектуры. Плохое проникновение антитела, однако, будет отображаться как районы в центре ткани без маркировки.
Способность анализировать MT изображения в значительной степени зависит от высокого качества изображений. Для захвата MT длина в 3-D, должны использоваться минимальный размер Z-шаг, цель и числовая апертура. Изображения, показанные здесь были захвачены с 63 X Нефть встраивание задачи с 1.4 числовая апертура, производить следующее: пиксель = 240 Нм, Z-шаг = 0,1 мкм, размер стека Z = 16.252 мкм. Потому что ширина одного MT-25 Нм, примерно в 10 раз ниже предела резолюции для световой микроскоп, это метрических не может быть точно измерить с помощью этой техники. Вместо этого может быть измерена только MT длины равным или больше чем размер минимальной пикселя, достижимой во всех трех измерениях. Линии и/или усреднение кадра может расширить определение сигнала MT. MT анализа должны быть зарезервированы для секции высокого качества. В то время как ткани с плохой фиксации не может отражаться и проанализированы, мягкая overfixation может уравновешиваться тщательно увеличение интенсивности лазера и получить обнаружить слабый сигнал при сохранении хорошей динамический диапазон. Бедных антитела проникновения, хотя не оптимальный, могут быть исправлены путем ограничения приобретения изображения хорошо обозначенные районы, что приводит к визуализации тоньше секции (5-10 мкм). Высокий фон от маркировки может быть компенсировано для настройки фильтра. Однако, если любой из этих корректировок сделали, необходимо проверить, что фильтры порог приемлемо на каждой плоскости Z-стека.
Программное обеспечение для анализа 3-D изображения позволяет экспериментатору определить MT длина, площадь, угол, изобилия и другие метрики в трехмерном пространстве разделов фиксированной ткани. Метод, описанный здесь предоставляет направления для получения таких данных, с использованием имеющегося программного обеспечения. Однако фильтрующие модули могут быть адаптированы к общественным достоянием программного обеспечения с соответствующим плагины или макросы, что делает анализ доступной для всех. До анализа raw изображений должны быть показатели, чтобы избежать включая фон и неконкретные сигналы в квантификации. После завершения анализа и данные передаются в работоспособной электронную таблицу, многие выводы могут быть сделаны из набора данных. Одним из расчеты, произведенные здесь был глу тубулина полосы полосы β-тубулина, или соотношение стабильной МТС всего МТС, где 1 показывает, что весь цитоскелета MT стабилизировалось в ROI. Если экспериментатор желает дополнить их количественных данных, генерации формат файла полированной тегами изображения (TIFF) изображение с масштаба бара является легким с 3-D изображение анализа программного обеспечения.
Этот assay позволяет функциональный анализ белков, замешанных в MT Ассамблее, в естественных условиях. Если immunolabeling осуществляется на чередующихся последовательных секций, этот протокол может использоваться для изучения динамичный и стабильный МТС в том же эмбриона. В будущем модификации, такие как увеличение моющие средства или измененные вложения углы позволит использование этих методов для пожилых эмбрионов и более широкий спектр анатомические вопросов.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Авторы не имеют ничего сообщать.
Acknowledgments
Конфокальный микроскоп был приобретен с средств от США Фонд национальной науки (NSF) Грант #DBI-0722569. Поддержка исследования осуществлялась в США национальные институты здравоохранения/Национальный институт Генеральной медицинских наук (NIH/NIGMS) Грант #GM085290 и Департамент обороны США (DOD) Грант #W81XWH-16-1-0466 присуждена Brewster Р.М. E. Vital была поддержана грантом UMBC Говард Хьюз медицинский институт через довузовского и Бакалавриат Наука образовательная программа, предоставить #52008090. С.п. Браун получил поддержку от США Департамента образования GAANN стипендий, стипендиатом Мейерхофф финансируется гранта NIH/NIGMS, #GM055036 и ассистентура исследований, финансируемых в США DOD Грант #W81XWH-16-1-0466.
