

Summary
微生物基因组测序过程中的污染仍然是一个大问题。在这里, 我们展示了一种方法来序列的缓步动物的基因组从一个单一的样本, 只有 50 pg 的基因组 DNA 没有整个基因组的放大, 以减少污染的风险。
Abstract
水熊蟲是微小的动物, 进入一个 ametabolic 的状态称为 anhydrobiosis 时, 面对干燥, 可以返回到他们的原始状态, 当水供应。像水熊蟲这样的微小动物的基因组测序有可能导致细菌污染, 例如, 关于这些动物水平基因转移的程度。在这里, 我们提供了一个超低的输入方法序列的基因组的缓步动物, Hypsibius dujardini, 从一个单一的标本。通过使用严格的洗涤和污染物排除以及高效提取单个个体的 50 ~ 200 pg 基因组 DNA, 我们构建了一个用 DNA 测序仪测序的库。这些图书馆是高度重现性和无偏见的, 并对序列读取与其他H. dujardini基因组的信息学分析显示了少量的污染。此方法可应用于无法用以前的方法排序的 unculturable 水熊蟲。
Introduction
水熊蟲是微小的动物, 可以进入一个 ametabolic 的状态称为 anhydrobiosis 时, 面对干燥。他们由水的吸收1,2恢复。在 ametabolic 状态下, 水熊蟲能够耐受各种极端环境, 其中包括极端温度3和压力4,5, 高剂量的紫外线光6, X 射线和伽玛射线7,8, 宇宙空间9。基因组数据是研究 anhydrobiosis 分子机制的不可缺少的基础。
以前试图序列的水熊蟲基因组已经显示细菌污染的迹象10,11,12,13,14。基因组测序从这些小有机体需要大量的动物和容易细菌污染;因此, 我们以前已经建立了一个排序协议使用一个超低的输入方法从一个单一的缓步动物样本, 以最小化15的风险。利用这些数据, 我们进一步进行了高品质的重测序和重组dujardini16,17的基因组。在这里, 我们详细描述了这个方法的基因组测序从一个单一的缓步动物个体(图 1)。这种测序方法的验证超出了本文的重点, 在我们上一份报告16中已经作了深入的讨论。
该方法由两部分组成: 分离出具有最低污染的单一缓步动物, 以及高质量的象形文字 DNA 水平提取。缓步动物是饥饿和冲洗彻底与水, 以及抗生素, 并观察下显微镜下500X 倍放大, 以确保清除任何细菌污染。以前的估计和测量表明, 单个的缓步动物包含大约 50-200 pg 的基因组 DNA16, 这是通过裂解甲壳素外骨骼的冻融循环或手工均匀化提取。这种基因组 dna 被提交到图书馆建设和排序的 dna 测序仪。另外一项信息学分析显示, 与以前的缓步动物测序项目相比, 高质量的测序以及较低的污染水平。
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. 准备
- 用蒸馏水 (DW) 制备2% 琼脂糖凝胶, 作为90毫米塑料培养皿中的溶剂, 10 毫升1% 青霉素/链霉素鸡尾酒与 DW。凝胶可以储存 2-3 周的孵化器设置在18摄氏度。
注: 避免任何贮存的凝胶低于10摄氏度, 因为低温会收缩琼脂糖凝胶, 造成一个微小的缝隙之间的凝胶和培养皿壁, 其中水熊蟲可以被困。
2. 样品制备和污染物排除
- 收集一个单一的缓步动物, 把它放在准备好的琼脂板上, 并用 DW 清洗 2x-3x, 以除去剩余的粒子。
- 孵化的缓步动物在 18-22 °c 24 小时, 以消除任何多余的食物从肠道。
