

Summary
Immunolabeling 的方法来分析不同群体的微管在发展斑马鱼大脑中描述, 这是广泛适用于其他组织。第一个协议概述了 immunolabeling 稳定和动态微管的优化方法。第二个协议提供了一种具体的图像和量化初生微管的方法。
Abstract
微管 (MTs) 是动态的和脆弱的结构, 是挑战的图像在体内, 特别是在脊椎动物的胚胎。本文介绍了 Immunolabeling 方法, 分析了斑马鱼胚胎发育神经管中 MTs 的不同种群。虽然重点是神经组织, 这种方法是广泛适用于其他组织。程序为早期 mid-somitogenesis 阶段的胚胎 (1 节到12节) 进行了优化, 但是它们可以适应其他阶段的一系列相对较小的调整。第一个协议提供了一种评估稳定和动态 MTs 的空间分布的方法, 并用图像处理软件对这些种群进行定量分析。这种方法补充了现有的工具, 图像微管动力学和分布在 real-time, 使用转基因线或瞬态表达的标记结构。事实上, 这些工具非常有用, 但是它们并不容易区分动态和稳定的 MTs。图像和分析这些不同的微管种群的能力对于理解细胞极化和形态发生的机制有着重要的意义。第二个协议概述了专门分析初生 MTs 的技术。这是通过在一段时间内捕获 MTs 的de 从头生长特性来完成的, 这是随着药物 nocodazole 的微管降解和药物冲洗后的恢复期。这项技术尚未被应用于斑马鱼胚胎的 MTs 研究, 但它是一个有价值的方法, 以调查的体内功能的蛋白质牵连的微管组装。
Introduction
微管 (MTs) 是α和β-蛋白的聚合物, 组合成线性 protofilaments, 其中的几个组合成中空的管子1,2。MTs 是极化结构, 具有快速增长的加端和缓慢增长的负端, 锚定在中心或其他微管组织中心 (MTOC)3。De 从头mt 形成是由核在γ-蛋白环复合体 (γ-土耳其), 它提供一个模板为 mt 汇编4。在任何给定的单元格中, 两个 MTs 的种群可以区别开来, 以不同的速率翻转。动态 MTs 通过在称为动态不稳定的过程中切换增长阶段和收缩期来探索它们的蜂窝环境5。与动态 mts 不同, 稳定 mts 是 non-growing 的, 其半衰期比动态 mts6长。
几十年的细胞生物学研究提供了一系列复杂的工具来研究 MT 的结构和功能, 并在这些骨架元素上形成了大量的知识。例如, mts 在建立和维护细胞极性方面起着核心作用, 这不仅归因于它们的固有极性, 而且也归功于稳定与动态 mts 的微分亚空间分布7, 8. 相比之下, 在更复杂的三维 (3 维) 环境 (如脊椎动物胚胎) 中, 对 mt 体系结构和功能的了解要少得多, 部分原因是高分辨率下的 mt 细胞骨架成像的挑战。尽管这一限制, 最近一代的 GFP 表达的转基因线, 标签 mts 或瞬态表达的荧光标记 MT 标记已增加我们的理解的动态变化, mts 接受和他们的细胞和斑马鱼胚胎发育的作用。整个 MT 网络可以成像的转基因线, 其中蛋白是直接标记为9或蛋白聚合物是间接标记使用 MT 相关的蛋白 Doublecortin 样激酶 (Dclk) 或 Ensconsin (EMTB)10, 11。其他的线 (和构造) 已经产生, 通过专门标记 mt 的正负端或中心锚定的减号11,12,13,来评估 mt 的内在极性14. 这些工具的威力在于能够研究在活体和发育中的生物体的 MT 动力学。这些研究表明, 例如, 在特定细胞群体中 MTs 的空间和动态分布, 发生形态发生的组织中有丝分裂纺锤体的方向 (细胞分裂平面的指示器), MT 聚合物的极性因为它涉及到的过程, 如细胞伸长和迁移, 和 MT 生长率确定的彗星速度9,13,15。这些工具的局限性在于它们不容易区分稳定的和动态的 MT 种群。
本文从丰富的细胞生物学文献中, 对斑马鱼胚胎的图像稳定和动态 MTs 的 immunolabeling 方法进行了描述, 这是对转基因品系的补充。这种 immunolabeling 方法在斑马鱼中的广泛应用, 由于在固定过程中保持 MT 完整性的困难而受到了一定的阻碍。协议 1概述了在开发的斑马鱼后脑的横断面上 immunolabeling 总、动态和稳定 MTs 的优化方法。此外, 还描述了使用商用软件的直接方法来量化这些 MT 的数量。稳定 mts 的区别, 从动态 mts 的几个修饰修改的α-蛋白, 如乙酰化和 detyrosination, 在稳定的 mts 积累了一段时间16,17。在斑马鱼胚胎中, 乙酰化发生在睫状和轴突 mts 上, 但不在稳定的相间 mts18上, 将此标记的用处限制在稳定 mts 的子集上。相反, detyrosination 似乎发生在所有稳定的 MTs 在斑马鱼胚胎18。这种修饰修饰暴露了α-蛋白 (detyrosinated 蛋白)18的羧基末端谷氨酸, 并且可以使用反谷氨酸-蛋白19检测。虽然 detyrosination 优先于稳定 MTs, 但实验证据表明, 此修饰修改是由于 MT 稳定性16造成的, 而不是原因。相互 MT 人口, 由动态 MTs 组成, 是区分使用抗体, 反 Tyr-蛋白, 特别承认 tyrosinated 形式的α蛋白19。随着这些标记和共聚焦成像的 immunolabeling, 对 MTs (长度、数量和相对丰度) 的定量分析可以在发展中神经管的定义区域进行。这里提供了一种简化的方法, 用于使用3维图像处理软件执行此分析。此方法可用于解决有关形态发生和细胞极性的建立或成熟的问题20。的确, 稳定 MTs 的极化阵列的细化伴随着许多发展事件, 包括感光细胞发生形态21, 上皮在发育中的神经管的18和轴突形成8。
协议 2描述了一个用于在其组装阶段 (核化/锚定和生长)2223中分析 MTs 的细胞生物学检测的体内。初生的 MTs 在中心有核, 并随后锚定到母亲中心的 subdistal 附属物23。介绍了一种分析降解后初生 MT 再生的方法。本议定书详细介绍了解 MTs 的 nocodazole 治疗、药物冲洗程序和后处理恢复期。再生定期监测根据协议 1中所述的一般过程, immunolabeling 的后冲蚀与中心 (反γ-蛋白) 和原子核 (4 "、6-diamidino-2-吲 (DAPI)) 的标记一起与总 MTs (抗β-蛋白) 的标记。该协议的 mt 降解步骤是必不可少的, 因为它能够评估de 从头mt 的增长, 而不是预先存在的 MTs 的扩展。因此, 这一技术有别于其他公布的程序, 以测量 MT 生长速率 (在没有降解) 通过使用一个加号标记, 如最终结合蛋白3融合到绿色荧光蛋白 (EB3-GFP), 如在et al.中所示,201211。此外, 这种方法对于分析de 从头MT 组件中的缺陷尤其有用, 例如以前报告的NEDD1突变体, 在该变种中, 对中心的γ蛋白的征募被削弱, 导致不完整神经管形成和神经元缺陷24。
Subscription Required. Please recommend JoVE to your librarian.