Materials
| Name | Company | Catalog Number | Comments |
| Agarose | Used to treat petridishes. Prepare 1% agarose by heating a solution of 1 gram agarose per 100 ml 1X embryo medium in a microwave until polymerized. |
||
| Kpipes | Sigma | P7643 | |
| NaCl | Sigma | S7653 | |
| Tris-HCl | Sigma | T3253-500G | |
| KCl | Sigma | P9333-500G | |
| CaCl2·2H2O | Sigma | C5080 | |
| NP-40 | American Bioanalyticals | AB01424 | |
| EGTA | Sigma | E3889-25G | |
| MgCl2 | Sigma | M2670-500G | |
| Bovine serum albumin (BSA) | Fisher | BP1605 | |
| Triton-x | American Bioanalyticals | AB02025 | |
| Anti-Fade mounting medium | Invitrogen | P10144 | |
| Mouse anti-β-tubulin | Developmental studies Hybridoma Bank | E7 | 1/200 |
| Rabbit anti-γ-tubulin | Genetex | GTX113286 | 1/500 |
| Rabbit anti-α-tubulin | Genetex | GTX108784 | 1/1000* |
| Rabbit anti-detyrosinated-tubulin | Millipore | AB3201 | 1/200-1/1000* Titrate antibody with first use of new lot. |
| Rabbit anti-tyrosinated-tubulin | Millipore | ABT171 | 1/500 |
| Mouse anti-centrin | Millipore | 04-1624 | 1/1000 |
| Goat 488 anti-rabbit | Thermofisher | A11008 | 1/500 |
| Goat 594 anti-rabbit | Thermofisher | A11012 | 1/500 |
| Goat 594 anti-mouse | Thermofisher | A11005 | 1/500 |
| Goat 488 anti-mouse | Thermofisher | A11001 | 1/500 |
| Vibratome | Vibratome | 1500 | |
| Forceps | World Precision Instruments | 555227F | |
| 100 mm petri dish | Cell treat | 229693 | |
| 35 mm petri dish | Cell treat | 229638 | |
| 50 ml falcon tube | Fisher | 14-432-22 | |
| Woven nylon mesh 70 um | Amazon.com | B0043D1SZG | |
| Micropipette | Gilson | F123602 | |
| Glass pipette | Fisher | NC-999363-9 | |
| Aquarium sealant | Amazon.com, by MarineLand | Silicone Sealer 1 oz (Tube) | |
| Ring stand | Fisher | 14-675BO | |
| Microbore PTFE Tubing, 0.022"ID | Cole-Parmer | WU-06417-21 | |
| Modeling clay | Amazon.com | Sargent Art 22-4000 | Any wax or oil based non-toxic modeling clay will suffice |
| Clamp | Fisher | 02-215-466 | |
| 60ml syringe | Fisher | 14-820-11 | |
| Embryo medium (E3) | 34.8 g NaCl 1.6 g KCl 5.8 g CaCl2·2H2O 9.78 g MgCl2·6H2O To prepare a 60X stock, dissolve the ingredients in H2O, to a final volume of 2 L. Adjust the pH to 7.2 with NaOH. Autoclave. To prepare 1X medium, dilute 16.5 mL of the 60X stock to 1 L. |
||
| Blocking Solution | 50 ml TBS-NP-40 2.5 ml normal goat serum 1 g BSA 625 µl Triton-X |
||
| TBS-NP-40 (pH 7.6) | 155 mM NaCl 10 mM Tris HCl 0.1% NP-40 |
||
| 2x MAB (pH6.4) | 160 mM KPIPES 10 mM EGTA 2 mM MgCl2 |
||
| Commercial 3-D Image processing Software | PerkinElmer | Volocity (V 6.2) | |
| Dry block heater | VWR | 12621-108 | Used as a hot plate to melt agarose in Protocol 1. |
| Dissecting Microscope | Leica | MZ12 | |
| Confocal Microscope | Leica | SP5 | |
| Flat embedding mold | emsdiasum.com | BEEM 70904-01 | |
| Public domain image processing software | NIH | ImageJ (V 1.5) | |
| * Success varies by lot number | |||
References
- Akhmanova, A., Steinmetz, M. O. Tracking the ends: a dynamic protein network controls the fate of microtubule tips. Nat Rev Mol Cell Biol. 9 (4), 309-322 (2008).
- Conde, C., Cáceres, A. Microtubule assembly, organization and dynamics in axons and dendrites. Nat Rev Neurosci. 10 (5), 319-332 (2009).
- Kaverina, I., Straube, A. Regulation of cell migration by dynamic microtubules. Semin Cell Dev Biol. 22 (9), 968-974 (2011).
- Kollman, J. M., Merdes, A., Mourey, L., Agard, D. A.
- Howard, J., Hyman, A. A. Growth, fluctuation and switching at microtubule plus ends. Nat Rev Mol Cell Biol. 10 (8), 569-574 (2009).
- Schulze, E., Kirschner, M. Dynamic and stable populations of microtubules in cells. J Cell Biol. 104 (2), 277-288 (1987).
