

Summary
環境毒素への暴露鋭く胚に影響を与えることができます。内分泌撹乱化学物質ビスフェノール類の神経系に悪影響を与えると知られているようです。ここで初期胚の毒素の機能の影響を研究する脊椎動物 (ニワトリ)の in vitro神経ネットワーク モデルを用いたプロトコルについて述べる。
Abstract
ビス フェノール、ビスフェノール A (BPA)、bis-フェノール-S (BPS) など、プラスチックおよび多数の日常製品の生産で広く使用されるエージェントを重合します。エストラジ オールのようなプロパティを持つ内分泌かく乱化合物 (EDC) として分類されます。内分泌かく乱化学物質、低用量で長期曝露は癌、行動障害、不妊症、初期の発育期間中に大きな脆弱性を含む様々 な健康の欠陥とリンクされています。BPA の神経機能の発達に及ぼす影響を研究するには、生体外で神経ネットワーク初期モデルとしてニワトリ胚脳から派生を使用しました。曝露がわかった BPA は具体的活動と同期をスパイクのネットワーク活動の開発に影響します。ネットワーク活動の変化は、分子標的薬または化合物と行動の結果への影響の重要なリンクです。多電極アレイ ネットワーク活動体外で薬剤の効果を研究する便利なツールがますますなっています。市場で利用できるいくつかのシステムがあるし、電極種類、電極の配列および分析ソフトウェアは、基本的な原則の質の数のバリエーションがありますが、データ取得が全体で同じです、さまざまなシステム。現在体外二次元文化の分析に限られていますが、これらの MEA システム ネットワークの活動生体内で脳スライスを有効に改良を行っています。ここでは、胚の露出と神経ネットワークの活動と同調、代表的な結果とともに記録の詳細なプロトコルを提供します。
Introduction
4, 4 '-イソプロピリデンジフェノールと呼ばれる、ビスフェノール A や抗酸化添加剤、BPA、さまざまなポリカーボネートで発見され感熱紙に至るエポキシ樹脂製品、Cd、および粉砕証拠ガラス内側にコーティング飲料缶。ビスフェノール A はエストロゲン1を模倣する内分泌撹乱剤であること知られているが、最後の 10-15 年間で毎日カジュアルな BPA 暴露による副作用に関する研究主出てきた。でも低レベルの BPA 暴露の影響は母親2,3の暴露により胚発生時など初期の発育期間の最も深遠です。子宮内への露出女性の胚の内分泌撹乱化学物質は、乳房がん4,5に腟からの増加の疾患感受性にリンクされています。動物の研究は、BPA への暴露は、非定型の脳の構造と機能異常動作6で明らかに実証しています。結果として、ヨーロッパと北アメリカでは、FDA を含む、最も規制機関で乳児の哺乳瓶で BPA の使用を禁止されています。規制に準拠するため、多くのメーカーは、4, 4'-sulfonyldiphenol またはビスフェノール S (BPS) に切り替えています。BPA と BPS、構造類似物と最近のレポートがエストロゲン転写7BPS の対等な効力を示した、という事実に基づく BPA を基準にこの化合物の毒性を検討することが重要です。