- 将饥饿的缓步动物在青霉素/链霉素抗生素中放置 2-6 小时, 去除任何细菌污染, 并使用 P10 吸管将被净化的动物放在干净的滑动玻璃上。
- 在显微镜下观察缓步动物500X 放大倍数, 并确认没有残留细菌。
注: 显微镜的高放大倍数是最佳的, 但光学显微镜可以在不应用盖玻片的情况下交替使用。 - 收集的人使用的 P10 吸管最多5µL 的液体, 将其放入低结合聚合酶链反应 (PCR) 管, 并尽可能多地去除多余的液体。
注意: 管内, PCR 管和吸管提示应该是低约束力, 以减少 DNA 的损失。
注意: 尽快进行同质化, 因为动物的快速脱水或死亡会严重损害基因组 DNA。
3. 均匀化和 DNA 提取
- 融汇动物以下列方法之一获得其基因组 DNA (gDNA)。
注意: 在下面的 DNA 提取步骤中, 使用材料表中所示的指定套件是非常关键的, 因为其他套件 (无论是柱形还是珠基) 在这个极低的输入中是不有效的。在使用前, 基因组裂解缓冲液已经提供了0.5% β巯基乙醇。- 融汇的动物与冻融循环18。
- 紧接着步骤 2.5, 添加100µL 的裂解缓冲液的 PCR 管含有缓步动物。
- 将 PCR 管放置在液氮中10分钟, 10 分钟, 将其移动到加热到37摄氏度的热块中。重复此步骤2x。
注: 本步骤可在均匀化不足或手动粉碎后进行重复。
- 手动粉碎动物。
- 紧接着步骤 2.5, 在显微镜下, 用吸管的 P10 尖端挤压该动物, 并将其压在 PCR 管壁上, 然后立即添加100µL 的溶解缓冲液。
注意: 在显微镜下观察这个过程是非常重要的, 因为缓步动物很容易溜掉, 而且低效的粉碎会导致无法提取 gDNA。确保缓步动物角质层断裂, 使裂解缓冲液渗入机体。
- 紧接着步骤 2.5, 在显微镜下, 用吸管的 P10 尖端挤压该动物, 并将其压在 PCR 管壁上, 然后立即添加100µL 的溶解缓冲液。
- 融汇的动物与冻融循环18。
- 在室温下孵化管30分钟以进行裂解。
注: 有效裂解至少需要30分钟, 但孵化时间可能更长。 - 将裂解混合物的完整容积 (100 µL) 转移到干净的1.5 毫升低结合微细。
- 将100µL 溶解缓冲剂添加到用于均匀化的低结合 PCR 管中, 现在是空的, 在吹打混合物后, 将其转移到步骤3.3 中使用的1.5 毫升低约束微细。重复此步骤2x。
- 与步骤3.4 中一样, 在低结合 PCR 管中加入300µL 裂解缓冲液, 并在吹打后将混合物移动到1.5 毫升低束缚微细。1.5 毫升低结合微细应包含600µL 的混合物后, 这一步。
注意: 这些步骤是为了尽量减少 gDNA 的损失, 通过多次洗涤样品来降低链到 PCR 管壁的损耗。 - 将600µL 的裂解混合物添加到放置在收集管中的旋柱上, 并将其离心在 1万 x g 处, 1 分钟。
- 将流量重新应用到柱上, 并将其离心在 1万 x g 处1分钟。
注意: 此步骤对于确保大多数 gDNA 绑定到该列至关重要。 - 将500µL 的洗涤缓冲器添加到旋转柱上, 并将其离心在 1万 x g 上1分钟. 将旋转柱转移到干净的 1.5 mL 微细。
- 应用20µL 10 毫米三盐酸, pH 8.5, 到自旋柱, 在室温下等待5分钟, 并离心它在 1万 x g 为1分钟。
注: 洗脱缓冲液不能含有 EDTA, 因为它干扰了库的制备酶。 - 重新应用流通过自旋柱, 并在室温孵化5分钟后, 离心机1分钟 1万 x g。
注意: 此步骤对于确保绑定到列的 gDNA 的最大洗脱至关重要。
