The results obtained by DLS measurements during the crystallization experiment show a detailed evolution of the hydrodynamic particle radii resulting in two main particle distribution fractions, which develop over time. In the first part of the experiment, the sample was slowly evaporated, in order to achieve a higher protein concentration. As shown in Figure 5B, the protein drop was concentrated from 14 mg/mL to 19.5 mg/mL. During this time, according to the radius distribution pattern in Figure 5A, the protein shows a monomeric behavior in solution with a constant particle size of approximately 2.5 nm. As the sample is being further evaporated, the monomeric protein remains stable in solution, without any signs of denaturation or sample aggregation.
The addition of precipitant solution to the protein drop is immediately initiated after sample evaporation and highlighted with grey in the radius distribution pattern and balance curves for better visualization (Figure 5). Based on the calculations derived from the sample weight, the highlighted particle fraction is attributed to initiation of crystal nucleation, resulting in a radius size of approximately 200 – 400 nm. This phenomenon (known as nucleation) is initiated at a precipitant concentration of 0.6 M in the protein droplet, at a time period of approximately 66 min from the initiation of the experiment and approximately 23 min in respect to the initiation of precipitant addition. As the concentration of the precipitant increases, the radius distribution shows a broad distribution of particles in solution. As more nuclei form, the initial crystalline entities are growing in size, reaching a radius distribution between 800 – 1,300 nm. It can be concluded that at this stage, the crystal nuclei continue to grow, and the protein fraction is gradually becoming poorer as the protein molecules are taken-up by microcrystals. As the precipitant concentration in the protein drop slowly increases, the formation of microcrystals is easily identified after 75 min, when the radius distribution continues to develop between 500 and 1,500 nm. The evolution of the radius distribution is also confirmed by CCD camera images, where protein microcrystals are visible at a precipitant concentration of 0.7 M in solution (Figure 7). As the precipitant addition is finished and the crystallization drop is kept constant, the radius distribution shows a predominant phase between 1,000 and 3,000 nm, while the fraction of nucleation events diminishes over time. At this moment, the crystallization drop is fully saturated with microcrystals and no further nucleation events are present in solution.
Using as experimental input the feedback data given by the microbalance, a crystallization phase diagram was drawn for a comprehensive understanding of the thaumatin crystallization process. In Figure 6, the experimental phase diagram shows three independent crystallization experiments, where the production of thaumatin T. danielli microcrystals is labeled as THM_2 Micro-crystals (Protocol). Two additional crystallization experiments labeled THM_1 Macro-crystals and THM_3 Macro-crystals have been used as input data in order to have a correct mapping of the different areas in the phase diagram (e.g., solubility or metastable region). Since the protocols for these experiments follow different crystallization pathways, the identification of the nucleation region becomes easier, and hence more accurate. For each experiment, the crystallization path is highlighted by numbers of order to emphasize the different approaches and number of crystallization steps that were used for a specific outcome, while the grey arrows represent an estimation of the final protein concentration when crystal growth uptakes protein molecules from solution in the formation of well-defined stable protein crystals.
The three thaumatin experiments presented in Figure 6 show different conditions when entering the nucleation region, and hence different final crystallization outcomes as it can be seen from the pictures below the phase diagram. In the case of THM1 and THM3, the protein solution passes the nucleation phase and re-enters the metastable region, where it rests until crystals form. Consequently, the experiments lead to large, well-defined shaped crystals surrounded by mother liquor. However, for the experiment presented in the protocol section, THM2_micro-crystals, the crystallization path follows a particular approach. In a previous section we mentioned that for a sample to give microcrystals, the protein solution has to be located deep in the nucleation phase, so that the nucleation events can form at a maximum rate. In the case of THM2_micro-crystals, the ratio of protein to precipitant concentration was adjusted in such a fashion that the sample not only enters the nucleation region, but as a precipitant is further added to the protein solution, and the conditions do not move to another region in the phase diagram, but remain in a highly supersaturated state within the nucleation region. As a result, the entropy of the solution decreases drastically as the protein droplet becomes immediately saturated with small crystalline entities.

Figure 1. Schematic representation of the crystallization experiment. The drawing shows an overview of the crystallization experimental chamber with all the technical parts required for conducting an automated crystallization experiment. Please click here to view a larger version of this figure.
Figure 2: Software window for DLS and sample parameters that are relevant for a crystallization experiment. Parameters include temperature, relative humidity, viscosity, etc.
Figure 3: Software control window for the micro-dosage systems involved in the crystallization experiment. The features allow adjustment of specific parameters for the generation of droplets or solution stream.
Figure 4: Software window for the schedule table describing the crystallization steps involved in the experiment. The initial conditions of the crystallization droplet are also integrated in this window.
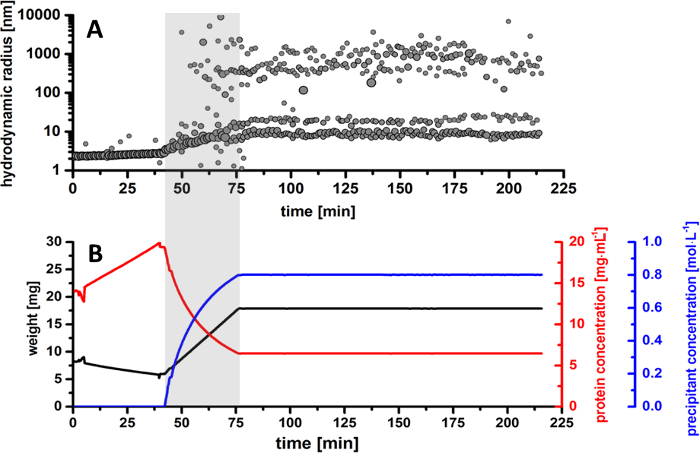
Figure 5. Overview of thaumatin T. daniellii microcrystals production. (A) Radius distribution of particle size in the protein drop during the entire crystallization process. (B) Monitored overview of experimental parameters. The plots represent the evolution over time for the weight of the protein droplet (black curve) together with the calculated protein concentration (red curve) and precipitant concentration (blue curve). Please click here to view a larger version of this figure.

Figure 6: Experimental crystallization phase diagram for thaumatin T. danielli together with the resulting outcomes. All plots are derived from experimental data, based on the feedback information given by the microbalance. The numbers attributed to each experiment that are visible between brackets represent the order and the number of steps taken during a crystallization protocol. Please click here to view a larger version of this figure.

Figure 7: Recorded photograph for THM2_Micro-crystals (Protocol) showing an abundant number of microcrystals saturated in solution. The picture was taken at 4 h (240 min) after setting the protein droplet in the experimental chamber for crystallization. Please click here to view a larger version of this figure.

Figure 8: Recorded photograph for THM_1 Macro-crystals showing a few large thaumatin crystals stable in solution. The picture was taken 20 h after setting the protein droplet in the experimental chamber for crystallization. Please click here to view a larger version of this figure.

Figure 9: Recorded photograph for THM_3 Macro-crystals showing various sizes of thaumatin crystals in solution. The picture was taken 20 h after setting the protein droplet in the experimental chamber for crystallization. Please click here to view a larger version of this figure.
| Substance | Molarity/Percentage | Time (s) |
| water | 0 | 100 |
| water | -25 | 2100 |
| water | 0 | 2100 |
Table 1: Automated schedule input for the sample evaporation step in the production of thaumatin microcrystals.
| Substance | Molarity/Percentage | Time (s) |
| water | 0 | 100 |
| prec | 0.8 | 1800 |
| water | 0 | 18000 |
Table 2: Automated schedule input for the precipitant addition step in the production of thaumatin microcrystals.