ここで私たちは脊椎動物モデル、ニワトリ胚を用いたニューロン回路網の BPA (と他の内分泌かく乱化学物質) の効果をテストするためのプロトコルについて説明します。体外培養ニューロンのシナプスを形成し、活動電位 (スパイクとも呼ばれる) を生成します。これらの文化のスパイクの活動は、多電極アレイ (MEA) システムを使用して記録できます。スパイクは、互いの 5 ms 内で発生したときに同期すると見なされます。最終的に同期初期のランダムな発火活動は神経ネットワーク8,9の開発の重要な機能です。さまざまなメソッドを使用して同期を測定ことができるし、いくつかのアルゴリズムは、文学9,,1011で説明。私たちの分析は、Paiva、同僚12は MEA の集録システムを駆動する録音ソフトウェア、統合によって開発されたアルゴリズムを使用します。鶏胚ニューロンの堅牢なスパイキング アクティビティは、ニューラル ネットワーク活動13に BPA の影響を研究するためのプロトタイプを提供します。スパイク活動を記録する多電極アレイを使用して、BPA 暴露が神経細胞のスパイク同期の9,14の開発を禁じることを見ました。ここでは、ニワトリ胚培養神経スパイク同期の開発に関する代表的な結果とともに文化のニワトリ胚ニューロンの BPA 暴露を勉強するため詳細な方法論を提供します。
Protocol
下記プロトコル (初期) の胚発生に伴う BPA 暴露の影響をテストが標準化されて、BPS、BPF、またはひよこの萌芽期ニューロンの他の EDC を使用変更する場合があります。
このプロトコルは、デラウェア州立大学の制度政策と鳥類胚の NIH のポリシーに従います。また、プロトコルは DSU の材料と化学物質安全性ガイドラインの確認で。
1. 素材のセットアップ
- 1 mM 原液 (10% のエタノールのミリリットル当たり 0.23 mg) を作るために 10% のエタノール (v/v) で BPA (m. w. 228.29) を解散します。Neurobasal 媒体でその後希釈液 (0.1 μ M、0.5 μ M、1 μ M、5 μ M、10 μ M) を行います。
注意: BPA は、環境の毒素および粉体を処理するときに注意が必要があります。 - 37 ° C で市販の卓上卵のインキュベーターでニワトリの卵を孵化します。E7 胚のため 7 日間卵を孵化させなさい。
- 3 に 4 h の 70% エタノールに浸漬して高さ定置滅菌 MEAs の必要な数を準備し、BSLII フードに滅菌水約 10 mL で 3 回すすぎ。
注: 電極は、8 x 8 のグリッドに配置された 64 のナノ多孔質白金電極を含みます。電極径が 30 μ m と電極間隔が 200 μ m。MEA の腐食につながる電極を殺菌のため変性エタノールを使用しないでください。 - (無菌状態を維持) にふた付け滅菌コンテナーの内部電極を配置し、テーブルの上のインキュベーターに移動します。55 ° C で少なくとも 6 時間焼くこの手順が適切な殺菌のため重要であり、温度は 60 ° C を超えないようにしてください。無菌性を維持し、さらに使用するまでストレージのための BSLII フードに Mea を含んでいる皿を移動します。
注: 我々 はプラスチック製のふたでオーブン安全ガラスのグラタン皿を使用し、表面は 70% のエタノールと乾燥のための BSLII フードの場所でそれを殺菌します。 - 細胞外マトリックス ECM ソリューションの約数。4 ° C でバイアルを一晩解凍し、-20 ° C で 100 μ 因数を凍結
注: ECM は冷凍で出荷されています。ECM 準備ができるまで [コートに室温で温めましたら、重合、一度コートに不可能だと部屋の温度にしないくるべきだった。成長因子なし ECM は未知の要因のための追加効果を防ぐために好まれる。 - (デュモン #5 鉗子、カーブタイプ鉗子、ハサミ、春はさみ) すべての解剖器械を滅菌と自己シール材 70% エタノールを解剖皿のコーティングし郭清のフードで乾燥させます。