4. 图书馆建设顺序
-
DNA 碎片
- 将 gDNA 淋洗剂组成的15µL 转移到15µL 微细, 用于 DNA 碎片化, 并使用桌面离心将管离心为1分钟。
注意: 在材料表中显示的指定微细对低输入是最佳的。低体积超声波的碎片是至关重要的, 由于 DNA 浓度极低, 这不能代替酶分裂。 - 碎片 gDNA 到 550 bp。
注: 我们使用的 550 bp 设置如下: 峰值事件功率 = 30 W, 责任因数 = 20%, 周期每突发 = 50, 治疗时间 = 二十三年代。 - 经过彻底的吹打, 转移10µL 的碎片 DNA 混合物到一个干净的低结合 PCR 管。
注意: 实验可以在这里停止。保存在4摄氏度或-20 摄氏度的 DNA。
- 将 gDNA 淋洗剂组成的15µL 转移到15µL 微细, 用于 DNA 碎片化, 并使用桌面离心将管离心为1分钟。
-
图书馆建设顺序
注: 由于输入 DNA 量低, 在下列过程中使用所述材料表中的指定套件是绝对关键的。- 添加2µL 的模板准备缓冲区和1µL 的模板制备酶, 并将其与吸管彻底混合。
- 在热循环仪上执行模板准备反应, 条件如下:22 °c 为25分钟, 55 °c 20 分钟, 4 °c 举行, 和一个加热盖子在 101-105 °c。一旦反应完成后, 继续下一步。
- 添加1µL 的图书馆合成缓冲器和1µL 的图书馆合成酶的模板制备反应产品和孵化混合物在22°c 40 分钟 (与4°c 举行)。一旦反应完成后, 继续下一步。
- 增加30µL 的图书馆放大主混合 (25 µL 的图书馆放大缓冲器, 1 µL 的图书馆扩增酶, 4 µL 核酸酶水) 和5µL 的索引试剂。
- 用吸管将所有东西彻底混合, 然后用台式离心机将混合物简单地离心。
- 执行 PCR 与表 1所列条件。
| 温度 | 时间 | 周期 |
| 72˚C | 3分钟 | |
| 85˚C | 2分钟 | |
| 98˚C | 2分钟 | |
| 98˚C | 20秒 | 4个周期 |
| 67˚C | 20秒 | |
| 72˚C | 40秒 | |
| 98˚C | 20秒 | 16个周期 |
| 72˚C | 50秒 | |
| 4˚C | 举行 |
表 1: PCR 条件。
-
PCR 产品的纯化
注: 此步骤可替代其他纯化方法。如果使用另一种方法, 请确保使用分光光度计检查生成的 DNA 纯度。吸光度比为260/280 和260/230 必须在1.8 以上。- 添加50µL 磁珠, 吸管的解决方案 10x, 并离心机它与桌面离心。
- 在室温下孵化溶液2分钟。
- 在磁性支架上孵育5分钟, 或直到溶液完全清除, 然后取出上清。
- 添加200µL 新鲜制备的80% 乙醇的低结合 PCR 管在磁铁立场, 等待三十年代, 并删除上清。重复此步骤2x。不要打扰珠子。
- 简单地将低结合 PCR 管与台式离心机分离, 并去除磁性支架上多余的乙醇。空气干燥的珠子, 但避免 overdrying。
- 并用重悬15µL 10 毫米三盐酸, pH 8.5, 彻底吸管溶液, 使磁性珠均匀分布, 在室温下孵化管2分钟, 并在短暂离心后, 孵化在一个磁性立场2分钟, 直到解决方案是明确的。
注: 洗脱缓冲器不应含有 EDTA, 因为它干扰测序化学。 - 转移上清, 不干扰颗粒, 到一个新的低结合 PCR 管。
注意: 实验可以在这里停止。保存的 DNA 在4摄氏度或在-20 摄氏度长时间储存。
5. DNA 的质量检查、定量和序列