Protocol
道德声明: 下面描述的程序遵循马里兰大学巴尔的摩县动物保育指南.
1. 使用 Immunolabeling 的稳定和动态 MTs 的分析 (协议 1)
- 手动 dechorionation 前固定的胚胎 (
- 通过浇注多余的系统水来获得新鲜的胚胎)然后将剩余的胚胎收集到塑料培养皿中 (参见 材料表 ).
- 清除系统水中的任何碎片, 将胚胎移植到一个充满了胚胎培养基的新皿中 (参阅 材料表 ), 以确保胚胎在清洁的环境中发育.
- 允许胚胎在28.5 和 #176 温度控制的恒温箱中发育到所需的阶段; C.
- 在 dechorionation 之前, 将小于24小时胚乳 (hpf) 的胚胎放置在玻璃盘中.
注: Dechorionate 前的胚胎固定, 以最大限度地快速渗透的固色剂和保存 MT 完整性。使用胚胎培养基代替系统水, 在 dechorionation 期间提供所需的额外 Ca 2 + . - 在培养皿中, 用细钳在解剖显微镜下手动取出胚胎中的 chorions。
- 在圆形的透明绒毛膜中捏出一个小区域, 用一对镊子将胚胎围住, 并将镊子轻轻地拉开, 在膜上形成破裂.
- 用镊子巧妙地窥探破裂的绒毛膜, 扩大开口。注意不要用钳子碰触胚胎, 因为它可能破裂.
- 分阶段胚胎的固定, dechorionated 胚胎移植到1.5 毫升离心管。使用玻璃巴斯德吸管移除尽可能多的胚胎培养基.
注: 对年轻 (mid-somitogenesis) 胚胎进行固定和药物治疗, 在形成神经中心调解疼痛感觉之前, 不需要额外程序来减轻安乐死期间的疼痛。发展阶段的定义是在 et al ., 1995 25 。4-5 和11-12 节阶段用于获取 图 2 和 3 的图像。- 准备4% 甲醛 (粉煤灰)/MT 组装缓冲 (单克隆人) 固定剂 (参见 材料表), 将1毫升8% 的每1毫升2X 单克隆抗体组合在一起, 并添加2和 #181; 每100% 毫升 X-100 总体积.
注意: 在处理含有粉煤灰和海卫 X-100 的溶液时戴上手套, 这是皮肤刺激物.- 将1毫升4% 的胚胎固定在28.5 和 #176 的5分钟; c. 用吸管吸出定影液, 用1毫升新鲜定影液代替, 在摇杆上室温 (RT) 孵育 3 h.
注: 样品必须在其生物温度 (28.5 #176; 斑马鱼 C) 中快速固定, 以防止温度依赖性 MT 降解. - 准备4% 甲醛 (粉煤灰)/MT 组装缓冲 (单克隆人) 固定剂 (参见 材料表), 将1毫升8% 的每1毫升2X 单克隆抗体组合在一起, 并添加2和 #181; 每100% 毫升 X-100 总体积.
注意: 在处理含有 NP-40 的溶液时戴上手套, 皮肤刺激性.
- 热 RT 4% 低熔点 (LMP) 琼脂糖嵌入介质在一个封闭的容器, 直到解决方案变得清晰使用热板设置为50和 #176; C 定位接近解剖显微镜.保持容器在样品之间闭合并且在整个嵌入过程中加热 (步骤 1.4. 4-1. 4.6).
- 使用玻璃吸管将胚胎从1.5 毫升离心管转移到培养皿中, 然后用 1X TBS-NP40 填充.
- 在解剖显微镜的放大倍数下, 用细钳从 somitogenesis 阶段胚胎 (4-5 和11-12 节) 中取出大的卵黄细胞 26 。用一副钳子将胚用尾芽夹住, 并将蛋黄细胞与另一对一起剥开, 以保存后脑组织。将 de-yolked 胚胎转移到培养皿中无蛋黄碎片的区域.
注: 在琼脂糖填充模中嵌入胚胎, 以防止 LMP 琼脂糖的过早硬化. - 填充一个 12 mm x 5 mm x 3 mm 的剖切模具与200和 #181; 我用微融化了 LMP 琼脂糖执行步骤 1.4. 5. 1.4. 6。迅速 (在二十年代内填充模具) 在 LMP 琼脂糖冷却到 RT 和固化前植入胚胎.