2. ひよこ萌芽期ニューロン文化とネットワーク活動
- めっきの日、バイアルを削除-20 ° C のフリーザーから ECM の 70% エタノール スプレーし、氷の場所します。BSLII フード内 300 μ L 冷たい neurobasal 培地を追加することによって 25% に希釈します。
- P200 を使用して pipetteman は、電極に触れるし、表面に薄い膜を残してすぐに削除するように注意して、MEA の中心に 25% の ECM の 100 μ L を追加します。MEA をカバーし、ニューロンをプレートに準備が整うまで CO2インキュベーター (37 ° C、5% CO2) に置きます。
- E7 卵 (胚は日 7、37 ° C で 7 日間培養) の外殻を 70% エタノールで滅菌します。冷滅菌ハンクのバランスの取れた塩のソリューション (HBSS) カルシウムなしを含む Sylgard 底解剖皿に胎児の首をはねます。目の周りカット、目玉を取り外します。鉗子・ デュモン #5 を使用して罰金して前脳とファイバーの蓋を公開する皮膚の外側の層を削除春はさみ腹側に切開を行います。皮をむくし、軟膜を慎重に削除します。もう一つのシャーレに脳を転送し、春ハサミで約 2 mm の小さな断片にそれをカットします。
注: が軟膜を除去した後、脳に接続されているすべての血管をしないでください。可能であればだが滅菌郭清のフードでは、解剖を行うことが好ましいがフード外郭清を実行すると、楽器と郭清範囲は徹底的に 70% のエタノールで滅菌されている限り、かなり良い結果を得た. - 転送滅菌ピペットを使用して、15 mL 遠心管に前脳の部分を収集し、遠心管の底に前脳シンクの部分をさせることの後できるだけ多くできるだけ HBSS の削除します。
- 0.05 %1 mL を加えてトリプシン/EDTA (37 ° C に加温) 37 ° C で 15 分間インキュベートし、。
- パスツール ピペットを使用して、慎重に組織の部分を乱すことがなくトリプシンを削除し、1 mL neurobasal 媒体を追加します。底にティッシュ シンクの作品を聞かせて、媒体を取り外します。この洗浄をもう一度繰り返します。この手順は、トリプシン/EDTA を洗浄することです。
- Neurobasal 培地 2 mL を追加し、triturating を開始します。製粉は、滅菌ファイアーポリッシュ パスツール ピペットを取って渡すそれを介して組織数回優しくティッシュのないのより多くの部分を見るまで含まれます。繰り返しのパスの後に任意の部分が残っている場合、底に沈殿するがまま。
注: は、triturating - 場合は泡が形成、停止および吸引により削除しながら泡を避けてください。 - 再浮遊細胞 1:10 neurobasal 媒体を希釈し、トリパン ブルー色素と診断を使用して実行可能なセルを数えます。トリパン ブルー溶液 0.5 mL 遠心管中の 50 μ L で希釈した細胞のミックス 50 μ。Coverslip 上に置いた後、血球に 10 μ L を追加し、明るい明確な細胞 (青色のセル死んでいるし、カウントすべきではない) を数えます。
注: 1 と 5 x 107セル/mL の間単一の E7 こま範囲から典型的な細胞の数。 - プレート、解離 ECM のセルは 2,200 セル/平方ミリメートルの密度で測定をコーティングします。これは、MEA 体制 MEA あたり約 130,000 セルにということを意味します。Neurobasal 培地 1 mL に、MEA のボリュームをもたらすと細胞接着や神経突起の伸展 (図 1) のために夜通し CO2インキュベーターで Mea を配置するを追加します。