- Gundersen, G. G., Kalnoski, M. H., Bulinski, J. C. Distinct populations of microtubules: Tyrosinated and nontyrosinated alpha tubulin are distributed differently in vivo. Cell. 38 (3), 779-789 (1984).
- Li, R., Gundersen, G. G. Beyond polymer polarity: how the cytoskeleton builds a polarized cell. Nat Rev Mol Cell Biol. 9 (11), 860-873 (2008).
- Asakawa, K., Kawakami, K. A transgenic zebrafish for monitoring in vivo microtubule structures. Dev Dyn Off Publ Am Assoc Anat. 239 (10), 2695-2699 (2010).
- Wühr, M., Tan, E. S., Parker, S. K., Detrich, H. W., Mitchison, T. J. A model for cleavage plane determination in early amphibian and fish embryos. Curr Biol CB. 20 (22), 2040-2045 (2010).
- Tran, L. D., Hino, H., et al. Dynamic microtubules at the vegetal cortex predict the embryonic axis in zebrafish. Development. 139 (19), 3644-3652 (2012).
- Butler, R., Wood, J. D., Landers, J. A., Cunliffe, V. T. Genetic and chemical modulation of spastin-dependent axon outgrowth in zebrafish embryos indicates a role for impaired microtubule dynamics in hereditary spastic paraplegia. Dis Model Mech. 3 (11-12), 743-751 (2010).
- Yoo, S. K., Lam, P. -Y., Eichelberg, M. R., Zasadil, L., Bement, W. M., Huttenlocher, A. The role of microtubules in neutrophil polarity and migration in live zebrafish. J Cell Sci. 125 (23), 5702-5710 (2012).
- Andersen, E. F., Halloran, M. C. Centrosome movements in vivo correlate with specific neurite formation downstream of LIM homeodomain transcription factor activity. Development. 139 (19), 3590-3599 (2012).
- Lee, S. -J. Dynamic regulation of the microtubule and actin cytoskeleton in zebrafish epiboly. Biochem Biophys Res Commun. 452 (1), 1-7 (2014).
- Bulinski, J. C., Gundersen, G. G. Stabilization and post-translational modification of microtubules during cellular morphogenesis. BioEssays. 13 (6), 285-293 (1991).
- Magiera, M. M., Janke, C. Chapter 16 - Investigating Tubulin Posttranslational Modifications with Specific Antibodies. Methods Cell Biol. 115, 247-267 (2013).
- Hong, E., Jayachandran, P., Brewster, R. The polarity protein Pard3 is required for centrosome positioning during neurulation. Dev Biol. 341 (2), 335-345 (2010).
- Westermann, S., Weber, K. Post-translational modifications regulate microtubule function. Nat Rev Mol Cell Biol. 4 (12), 938-948 (2003).
- Jayachandran, P., Olmo, V. N., et al. Microtubule-associated protein 1b is required for shaping the neural tube. Neural Develop. 11, 1 (2016).
- Nam, S. -C. Role of Tau, a microtubule associated protein, in Drosophila photoreceptor morphogenesis. Genes N Y N 2000. 54 (11), 553-561 (2016).
- Abal, M., Piel, M., Bouckson-Castaing, V., Mogensen, M., Sibarita, J. -B., Bornens, M. Microtubule release from the centrosome in migrating cells. J Cell Biol. 159 (5), 731-737 (2002).
- Delgehyr, N., Sillibourne, J., Bornens, M. Microtubule nucleation and anchoring at the centrosome are independent processes linked by ninein function. J Cell Sci. 118 (8), 1565-1575 (2005).
- Manning, J. A., Lewis, M., Koblar, S. A., Kumar, S. An essential function for the centrosomal protein NEDD1 in zebrafish development. Cell Death Differ. 17 (8), 1302-1314 (2010).
- Kimmel, C. B., Ballard, W. W., Kimmel, S. R., Ullmann, B., Schilling, T. F.
- Beck, A. P., Watt, R. M., Bonner, J. Dissection and Lateral Mounting of Zebrafish Embryos: Analysis of Spinal Cord Development. JoVE J Vis Exp. (84), e50703 (2014).
- FÖldes-Papp, Z., Demel, U., Tilz, G. P. Laser scanning confocal fluorescence microscopy: an overview. Int Immunopharmacol. 3 (13-14), 1715-1729 (2003).
- Ferreira, T., Rasband, W. S. ImageJ User Guide - IJ 1.46. , Available from: https://imagej.nih.gov/ij/docs/guide/ (2010).
- Z-functions - ImageJ. , Available from: https://imagej.net/Z-functions (2017).