- 用细钳将 tailbud 从培养皿中的 de-yolked 胚转移到琼脂糖填充的模具, 并在解剖显微镜下将其锥形端.
- 使用细钳将胚胎定向到模具中, 使 vibratome 在所需的平面上切割。通过定向胚胎来创建横断面, 使后脑组织与模具的长度平行, 其背表面朝向边缘, 其前表面朝向锥形区域末端。对其余的胚胎重复步骤 1.4. 4-1. 4.6.
- 允许琼脂糖嵌入在 RT 中固化5分钟.
- 生成40和 #181; m 部分的最高轴琼脂糖嵌入胚胎 (步骤 1.4. 1-1. 4.7) 使用 vibratome 与切片菜填充 1x TBS-NP40。将感兴趣的部分转移到500和 #181 的24井板; L 1x TBS-NP40 使用细钳。每井只放置一个胚胎的部分.
注意: 有关详细信息, 请参阅参考 18 。确保在至少250和 #181 的情况下始终保持水分, 以低速 (10-25 rpm) 的缓冲和岩石的 L 为剩余的步骤, 以防止分离琼脂糖嵌入。在堵塞和洗涤液中存在的洗涤剂应减少液体介质的表面张力, 并允许浸入部分。检查在所有操作期间和之后, 部分留在井中。请小心避免在洗涤过程中意外丢弃节.
注: 使用包含每种二次抗体宿主种类的5% 血清的阻断溶液 (参见 材料表 ).
注: 对总 mts (反和 #946;-蛋白, 或反和 #945;-蛋白) 和稳定 mts (反谷氨酸-蛋白) 或动态 mts (反 Tyr 蛋白) 的主抗体孵化的双标签部分。选择主要的抗体, 已提出在不同的寄主物种时, 双标记的总和 post-translationally 修饰和 #945; 蛋白的人口。有关抗体稀释, 请参阅 材料表 .
注: 从这一点开始, 在每次操作后, 用铝箔包裹含有二次抗体的井碟, 以防淬火。选择与主抗体的宿主免疫球蛋白反应的二级抗体.选择具有独立、不重叠发射光谱的二次抗体荧光。有关抗体稀释, 请参阅 材料表 .
注: 核标记为步骤1.12 中执行的 MT 量化提供了蜂窝上下文.
注: 在使用显微镜之前, 在幻灯片背面使用一个细尖的永久性标记来旋转部分, 有助于识别剖面.
- 在反向激光扫描共聚焦显微镜上安装剖面, 将幻灯片粘贴到舞台上, 片面向目标。确定控制幻灯片上适当的光学 (目标、激光和通道设置, 如增益和偏移), 并使它们在示例 27 之间保持一致。避免 oversaturating 像素以防止数据丢失.
- 使用选定的第二抗体荧光的通道设置捕获 Z 叠共聚焦图像, 并保存图像文件 27 。为每个部分获取 Z 堆栈.
注意: 使用以下获取设置复制用于获取 图 2 和 3 中的图像的参数: 模式 = XYZ;物镜放大倍数 = 63X 油浸透镜;目标数值孔径 = 1.4;Z 步 = 0.1 和 #181; m;Z 深度 = 16.23 和 #181; m. 使用以下通道设置: DAPI 励磁与20% 紫外线范围激光, 发射过滤器范围 = 430-480 nm, 光电倍增管 (pmt) 增益 = 525 V, 和 pmt 偏移 =-1.72%;448 nm 荧光 (参考 材料表 ) 励磁与 20% 488 nm 激光器, 发射过滤范围 = 493-573 nm, pmt 增益 = 689 V, pmt 偏移 =-0.2%;594毫微米荧光励磁与 32% 594 毫微米 laser, 发射过滤器范围 = 608-706 毫微米, pmt 增益 = 768 V 和 pmt 偏移量 =-6.8%. - 使用唯一的描述性文件名保存原始数据文件, 并在图像分析软件中创建用于编辑的副本.
- 使用公共域3维图像分析软件 ( 如 、ImageJ) 打开数据文件副本。检查每个通道是否显示为单个图像序列 (Z 堆栈).
- 使用以下菜单序列分割图像通道: 和 #8220; 图像/颜色/分割通道和 #8221;.
- 通过使用以下菜单序列覆盖感兴趣的通道来创建合并图像: #34; 图像/颜色/合并通道. #34; 选择 594 nm、488 nm 和 DAPI 通道分别为红色、绿色和蓝色。检查和 #34; 创建复合 #34; 选择和 #34; 确定 #34; 28 .
注意: 省略 DAPI 通道以在 图 2 和 3 的最大投影中更好地传递特定于 MTs 的详细信息, 方法是仅为其他两个通道选择假颜色. - 检查合并的 z 堆栈, 并记下所有可见通道的内部最佳 z 平面的开始和结束位置。关闭通常具有次优信号的外部 Z 平面, 这是由于剖面的不均匀表面造成的。有关详细信息, 请参阅参考 29 .
- 将合并的 z 堆栈可视化为单个2维图像, 方法是使用以下3维图像分析菜单序列执行 z 堆栈的最大强度投影: 和 #34; 图像/堆栈/Z 项目. #34; 输入内部最佳的起始和结束位置Z-平面从步1.11.3 作为和 #34; 开始切片和 #34; 和 #34; 停止切片, #34 分别。选择和 #34; 最大强度和 #34; 作为投影类型和点击和 #34; OK 和 #34;。有关详细信息, 请参阅参考 28 .
- 打开商业3维图像分析软件。选择和 #34; 创建库和 #34; 并为图像库提供描述性名称。单击并 #34; 创建. #34; 将从共焦显微镜生成的原始图像文件拖动到库中。较大的文件需要更多的时间来传输.