注: さまざまな分野、電極数の測定、異なるセル密度が試みたこと - 一般的には、細胞密度が高い大きいネットワーク活動の結果、また短い住まれていた文化に 。 - 次の日は、手順 1.1 処理 MEA に 1 mM の在庫から neurobasal 中 10 μ M の最終的な集中に BPA を追加します。対照 MEA、同じ量の 10% エタノール水溶液 (v/v) に追加します。
- ネットワーク活動が増加すると 7 日間の in vitro (7DIV)、その後、毎日まで 10 μ M BPA 一日おきを含む neurobasal 培地で過ごした交換媒体。
注: ネットワークの活動が大きくなる代謝活性を増加させるし、メディアの変更より頻繁にする必要があります。オレンジを回す媒体をてはいけないです。
3. 神経ネットワークの活動の記録
- 取得の日、新鮮な neurobasal 培地で培養液を交換し、録音の前に 2 時間の最小値のための CO2インキュベーターに電極を返します。録音ソフトウェアを起動し、温度のアイコンをクリックして、MEA の温度を 37 ° C に設定 (補足図 3.1a-b)。
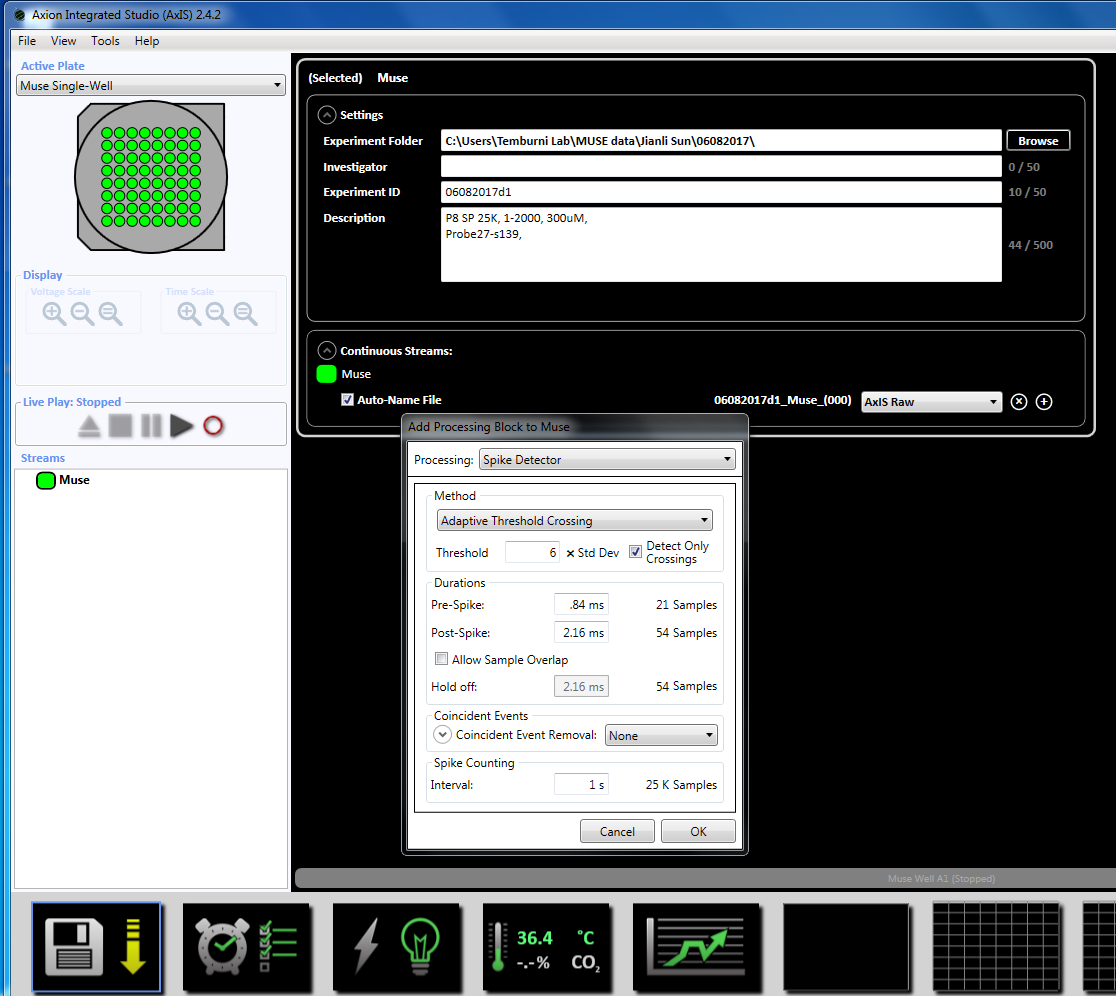
注: 録画めっきの 3 日目には早く始めることができますが、文化が生きている限り続けることができます。 - (左のウィンドウ パネルの [ストリーム) の下の「ミューズ」アイコンを右クリックして集録パラメーターを設定「追加処理」と「スパイク検出器」を選択し、ポップアップ ウィンドウで [ok] をクリックします。「スパイク検出器 (6 x STD)"ファイル名の下に表示されます (補足図 3.2a-b)。
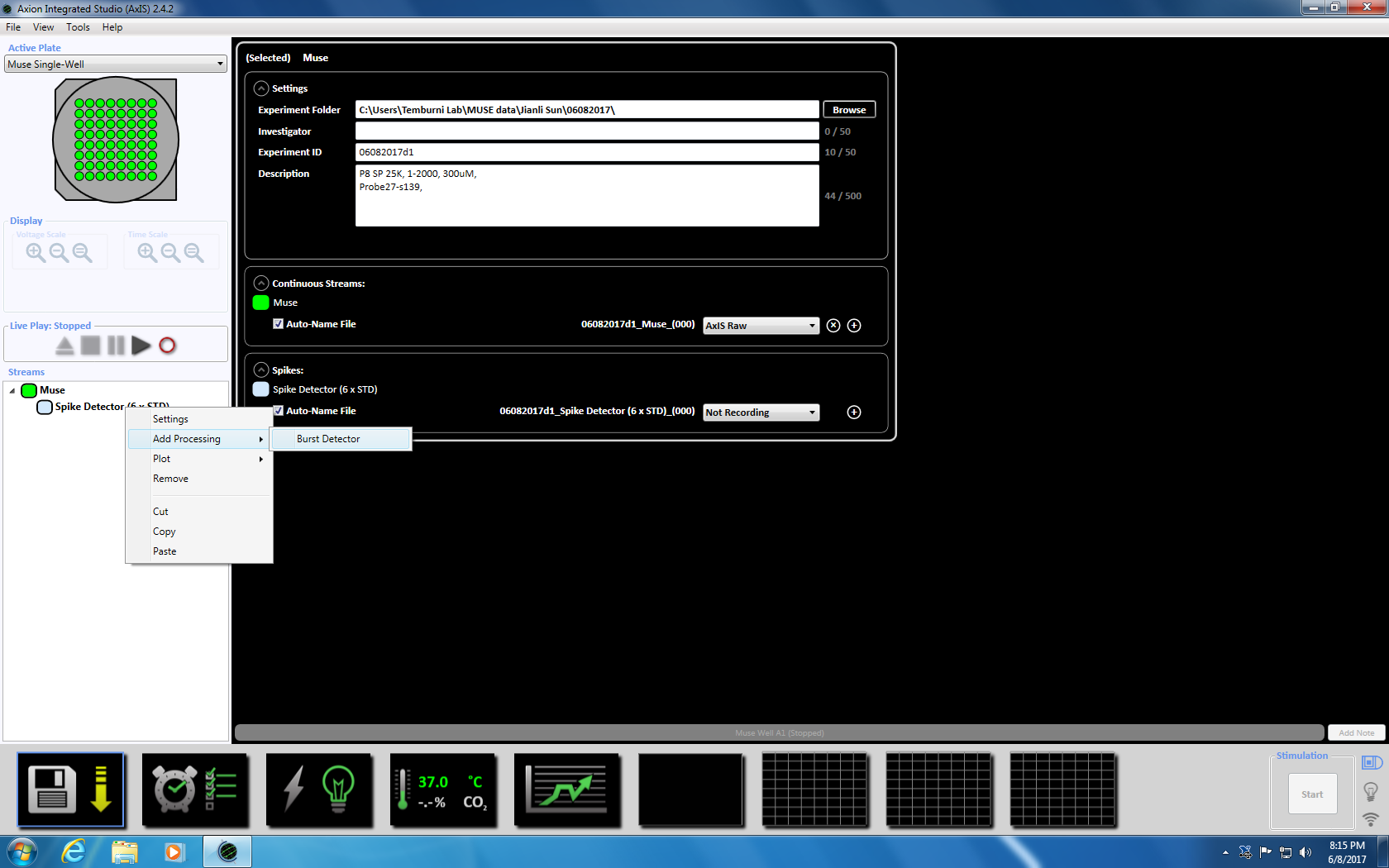
- 次スパイク検出器を右クリックして、「追加処理」と「バースト検出器」を選択し、ポップアップ ウィンドウで [ok] をクリックします。「バースト検出器」(ISI) はミューズのアイコンの下に表示されます (補足図 3.3a-b)。
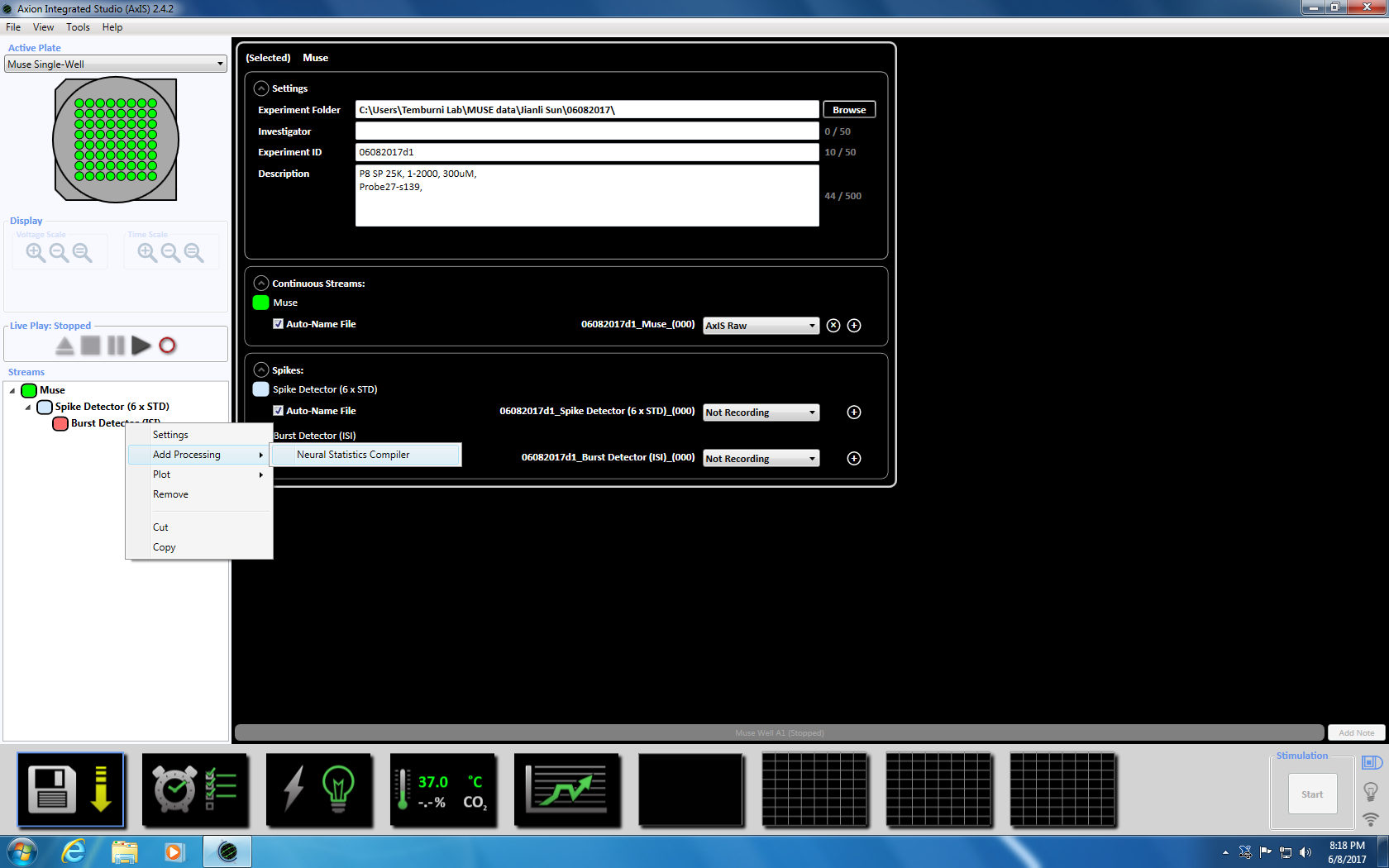
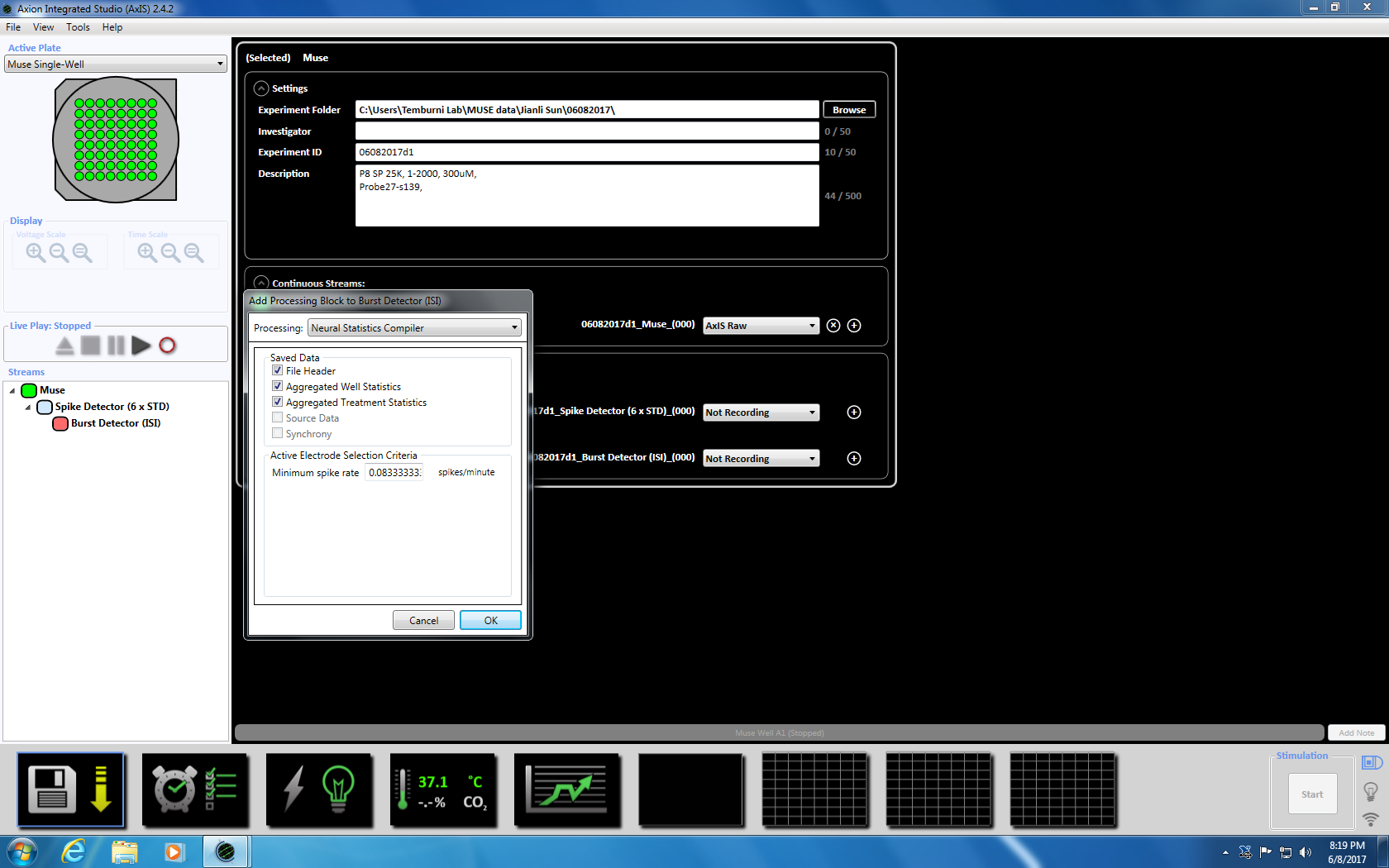
- 次にバースト検出器を右クリックして、「追加処理」と「神経統計コンパイラ」を選択ポップアップ ウィンドウで、[ファイル ヘッダー、よく統計情報を集計および同期が選択されていることを確認します。[Ok] を。「統計コンパイラ」が下に表示されます (補足図 3. 4a-b)。
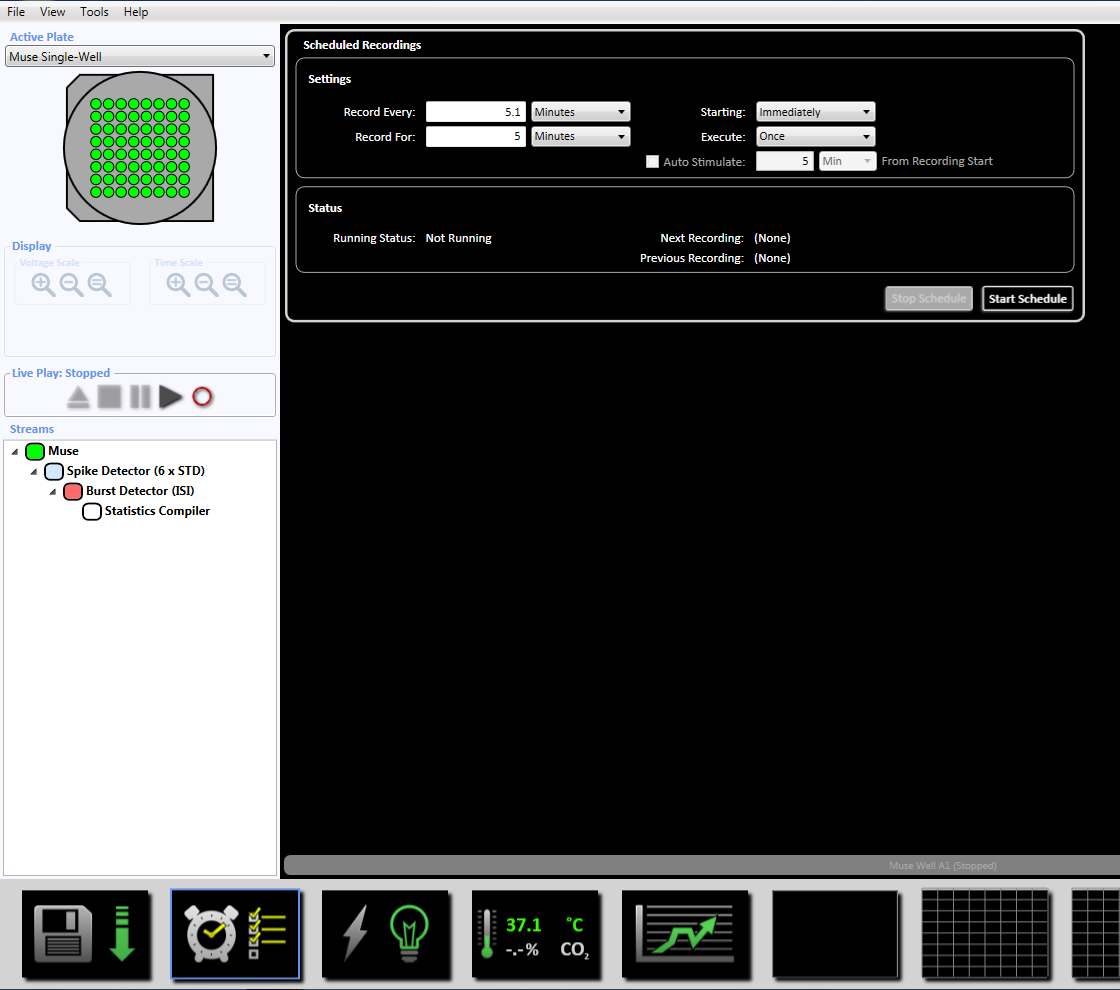
注: これは適応的しきい値交差 (6 x STD フィルター)、5、平均発火率検出 10 スパイク数の最小 100 ms スパイク間隔を設定 20 ms で同期パラメーターとあたり最小 0.083333 スパイクで最小スパイク率 s。 - 下部に時計のアイコンをクリックして 5 分 (300,000 ミリ秒) の録音時間の予約録画を設定します。これは、5.1 分ごとを記録し、5 分間記録設定] セクション内の情報を変更することによって実現されます。これはすぐに開始されます、一度実行されます (補足図 3.5)。