注: 由于 DNA 含量低, 本步骤前未进行质量检查。
-
DNA 库大小分布的验证
注: 其他高灵敏度的电泳系统可以使用数字报告的大小分布。- 返回电泳缓冲试剂和凝胶盒 (25-1000 bp 在范围内) 到室温。
- 添加3µL 电泳缓冲试剂与1µL 的测序库, 并将其彻底混合1分钟与涡, 并简要离心他们与台式离心机。
- 与相关软件进行电泳和验证库大小分布。主要的碎片峰值应该从大约300到 1000 bp 不等。
-
DNA 量化
注: 其他基于荧光的方法或定量 PCR 也可以使用, 但应避免分光光度法, 因为它是不够准确, 以量化测序库。- 添加796µL 溶液缓冲器和4µL 荧光试剂, 并将其彻底混合。将190µL 的工作溶液分配给两个化验管和197µL 到一个化验管。
- 将已知的 DNA 浓度的10µL 标准添加到每个检测管中, 其中包含190µL 的工作溶液, 3 µL 准备好的库到含有197µL 工作溶液的检测管。
- 将管子简单地涡旋, 然后将其离心在台式离心机上。使用具有3µL 设置的荧光计量化 DNA。
-
DNA 库序列
注意: 测序平台必须与指定的库结构套件兼容。- 根据制造商的协议准备排序库。
- 将试剂盒和流动单元设置为测序仪, 并按照制造商的协议输入顺序运行信息。
- 运行排序。
注: 我们已经进行了两个排序运行: 一个样本/运行, 以及四样本复用一次运行。
6. 计算分析
- 基调用, 如果需要, 解读取。
- 使用 FastQC19验证序列数据的质量。
注意: 为了更深入地验证获得的数据, 请参阅我们以前的报告16。
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
污染物排除:
该协议包括彻底清洗缓步动物和抗生素治疗灭菌, 以减少污染。它还包括一个视觉检查过程, 以确保这些过程的完整性。在验证过程 (协议步骤 2.4) 中所做的显微镜图像如图 2所示。当观察到500X 放大, 细菌细胞可以被视为小颗粒移动周围的缓步动物个人。
DNA 库质量验证:
所构建的 DNA 序列库总量约为 109.5 ng (7.3 µL x 15 µL)16。为了验证碎片的长度分布, 电泳模式应类似于图 3.当我们将碎片大小设置为 550 bp 的 DNA 剪切系统时, 该库应该是 550-600 bp, 包括测序适配器。可以观察到, 序列库的大多数包含在 200-1000 bp 之间, 并且在复制 (N1-N4) 之间是一致的。
序列数据分析:
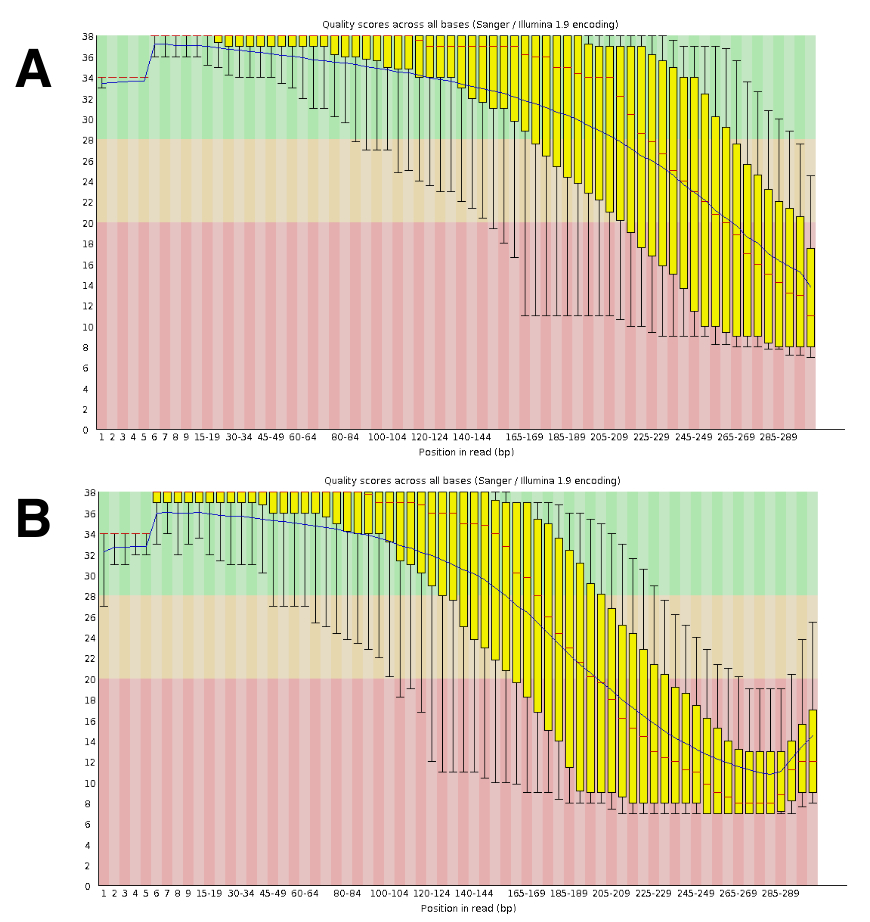
DNA 测序产生了大约20到25米配对读取每运行。质量的验证是使用 FastQC 进行的 (图 4)。沿顺序读取的质量分布是 300 bp 配对运行的典型特征。

图 1: 此协议的工作流。此图显示了该协议的摘要。请单击此处查看此图的较大版本.

图 2: 无菌缓步动物的代表性照片.该图显示了被污染的 (左) 缓步动物和清洁 (右) 缓步动物 (Hypsibius dujardini) 的图像以及进一步放大的图像 (底部)。缓步动物周围的棒状细胞是污染物, 用箭头表示。刻度条指示100µm.请点击这里查看这个数字的大版本。

图 3: 验证构造 DNA 库的片段长度分布.此面板显示排序库大小的分布。紫色和绿色线条分别表示1500和 25 bp 的上、下标记。L = 梯子, S = 1 样品/奔跑, N1-N4 = 4 复制或奔跑。请单击此处查看此图的较大版本.