注意: 表 1 中设置的目标是首先设置一个阈值, 以丢弃其分布和大小与所分析对象的大小不一致的信号。例如, 在计算原子核时, 消除信号不太大, 不能成为原子核.
注: 平均 MT 丛长度 = 与 #39 的总和; #946 的平均值; 蛋白和 #39; 每个胚胎除以胚胎总数。请参阅 表 2 的20行。格式化电子表格, 使变量和实验组易于绘制.
2。 De 从头 MT 汇编分析 (协议 2)
- 在实验前2天构造和测试井流设备 ( 图 1 ).
注: 该仪器可在使用来自 材料表 的 nocodazole 处理后同时冲洗多个实验组。硅胶封口机需要至少24小时的干燥时间, 才不会对胚胎产生毒性风险。- 使用夹具或带锯将50毫升离心管分成半纵长.
- 切割7半圆, 半径为3厘米, 出70和 #181; m 尼龙网布和修剪他们, 以适应紧密的一半分裂离心管。胶水的半圆到离心管平行于10毫升级标记使用水族馆安全的硅封口机。允许设备干燥2天, 并通过浸泡在一个烧杯的水2-3 小时冲洗.
- 将切割离心管的顶部 (螺纹) 端与建模粘土相线, 使流经该流体装置的液体的高度具有深度和 #188; 英寸 ( 图 1 , 视图2和 3).
- 通过从50毫升注射器中取出柱塞并将12英寸的细管插入尖端, 准备冲洗装置。推动油管, 只要它会去和密封周围的连接使用建模粘土.
- 预湿网格使用胚胎培养基, 使液体贯穿整个流动装置。将装置放在冰面上, 使所有车厢内的液体池都能倒出, 而在粘土轮辋所在的前部仍可清空。使用环形支架, 将冲洗装置挂在冰上流经的设备上方 ( 图 1 ).
- 冷却200毫升的胚胎培养基在冰上, 倒入足够的冲洗装置, 以确保所有的气泡被清除, 流速约7毫升/分钟. 通过改变注射器的高度来调节流速.
- 酶 dechorionate 胚胎
- 在20毫升的胚胎培养基中稀释1毫升的10毫克/毫升非特异性蛋白酶, 从而制作出非特异蛋白酶的工作溶液.
- 在 timepoint 前1小时对胚胎进行化学 dechorionation, 期望它们达到预期的发育阶段。chorions 从100毫米培养皿中去除胚培养基, 加入20毫升的非特异性蛋白酶工作溶液.
- 在37和 #176 孵育胚胎; C 为 5 min.
注: 不超过5分钟或使用更高浓度的非特异性蛋白酶溶液, 因为这将导致的胚胎分裂一旦治疗与 nocodazole. - 用大约25毫升的胚胎培养基快速移出非特异性蛋白酶和填充皿。重复一次.
- 使用1毫升玻璃吸管, 将小于 24 hpf 的胚胎移植到玻璃器皿中, 以保护它们免受损坏.
- 完成 dechorionation 通过手动删除 chorions 使用一对优良的镊子, 如步骤1.1.5 中所述.
- 在28.5 和 #176 中放置含有 dechorionated 胚胎的玻璃培养皿; C 孵化器至少30分钟, 直到达到预期的发育阶段.
- 解现有 MTs
- 准备一个5和 #181 的工作解决方案; nocodazole 50 和 #181 的结合; 1 毫克/毫升 nocodazole 与10毫升冰冷胚培养基.
注意: 使用手套时, 处理 nocodazole, 皮肤刺激性. - 用10毫升冷 nocodazole 工作液交换 nocodazole 治疗组的胚培养基。将培养皿放在冰上, 为发育阶段 (例如, 4-5 节胚胎的1小时) 提供适当的时间。将未经处理的控制胚胎留在冰上的培养皿中, 并在步骤2.3.4.1 中与冲洗样品一起固定.
- 用火抛光1毫升玻璃吸管将胚胎移植到流经的设备, 每个实验组使用单独的隔间。启动 nocodazole 冲洗, 将冰冷胚培养基倒入50毫升注射器的顶部.
注: 每个实验组至少使用30胚胎。实验组可由控制胚胎或多种吗或 RNA 注入胚胎组成。冲洗将需要总共约150毫升的胚胎培养基将增加每8-10 分钟的 nocodazole, 同时继续抑制 MT 生长与冰。将胚胎保持在冰上对这种方法的成功至关重要, 因为 MTs 在低温下是不稳定的, 而且在早期胚胎中会出现冷的延迟. - 允许 MTs 在 RT 中经过20分钟的冲洗后再生长, 将胚胎移植到含有温热 (28.5-#176; C) 的玻璃培养皿中, 使用火抛光1毫升玻璃吸管。胚胎一旦转移就开始计时
- 将控制和冲洗胚胎在1分钟、5 min 和10分钟内, 由移大约10胚胎放入1.5 毫升的离心管中, 填充1毫升4% 的粉煤灰/单克隆 (28.5 和 #176; C), 并按照步骤1.2.3 中的指示进行操作.
- 准备一个5和 #181 的工作解决方案; nocodazole 50 和 #181 的结合; 1 毫克/毫升 nocodazole 与10毫升冰冷胚培养基.
- 为 immunolabeling 准备示例, 如1.3 至1.5 节中所述。
- Immunolabel 漂浮部分和图像胚胎, 如1.6 节-1.10 所述, 主要抗体规格如下: 使用1:500 兔抗 #947;-蛋白和1:200 鼠抗 #946;-蛋白.