- MEA を移動する前に、MEA の温度が 37 ° C に達していることを確認します。インキュベーターから MEA を削除、記録ユニット上に配置し、MEA をロックします。ネットワーク アクティビティを予約録画セクションの開始レコードまたは「ファイルの再生」ウィンドウでレコードのアイコンをクリックしてして録音を開始します。通常、5 分 (300,000 ミリ秒) の録音時間は最大 10 分間のより長い録音を得ることができるが、十分です。
- 録音後、MEA をインキュベーターに戻ります。
注: する必要があります注意無菌性を維持し、MEA はインキュベーター外時間を最小限に抑えます。
4. データ解析
- 生データの分析はオフライン記録ソフトウェアを使用して行うことができます。ファイルのプルダウン メニューをクリックし、録音を開きます。分析する生データ ファイルを選択します。必要なスパイキング パラメーターをキャプチャする生録音を再生します。
- スパイクの平均数を取得し、スパイク検出器モジュールを使用して、同期のインデックスを取得するため神経統計コンパイラ モジュールを使います。
- スパイク検出器、バースト検出器と統計コンパイラ モジュール (セクション 3) 前に、をロードします。
- スパイク検出器、バースト検出器と統計コンパイラ モジュール ウィンドウ内にあるプルダウン メニューからそれぞれスパイク、ネットワーク バースト リスト、および高度な指標を選択します。
- 指示されたフォルダーで .csv ファイルを作成するデータ ファイルの録音を開始するレコードをクリックします。スパイク パラメーター スパイクおよび同調のインデックスの合計数は、.csv ファイルに記録されます。
メモ: プロセッサ時間と計算資源を消費する可能性がありますこれと、raw データを記録しながら、リアルタイム解析は推奨されません。
Representative Results
BPA のニワトリ胚神経細胞培養における同調性開発に及ぼす影響を検討しました。アクティビティのスパイクの同期は、神経ネットワーク体外の正常な発達の指標です。お互いの 5 ms 内で 2 つのスパイクが発生すると、彼らは同期と見なされます。生体外で神経文化最初に表示ランダム スパイク活動-スパイクがランダムに発生し、スパイク間間隔が正規分布を示します。文化が成熟し、スパイクのアクティビティの同期になるより、スパイク間間隔は一定であります。文化の同期発火活動に同調インデックス (SI) を導出する録音ソフトウェア12を使用してスパイクの相互相関解析による測定しました。スパイクの活動と同調インデックス (図 2と2 b) を減らして BPA 暴露がネットワーク アクティビティの開発に影響を与える悪影響ことがわかった。要約すると、我々 は文化でひよこニューロンの同期開発によって評価することができます神経の機能を含む、その開発を胎児の BPA 暴露の有害な影響に影響を示しています。

図 1:解離、めっき、およびネットワーク上のアクティビティを記録するひよこ前脳の解剖から手順を描いたプロトコルの概略図。この図の拡大版を表示するのにはここをクリックしてください。