图 4: 用 FastQC 验证 DNA 序列质量的例子.DNA 序列数据被提交到 FastQC, 以验证顺序性能。显示每个基序列质量的 DRR055040 的代表结果 (DDBJ 序列读存档 DRA00445516)。(A) 本小组显示远期读数 (R1)。(B) 本小组显示反向读数 (R2)。请单击此处查看此图的较大版本.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
细菌污染对微生物的基因组测序构成威胁。虽然以前关于缓步动物基因组测序的研究已经通过广泛的信息学方法12、20来筛选出污染, 但我们对单个个体的基因组进行了排序, 以尽量减少污染物的风险。由于单个缓步动物包含大约 50-200 pg 基因组 DNA16和包裹在厚层甲壳素外骨骼, 排除污染物和高质量的 DNA 提取是本议定书的关键点。现有的水熊蟲文化不是无菌的, 而从野外收集的细菌在表面上携带大量的污染物, 以及肠道内的食物残留。水熊蟲以前的基因组测序项目已将 1万-10万人统称为一个样本12,14, 这意味着结果很可能受到细菌污染的影响。在他们的报告中, 布思比等。通过使用他们的阴性趋光性行为, 收集了dujardini的个体14, 并且小组没有使用任何抗菌方法。
为了直观地检查是否有污染物, 我们在抗生素 (青霉素/链霉素) 中孵化缓步动物, 并在500X 显微镜下对个体进行检查。通过隔离单个个体, 并仔细检查它的任何污染物, 我们尽量减少可能的污染物的风险。测序数据也证实了低污染水平,16。至于 DNA 提取, 我们采用了人工均匀化, 以及热均匀化18。通过将缓步动物个体提交到液氮和37°c, 在甲壳素外骨骼中诱导裂纹, 裂解缓冲液能够穿透体内, 溶解细胞。当 DNA 产量低于预期时, 可以进行热和人工均质, 以最大限度地提高产量。
本文所述的方法有若干限制。首先, 从线虫的研究中应用冻融循环的均匀化;因此, 该方法可能只对 ecdysozoa 有效。其次, 由于 dna 序列库阶段 dna 片段的放大, PCR 误差的可能性不容忽视。因此, 对于需要高精度读数 (即 SNP 分析) 的分析, 不建议使用序列数据。此外, 正如我们在《议定书》中所指出的那样, 由于输入 DNA 量很低, 在材料表中显示的指定 SNA-序列套件的使用是绝对关键的。此 DNA 库构造套件 ligates Illumina 适配器序列在放大之前;因此, 使用 PacBio 或纳米技术, 无法将此库应用于长读排序。最后, 本协议中构建的 DNA 库的质量检查只有一次, 在测序库构造之后才发生。这是由于 dna 的低输入, 因为大多数 dna 定量和电泳方法无法检测到 50-200 pg 的 dna。因此, 我们进行了质量检查, 如电泳 (图 1) 和荧光 quantifications, 只有在 PCR 扩增后。
对这些数据的生物信息学分析的充分讨论超出了本文的范围;不过, 我们已简略说明我们进行了几项分析。用 FastQC19对测序数据进行质量检查, 计算每基质量、序列复制等. 已验证的序列数据可以提交到基因组组件。我们已经组装了一个 132 Mb 的基因组与 MaSuRCA v3.1.321 , 并已比较的映射统计数字与 BWA22和 QualiMap23的 DNA 测序库与其他H. dujardini基因组汇编16。此外, 我们还使用了这一 DNA 测序数据, 以排除污染物在我们的研究17, 并指出, 测序读数均匀分布在整个基因组。
大多数非模型生物的项目从培养足够的样本材料开始, 就像水熊蟲24的情况一样。技术进步的文化技术已经使大量的缓步动物文化, 但目前的文化方法还没有无菌性, 因为大多数水熊蟲仍然 unculturable 在实验室里, 几乎不可能进行基因组或转录测序。这一单一个体的 DNA 测序方法使得分析稀有的缓步动物物种成为可能, 其中包括研究较少的海洋物种。通过在更广泛的种类地区进行比较基因组学, 可以进一步了解水熊蟲的 anhydrobiosis 机制。
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
作者没有什么可透露的。
Acknowledgments
作者感谢 Nozomi、Takai 和 Nahoko 在基因组测序中的技术支持。这项工作得到了日本促进科学学会 (jsp) 研究员的资助, KAKENHI 为年轻科学家提供援助 (22681029 号), 以及 KAKENHI 的科研资助 (B), 来自 jsp 的17H03620 号, 由一个来自住友基金会 (140340 号) 基础科学研究项目的赠款, 部分是由山形县政府和日本鹤的研究基金提供的。以小球藻为原料, 提供水熊蟲的小球藻。
Materials
| Name | Company | Catalog Number | Comments |
| SZ61 microscope | OLYMPUS | ||
| BactoAgar | Difco Laboratories | 214010 | |
| Penicillin Streptomycin (10,000 U/mL) | Gibco by life technologies | 15140-148 | |
| VHX-5000 System | Keyence | ||
| 0.2mL Silicone coating tube | Bio Medical Science | BC-bmb20200 | |
| Quick-DNA Microprep Kit | ZYMO Research | D3021 | Use of this kit is absolutey critical; see step 3.1 |
| 1.5 mL microtube | greiner bio-one | 616-201 | See 4.1.1 |
| HIgh speed refrigerated micro centrifuge | TOMY | MX-307 | |
| Covaris M220 | Covaris Inc. | 4482277 | |