- 步骤1.12 中所述, 使用3维图像分析软件处理和分析图像.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
基于 immunolabeling 的稳定动态 MTs 分析
在协议 1中, 利用谷氨酸-蛋白和 Tyr 蛋白作为稳定和动态 MTs 的标记, 揭示了神经管发育早期 (神经脊) 和晚期 (神经根) 阶段的 MT 亚的分布。动态 MTs 在神经龙骨阶段 (4-5 节) (图 2A-D) 的后脑中占主导地位。当龙骨发展成神经杆 (11-12 节), 增强上皮的阶段, 定性地较少 MTs 是免疫反应性与抗 Tyr-蛋白抗体 (图 2E H), 特别是在腹杆。相反, 谷氨酸-蛋白是分散和点状的整个神经龙骨 (图 3A-D), 但丰富的腹神经杆沿 MT 大片 (图 3E H)。箭头指向特定的 MT 束或标记增加的结构。
虽然抗谷氨酸-蛋白和抗 Tyr 蛋白抗体是在同一寄主物种 (防止双重标记实验), 这些结果表明, 稳定和动态 MT 标记很少重叠斑马鱼后脑。首先, 腹神经杆比动态 (图 2f) MTs 更稳定 (图 3f)。这一趋势在背神经杆反转, 与斑马鱼 neurulation 的模型一致, 在这种模式下, 背部组织保持动态, 直到神经管形成20。其次, 当有丝分裂纺锤体完全贴上神经龙骨的 Tyr-蛋白抗体 (图 2D, 箭头) 时, 只有主轴的底座, 与中心重合, 用稳定标记谷氨酸-蛋白 (图 3D, 箭头)。β-蛋白免疫荧光, 共同对两种化验, 通知实验者的分布的所有 MTs, 并提供了一个基础, 消除非特异性标记。
使用3维图像分析软件测量对象会产生大量数据, 可以将其组织成一个方便的表 (表 2)。要进行长度、计数和面积测量, 我们只使用可供分析的数据的子集。我们不进一步分析的数据的一个组成部分是标识的对象的数量。这个数字是作为一个内部质量控制, 因为这个数字不应该很大的差异, 在类似的部分, 和原子核的比例, MTs 应保持相似的一个单一的处理条件。"异常值" 是指需要使用调整后的滤镜重新运行分析, 或者图像的标记太差而无法进行分析的指示器。因此, 所有的离群图像应分析与调整设置。对于可能导致异常物体计数的标记或物理损伤的迹象, 应检查离群点部分。一旦分析完成和质量控制, 有用的信息可以从原始数据中恢复, 如总 mts 和稳定 mts 的平均长度或稳定 mts 与总 mts 的比率 (表 3)。除了这些测量之外, 还可以使用3维图像分析软件获得许多其他度量, 这些数据可以用来推断 MTs 或它们与其他蜂窝结构 (核、中心、等) 的关系。
De 从头MT 组件检测
nocodazole 处理聚 MTs 导致漫贴标记 (图 4A、4D、和4G)。随着 MTs 的再生, 它们从中心 (图 4B、4E、和4H) 扩展, 但是, 由于它们的平面轨迹 (图 4C、4F、和4I), 这在单个平面中可能并不明显。然而, 一些图像分析软件能够测量长度在3维, 使评估 MT 增长后, nocodazole 冲洗 (表 4)。从表 4中的数据集可以获得的一个重要观察是, 在分析了神经管的所有区域的 nocodazole 冲刷后, MTs 的平均长度似乎会随着时间的推移而增加。如上所述, 从3维图像分析软件获得的其他类型的度量可以提供单元格上下文来解释 MT 数据 (例如, 每个核的 MTs 比率)。
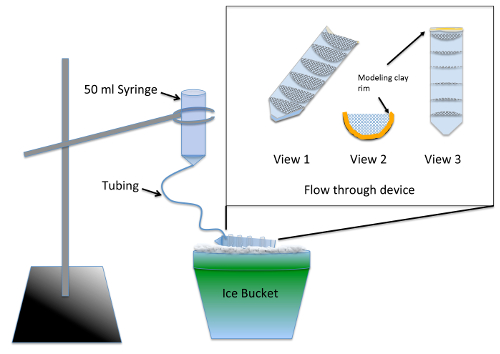
图 1: 用于的冲洗装置的图示de 从头MT 组件检测该插页是一个特写的流动通过的设备, 由网粘成50毫升离心管切割纵向。网格划分的流动装置, 使多个实验组可以同时处理。在使用过程中, 将胚胎培养基添加到注射器中, 并慢慢流经油管以填满流经的装置, 为所有实验组提供恒定冲洗。请单击此处查看此图的较大版本.

图 2:使用 immunolabeling 来图像动态 MTsDechorionated 胚胎是固定在适当的阶段 (4-5 在A-D和12-13 节在E H), 横切片通过后脑, 和 immunolabeled 抗体对β-蛋白 (绿色在a和e)要在B和F中标记所有 MTs 和 tyrosinated α-蛋白 (红色) 以显示动态 MT 群。在合并图像 (C、G) 及其更高的放大 (d、h) 中, 可以看到高动态 MTs, 这是黄色标签可见的区域 ( d、h中的箭头)。刻度条 = 25 µm (A-C 和 E-G) 和10µm (D 和 H)。请单击此处查看此图的较大版本.

f清楚 3: 使用 immunolabeling 图像稳定的 MTs.Dechorionated 胚胎是固定的, 通过后脑和 immunolabeled 在适当的阶段 (4-5 节在A-D 和12-13 节在E H)。稳定的 mts 被标记抗体反对 detyrosinated 形式α-蛋白 (谷氨酸-蛋白) (红色在B和F), 而总 mts 是形象化的以一般β-蛋白抗体 (绿色在a和E)。合并图像中的红色和黄色信号 (C、G) 及其较高的放大 (d、h) 表示高 MT 稳定性的区域 (d、h 中的箭头)。刻度条 = 25 µm (A-C 和 E-G) 和10µm (D 和 H)。请单击此处查看此图的较大版本.