図 2:(A)スパイクの数の平均値は有意に低かった BPA 扱わ E7 ひよこ前脳文化制御文化と比較されたとき。スパイクの平均数は、録音ソフトウェアを使用して抽出しました。手段 (コントロール、14,890 ± 949.0、N = 15 と BPA、5,624 ± 465.9、N = 22) 有意差 (対になっていない t 検定, p < 0.0001)。(B)平均同期インデックス (SI) が有意に低かった BPA 扱わ E7 ひよこ前脳文化制御文化と比較されたとき。SI は、レコーディング ソフトウェア内神経統計コンパイラ モジュールを使用して抽出しました。SI 手段 (コントロール、0.5159 ± 0.06547 N = 15 と BPA、0.1140 ± 0.01840 N = 22) 有意差 (対になっていない t 検定, p < 0.0001)。エラーバーは標準偏差を表しています。この図の拡大版を表示するのにはここをクリックしてください。
Discussion
急速な成長の胚など開発フェーズは、BPA を含む様々 な内分泌かく乱化合物の有害な影響を特に受けやすい。体外の脊椎動物の神経ネットワーク モデルの BPA 暴露の影響を調べるため詳細なプロトコルを提供します。BPA を開発神経回路網 (図 2の b) の同期のインデックスと同様に、スパイク率下げることがこのプロトコルを使用して、設立
我々 の方法論は初期胚の BPA 暴露の研究に設計され、他の内分泌かく乱化学物質の効果を研究に簡単に合わせることができます。ひよこから神経胚の文化は、ラットやマウスなど他の脊椎動物モデルに比べて確立する比較的簡単です。また、別の動物室の必要はありません-実験室ベンチに単純なインキュベーターはこれらの試金を遂行するのに十分な。EDC は、BPA のネットワーク活動に及ぼす影響を評価するため多電極システム (MEA) の使用について述べる。ここで説明されているプロトコルは、他のシステムに適用できます。このプロトコルの重要な側面は、不妊のメンテナンスです。これは無菌条件下で胚の解剖が含まれています-70% エタノールと滅菌のハンクのバランスの取れた塩溶液の使用楽器を殺菌で十分です。週間の期間にわたってデータが収集される、電極の滅菌処理を維持するために重要です。一度確立された神経文化が培養する長期 - 最大 3ヶ月-、内分泌かく乱化学物質と神経ネットワークの他の化合物への長期露出を調べる場合に役立ちます。このプロトコルの別の重要な側面は、郭清からめっきまでの時間です。最適な時間は 30 分、トリプシンとティッシュの孵化を含みます。長い所要時間、細胞の予後。
ここで任意の化学的または行動と神経機能に影響を与える薬剤の評価に適用できる基本的なプロトコルについて述べる。ここでは、我々 は 2,200 mm2あたりのセル密度を使用しています。ただし、これは変更することができます、他の細胞の密度を使用できます。一般的には、細胞密度の増加増加するネットワークの活動 - 短い時間でより多くのスパイクがわかりました。メソッドは、説明、評価ネットワーク活動に及ぼす化学薬品に非常に役立ちますが、制限があります。メソッドの最大の制限の 1 つはこれらの培養される 2 次元は脊椎動物の脳の三次元のアーキテクチャを反映していない可能性があります。これは、スライスの録音を使用して解決できます。別の方法としてはによって、ニワトリ胚に治療法を適用する卵15の広範な最後のカットの小さいウィンドウの治療レジメンの終わりに脳を解剖、vibratome の厚いセクションの MEA の上に置くネットワーク アクティビティの記録です。
私達のプロトコルはネットワーク活動の発達に及ぼす内分泌かく乱化学物質の試験を有効に、これらの化学物質の影響の機構基礎の調査を提供しています重要です。
Disclosures
著者は、彼らは競争の興味があることを宣言します。
Acknowledgments