图 4:使用 immunolabeling 来图像初始化的 MTs.Dechorionated 胚固定在4-5 节, 横切片通过后脑。immunolabeled 用β-蛋白 (D, E,和F) 来标记生长 MTs 和γ-蛋白 (A、B和C), 以标记成核点/中心。神经管的背部区域被装箱 (a, D;B、E和-f) 并以更高的放大倍数显示 (分别为G、H、I) 以显示细胞核 (DAPI、蓝色)、中心 (γ-蛋白、红色) 和总 MTs (β-蛋白、绿色)。白色箭头: MTs 和中心的定位;黄色箭头: 单元格的第二个中心可见。刻度条 = 25 µm (A-F) 和10µm (G I)。请单击此处查看此图的较大版本.

表 1: 3 维图像分析软件中筛选对象的默认设置。

表 2: 使用3维图像分析软件获得的具有代表性的原始数据集, 用于分析稳定的 MTs。每列表示来自单个节的度量值。最小测量量;最大尺寸:SD: 标准偏差;SE: 标准错误。

表 3: 可从3维图像分析软件获得的数据集的示例, 用于量化稳定的 MTs。通过取所有样本中相关标签的平均骨架长度平均值 (参见表 2) 和稳定的与总 mts 的比值, 来选择测量总 (β-蛋白) 和稳定 (谷氨酸-蛋白) mts 的平均值长度 (每β-蛋白条纹的谷氨酸-蛋白条纹) 计算的平均β-蛋白计数除以平均谷氨酸-蛋白计数。

表 4:可从3维图像分析软件中获得的数据集的示例分析de 从头MT 程序集。代表的结果来自de 从头MT 组装实验, 比较三恢复时间点 (1、5和10分钟) 获得的数据集, nocodazole 冲洗后。对于每个时间点, 为核计数获得的测量, 中心 (γ-蛋白点), 总 MTs 的数量 (β-蛋白条纹), 显示对被分析的被选定的区域 (开发的神经管的横断面)。
Subscription Required. Please recommend JoVE to your librarian.
Discussion
目前在斑马鱼的早期发展中有许多成像 MT 动力学的方法, 从标记分子的活体成像到固定组织的 immunolabeling11,12,13,14。虽然单个单元中的 mts 可以存在于动态或稳定状态, 但上皮是一个过程, 其中 mts 随着时间的推移而逐步稳定。使用稳定和动态 MTs 的标记提供了一种可视化这种现象的方法。这里提出的方法利用3维成像软件的威力来量化从动态到稳定的 MT 种群在胚胎斑马鱼组织的横断面上的转变。在协议 2中, 该方法用于标记新生 MTs 的不同种群, 并跟踪它们的成核和增长超时。
由于他们的解倾向, MTs 在他们的家乡很难想象。因此, 该方法的关键组成部分是快速固定在整个胚胎的 MTs。这是通过开始固定在生理温度和使用一个缓冲, 既稳定了 MTs 和增加了胚胎的渗透性。固定时间也很重要, 因为限制固定不能逮捕 MTs, 而 over-fixation 可以掩盖表位, 干扰抗体结合。建议的固定时间3-4 小时与胚胎, 在 mid-gastrulation 高达 24 h 胚乳。胚胎朝向更年轻的结束时间刻度应该被固定为接近 3 h, 而年长的胚胎可能需要整个 4 h。即使有适当的固定, MTs 将解与时间, 所以切片和 immunolabeling 必须发生在一个星期的修复。
一旦组织修复得当, immunolabeling 就会出现问题。最常见的问题是, 通过中心的组织, 特别是, 如果太多的部分是在同一井孵化。增加初级抗体的浓度和主、二级抗体的培养时间, 同时增加洗涤剂改善性的胚胎, 将改善大多数 immunolabeling 的问题。如果抗体标记由于固定问题或 immunolabeling 问题而失败, 可以通过检查抗体标记模式来确定病因。不良的固定将导致在细胞膜上的强烈标签和扩散标签的细胞质, 而 over-fixation 将导致弱标记, 保留 MT 的建筑。然而, 抗体的渗透率很低, 会出现在组织中心的区域, 没有标记。
有意义地分析 MT 图像的能力取决于高质量的成像。为了在3维中捕获 MT 长度, 应该使用最小的 Z 步长来实现目标和数值孔径。图中所示的图像是用63X 油再现物镜捕捉的, 1.4 数值孔径产生如下: 像素 = 240 nm, z 步 = 0.1 µm, z 堆栈大小 = 16.252 µm。因为单个 MT 的宽度是 25 nm, 大约是光显微镜分辨率的10倍, 所以不能用这种方法准确地测量这个指标。相反, 只有 MT 长度等于或大于在所有三维度中可达到的最小像素大小。线和/或帧平均可以提高 MT 信号的定义。MT 分析应预留高质量的部分。虽然不固定的组织不能成像和分析, 温和的 overfixation 可以抵消, 谨慎增加激光强度和增益, 以检测微弱的信号, 同时保持良好的动态范围。抗体穿透力差, 而不是最佳, 可以通过限制图像采集到 well-labeled 区域来纠正, 导致成像一个较薄的部分 (5-10 µm)。通过调整过滤器设置, 可以补偿来自标签的高背景。但是, 如果进行了这些调整, 则需要验证在 Z 堆栈的每个平面上是否可以接受筛选阈值。
3维图像分析软件允许实验者在固定组织部分的3维空间中量化 MT 长度、面积、角度、丰度和其他指标。此处描述的方法提供了使用商业可用软件获取此类数据的方向。然而, 过滤模块可以适应公共领域软件增强与相关的插件和/或宏, 使分析可供所有人。在分析之前, 必须值原始图像, 以避免在量化中包含背景和非特定信号。一旦分析完成, 并将数据转移到一个可行的电子表格中, 就可以从数据集进行许多推论。这里所做的计算之一是每β-蛋白条纹的谷氨酸-蛋白条纹, 或稳定 mts 与总 mts 的比值, 其中1表示整个 MT 骨架在 ROI 中稳定。如果实验者希望补充他们的定量数据, 生成一个抛光标记的图像文件格式 (TIFF) 图像与规模酒吧是毫不费力的3维图像分析软件。
这一分析允许的蛋白质的功能研究 MT 组装,在体内。如果在交替序列剖面上执行 immunolabeling, 该协议可用于研究同一胚胎中的动态和稳定的 MTs。在未来, 如增加洗涤剂或改变嵌入角度的修改, 将允许使用这些方法的老胚胎和更广泛的解剖问题。
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
作者没有什么可透露的。
Acknowledgments
共焦显微镜是在美国国家科学基金会 (NSF) 的资助下购买的, 赠款 #DBI-0722569。这项研究得到了美国国立卫生研究院/国家综合医学研究所 (NIH/研究院) 赠款 #GM085290 和美国国防部 (DOD) 赠款 #W81XWH-16-1-0466 授予 rm 布鲁斯特。重要的是通过前学院和本科生科学教育计划, 授予 #52008090 UMBC 从霍华德休斯医学院的赠款支持。某地布朗得到美国教育部 GAANN 奖学金的支持, 约瑟夫·迈尔霍夫研究生奖学金由 NIH/研究院补助金 #GM055036 资助, 以及由美国国防部拨款 #W81XWH-16-1-0466 资助的研究助理。
Materials
| Name | Company | Catalog Number | Comments |