本研究は、NSF (HBCU アップ研究開始賞、HRD 1401426 と EPSCoR EPS 0814251) と NIH (COBRE 1P20GM103653 01A1) によってサポートされます。K. s. は、デラウェア州 INBRE III 6404 から交わりによってサポートされます。
Materials
| Name | Company | Catalog Number | Comments |
| #5 foreceps | Fine Science Tools | 11251-10 | |
| Axion Muse MEA | Axion Biosystems | M64-GL1-30Pt200 | Will be called MEA system in manuscript |
| Axis Software | Axion Biosystems | Will be called recording software in the manuscript | |
| BPA | Sigma-Aldrich | 239658-250g | |
| curved forceps | Fine Science Tools | 11272-50 | |
| EtOH | Sigma-Aldrich | 64-17-5 | |
| Fertilized chicken eggs | from any local farm or Spafas | ||
| HBSS | Fisher | 14170112 | |
| Hemacytometer | Fisher | 02-671-6 | |
| Matrigel Growth Factor Reduced, Phenol Red-Free | BD Biosciences | 356231 | Will be called Extra Cellular Matrix (ECM) in the manuscript |
| Neurobasal medium | BrainBits | Nb4-500 | |
| Neuroexplorer statistical software | Nex Technologies | Neuroexplorer version 5 | |
| Pasteur pipettes | Fisher | 13-678-20A | |
| spring scissors | Fine Science Tools | 15514-12 | |
| Sylgard bottom dissection dishes | Living Systems Instrumentaion | DD-90-S-BLK-3PK | |
| Trypan Blue dye | Fisher | 15-250-061 | |
| Trypsin-EDTA | Fisher | 15400054 |
References
- Anahara, R., Yoshida, M., Toyama, Y., Maekawa, M., Masayuki, K., Ishino, F., Toshimori, K., et al. Estrogen agonists, 17 beta-estradiol, bisphenol A, and diethylstilbestrol, decrease cortactin expression in the mouse testis. Arch. of Histol. Cytol. 69 (2), 101-107 (2006).