| Agarose | Used to treat petridishes. Prepare 1% agarose by heating a solution of 1 gram agarose per 100 ml 1X embryo medium in a microwave until polymerized. |
||
| Kpipes | Sigma | P7643 | |
| NaCl | Sigma | S7653 | |
| Tris-HCl | Sigma | T3253-500G | |
| KCl | Sigma | P9333-500G | |
| CaCl2·2H2O | Sigma | C5080 | |
| NP-40 | American Bioanalyticals | AB01424 | |
| EGTA | Sigma | E3889-25G | |
| MgCl2 | Sigma | M2670-500G | |
| Bovine serum albumin (BSA) | Fisher | BP1605 | |
| Triton-x | American Bioanalyticals | AB02025 | |
| Anti-Fade mounting medium | Invitrogen | P10144 | |
| Mouse anti-β-tubulin | Developmental studies Hybridoma Bank | E7 | 1/200 |
| Rabbit anti-γ-tubulin | Genetex | GTX113286 | 1/500 |
| Rabbit anti-α-tubulin | Genetex | GTX108784 | 1/1000* |
| Rabbit anti-detyrosinated-tubulin | Millipore | AB3201 | 1/200-1/1000* Titrate antibody with first use of new lot. |
| Rabbit anti-tyrosinated-tubulin | Millipore | ABT171 | 1/500 |
| Mouse anti-centrin | Millipore | 04-1624 | 1/1000 |
| Goat 488 anti-rabbit | Thermofisher | A11008 | 1/500 |
| Goat 594 anti-rabbit | Thermofisher | A11012 | 1/500 |
| Goat 594 anti-mouse | Thermofisher | A11005 | 1/500 |
| Goat 488 anti-mouse | Thermofisher | A11001 | 1/500 |
| Vibratome | Vibratome | 1500 | |
| Forceps | World Precision Instruments | 555227F | |
| 100 mm petri dish | Cell treat | 229693 | |
| 35 mm petri dish | Cell treat | 229638 | |
| 50 ml falcon tube | Fisher | 14-432-22 | |
| Woven nylon mesh 70 um | Amazon.com | B0043D1SZG | |
| Micropipette | Gilson | F123602 | |
| Glass pipette | Fisher | NC-999363-9 | |
| Aquarium sealant | Amazon.com, by MarineLand | Silicone Sealer 1 oz (Tube) | |
| Ring stand | Fisher | 14-675BO | |
| Microbore PTFE Tubing, 0.022"ID | Cole-Parmer | WU-06417-21 | |
| Modeling clay | Amazon.com | Sargent Art 22-4000 | Any wax or oil based non-toxic modeling clay will suffice |
| Clamp | Fisher | 02-215-466 | |
| 60ml syringe | Fisher | 14-820-11 | |
| Embryo medium (E3) | 34.8 g NaCl 1.6 g KCl 5.8 g CaCl2·2H2O 9.78 g MgCl2·6H2O To prepare a 60X stock, dissolve the ingredients in H2O, to a final volume of 2 L. Adjust the pH to 7.2 with NaOH. Autoclave. To prepare 1X medium, dilute 16.5 mL of the 60X stock to 1 L. |
||
| Blocking Solution | 50 ml TBS-NP-40 2.5 ml normal goat serum 1 g BSA 625 µl Triton-X |
||
| TBS-NP-40 (pH 7.6) | 155 mM NaCl 10 mM Tris HCl 0.1% NP-40 |
||
| 2x MAB (pH6.4) | 160 mM KPIPES 10 mM EGTA 2 mM MgCl2 |
||
| Commercial 3-D Image processing Software | PerkinElmer | Volocity (V 6.2) | |
| Dry block heater | VWR | 12621-108 | Used as a hot plate to melt agarose in Protocol 1. |
| Dissecting Microscope | Leica | MZ12 | |
| Confocal Microscope | Leica | SP5 | |
| Flat embedding mold | emsdiasum.com | BEEM 70904-01 | |
| Public domain image processing software | NIH | ImageJ (V 1.5) | |
| * Success varies by lot number | |||
References
- Akhmanova, A., Steinmetz, M. O. Tracking the ends: a dynamic protein network controls the fate of microtubule tips. Nat Rev Mol Cell Biol. 9 (4), 309-322 (2008).
- Conde, C., Cáceres, A. Microtubule assembly, organization and dynamics in axons and dendrites. Nat Rev Neurosci. 10 (5), 319-332 (2009).
- Kaverina, I., Straube, A. Regulation of cell migration by dynamic microtubules. Semin Cell Dev Biol. 22 (9), 968-974 (2011).
- Kollman, J. M., Merdes, A., Mourey, L., Agard, D. A. Microtubule nucleation by γ-tubulin complexes. Nat Rev Mol Cell Biol. 12 (11), 709-721 (2011).
- Howard, J., Hyman, A. A. Growth, fluctuation and switching at microtubule plus ends. Nat Rev Mol Cell Biol. 10 (8), 569-574 (2009).
- Schulze, E., Kirschner, M. Dynamic and stable populations of microtubules in cells. J Cell Biol. 104 (2), 277-288 (1987).
- Gundersen, G. G., Kalnoski, M. H., Bulinski, J. C. Distinct populations of microtubules: Tyrosinated and nontyrosinated alpha tubulin are distributed differently in vivo. Cell. 38 (3), 779-789 (1984).
- Li, R., Gundersen, G. G. Beyond polymer polarity: how the cytoskeleton builds a polarized cell. Nat Rev Mol Cell Biol. 9 (11), 860-873 (2008).
- Asakawa, K., Kawakami, K. A transgenic zebrafish for monitoring in vivo microtubule structures. Dev Dyn Off Publ Am Assoc Anat. 239 (10), 2695-2699 (2010).
- Wühr, M., Tan, E. S., Parker, S. K., Detrich, H. W., Mitchison, T. J. A model for cleavage plane determination in early amphibian and fish embryos. Curr Biol CB. 20 (22), 2040-2045 (2010).
- Tran, L. D., Hino, H., et al. Dynamic microtubules at the vegetal cortex predict the embryonic axis in zebrafish. Development. 139 (19), 3644-3652 (2012).
- Butler, R., Wood, J. D., Landers, J. A., Cunliffe, V. T. Genetic and chemical modulation of spastin-dependent axon outgrowth in zebrafish embryos indicates a role for impaired microtubule dynamics in hereditary spastic paraplegia. Dis Model Mech. 3 (11-12), 743-751 (2010).
- Yoo, S. K., Lam, P. -Y., Eichelberg, M. R., Zasadil, L., Bement, W. M., Huttenlocher, A. The role of microtubules in neutrophil polarity and migration in live zebrafish. J Cell Sci. 125 (23), 5702-5710 (2012).
- Andersen, E. F., Halloran, M. C. Centrosome movements in vivo correlate with specific neurite formation downstream of LIM homeodomain transcription factor activity. Development. 139 (19), 3590-3599 (2012).
- Lee, S. -J. Dynamic regulation of the microtubule and actin cytoskeleton in zebrafish epiboly. Biochem Biophys Res Commun. 452 (1), 1-7 (2014).
- Bulinski, J. C., Gundersen, G. G. Stabilization and post-translational modification of microtubules during cellular morphogenesis. BioEssays. 13 (6), 285-293 (1991).
- Magiera, M. M., Janke, C. Chapter 16 - Investigating Tubulin Posttranslational Modifications with Specific Antibodies. Methods Cell Biol. 115, 247-267 (2013).
- Hong, E., Jayachandran, P., Brewster, R. The polarity protein Pard3 is required for centrosome positioning during neurulation. Dev Biol. 341 (2), 335-345 (2010).
- Westermann, S., Weber, K. Post-translational modifications regulate microtubule function. Nat Rev Mol Cell Biol. 4 (12), 938-948 (2003).
- Jayachandran, P., Olmo, V. N., et al. Microtubule-associated protein 1b is required for shaping the neural tube. Neural Develop. 11, 1 (2016).
- Nam, S. -C. Role of Tau, a microtubule associated protein, in Drosophila photoreceptor morphogenesis. Genes N Y N 2000. 54 (11), 553-561 (2016).
- Abal, M., Piel, M., Bouckson-Castaing, V., Mogensen, M., Sibarita, J. -B., Bornens, M. Microtubule release from the centrosome in migrating cells. J Cell Biol. 159 (5), 731-737 (2002).
- Delgehyr, N., Sillibourne, J., Bornens, M. Microtubule nucleation and anchoring at the centrosome are independent processes linked by ninein function. J Cell Sci. 118 (8), 1565-1575 (2005).
- Manning, J. A., Lewis, M., Koblar, S. A., Kumar, S. An essential function for the centrosomal protein NEDD1 in zebrafish development. Cell Death Differ. 17 (8), 1302-1314 (2010).
- Kimmel, C. B., Ballard, W. W., Kimmel, S. R., Ullmann, B., Schilling, T. F. Stages of embryonic development of the zebrafish. Dev Dyn Off Publ Am Assoc Anat. 203 (3), 253-310 (1995).
- Beck, A. P., Watt, R. M., Bonner, J. Dissection and Lateral Mounting of Zebrafish Embryos: Analysis of Spinal Cord Development. JoVE J Vis Exp. (84), e50703 (2014).
- FÖldes-Papp, Z., Demel, U., Tilz, G. P. Laser scanning confocal fluorescence microscopy: an overview. Int Immunopharmacol. 3 (13-14), 1715-1729 (2003).
- Ferreira, T., Rasband, W. S. ImageJ User Guide - IJ 1.46. , Available from: https://imagej.nih.gov/ij/docs/guide/ (2010).
- Z-functions - ImageJ. , Available from: https://imagej.net/Z-functions (2017).