- Dodds, C. Synthetic oestrogens. Br. Med. Bull. 11 (2), 131-134 (1955).
- Grignard, E., Lapenna, S., Bremer, S. Weak estrogenic transcriptional activities of Bisphenol A and Bisphenol S. Toxicology In Vitro. 26 (5), 727-731 (2012).
- Herbst, A. L., Ulfelder, H., Poskanzer, D. C. Adenocarcinoma of the vagina: association of maternal stilbestrol therapy with tumor appearance in young women. New Eng. J. Med. 284, 878-881 (1971).
- Jenkins, S., Raghuraman, N., Eltoum, I., Carpenter, M., Russo, J., Lamartiniere, C. A. Oral exposure to bisphenol A increases dimethylbenzanthracene-induced mammary cancer in rats. Environ Health Persp. 117 (6), 910-915 (2009).
- Okada, A., Kai, O. Effects of estradiol-17beta and bisphenol A administered chronically to mice throughout pregnancy and lactation on the male pups' reproductive system. Asian J Androl. 10 (2), 271-276 (2008).
- Palanza, P., Gioiosa, L., Vom Saal, F. S., Parmigiani, S. Effects of developmental exposure to bisphenol A on brain and behavior in mice. Environ. Res. 108 (2), 150-157 (2008).
- Mimoto, A., Fujii, M., Usami, M., Shimamura, M., Hirabayashi, N., Kaneko, T., et al. Identification of an estrogenic hormone receptor in Caenorhabditis elegans. Biochem. Biophys. Res. Commun. 364 (4), 883-888 (2007).
- Allard, P., Colaiacovo, M. Bisphenol A impairs the double-strand break repair machinery in the germLine and causes chromosome abnormalities. Proceedings Natl. Acad. Sci. 107 (47), 20405-20410 (2010).
- Mersha, M. D., Patel, B. M., Patel, D., Richardson, B. N., Dhillon, H. S. Effects of BPA and BPS exposure limited to early embryogenesis persist to impair non-associative learning in adults. Behav. Brain Funct. : BBF. 11, 27 (2015).
- Chen, Y., Shu, L., Qiu, Z., Lee, D. Y., Settle, S. J., et al. Exposure to the BPA-Substitute Bisphenol S Causes Unique Alterations of GermLine Function. PLOS Genetics. 12 (7), 1006223 (2016).
- Shein Idelson, M., Ben-Jacob, E., Hanein, Y. Innate Synchronous Oscillations in Freely-Organized Small Neuronal Circuits. PLoS ONE. 5 (12), 14443 (2010).
- Baltz, T., Herzog, A., Voigt, T. Slow Oscillating Population Activity in Developing Cortical Networks: Models and Experimental Results. J. Neurophysiol. 106 (3), 1500-1514 (2011).
- Pettmann, B., Louis, J. C., Sensenbrenner, M. Morphological and Biochemical Maturation of Neurones Cultured in the Absence of Glial Cells. Nature. 281 (5730), 378-380 (1979).
- Zhang, H., Wu, C. Y., Wang, W., Harrington, M. A. Interneuronal Synapses Formed by Motor Neurons Appear to Be Glutamatergic. NeuroReport. 22 (16), 809-813 (2011).
- Paiva, A. R. C., Park, I., Príncipe, J. C. A Comparison of Binless Spike Train Measures. Neural Comp. and Appl. 19 (3), 405-419 (2010).
- Cretu, A., Fotos, J. S., Little, B. W., Galileo, D. S. Human and Rat Glioma Growth, Invasion, and Vascularization in a Novel Chick Embryo Brain Tumor Model. Clin Exp Metastasis. 22, 225-236 (2005).
- Chiappalone, M., et al. Dissociated cortical networks show spontaneously correlated activity patterns during in vitro development. Brain Res. 93, (2006).
- Li, X., et al. Long-term recording on multi-electrode array reveals degraded inhibitory connection in neuronal network development. Biosens Bioelectron. 22 (7), 1538-1543 (2007).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}