| ThruPLEX DNA-Seq kit | Rubicon Genomics | CAT. NO. R400406 | Use of this kit is absolutey critical; see step 4.2 |
| Thermal Cycler | Bioer Technology | TC-96GHbC | |
| AMPure XP reagent | BECKMAN COULTER Life Science | A63881 | |
| Ethanol | Wako | 054-027335 | |
| EB buffer | QIAGEN | 19086 | |
| 2200 TapeStation | Agilent | G2965AA | |
| D1000 Reagents | Agilent | 5067-5583 | |
| D1000 ScreenTape | Agilent | 5067-5582 | |
| Qubit dsDNA BR Buffer/Reagent | ThermoFisher Scientific | Q32850 | |
| Cubee Mini-Centrifuge | RecenttecGenereach | R5-AQBD01aqbd | |
| MiSeq 600 cycle v3 | Illumina Inc. | MS-102-3003 | |
| MiSeq Sequencer | Illumina Inc. | SY-410-1003 |
References
- Crowe, J. H., Hoekstra, F. A., Crowe, L. M. Anhydrobiosis. Annual Review of Physiology. 54 (1), 579-599 (1992).
- Mobjerg, N., et al. Survival in extreme environments - on the current knowledge of adaptations in tardigrades. Acta Physiologica. 202 (3), 409-420 (2011).
- Becquerel, P. La suspension de la vieau dessous de 1/20 K absolu par demagnetization adiabatique de L'alun de fer dans le vide les plus eléve. Comptes Rendus de l'Académie des Sciences. 231, 261-264 (1950).
- Ono, F., et al. Effect of ultra-high pressure on small animals, tardigrades and Artemia. Cogent Physics. 3 (1), 1167575 (2016).
- Horikawa, D. D., et al. Tolerance of anhydrobiotic eggs of the Tardigrade Ramazzottius varieornatus to extreme environments. Astrobiology. 12 (4), 283-289 (2012).
- Horikawa, D. D., et al. Analysis of DNA repair and protection in the Tardigrade Ramazzottius varieornatus and Hypsibius dujardini after exposure to UVC radiation. PLoS One. 8 (6), e64793 (2013).
- Horikawa, D. D., et al. Radiation tolerance in the tardigrade Milnesium tardigradum. International Journal of Radiation Biology. 82 (12), 843-848 (2006).
- May, R. M., Maria, M., Gumard, J. Action différentielle des rayons x et ultraviolets sur le tardigrade Macrobiotus areolatus, a L'état actif et desséché. Bulletin Biologique de la France et de la Belgique. 98, 349-367 (1964).
- Jonsson, K. I., Harms-Ringdahl, M., Torudd, J. Radiation tolerance in the eutardigrade Richtersius coronifer. International Journal of Radiation Biology. 81 (9), 649-656 (2005).
- Bemm, F., Weiss, C. L., Schultz, J., Forster, F. Genome of a tardigrade: Horizontal gene transfer or bacterial contamination? Proceedings of the National Academy of Sciences of the United States of America. 113 (22), E3054-E3056 (2016).
- Delmont, T. O., Eren, A. M. Identifying contamination with advanced visualization and analysis practices: metagenomic approaches for eukaryotic genome assemblies. PeerJ. 4, e1839 (2016).
- Koutsovoulos, G., et al. No evidence for extensive horizontal gene transfer in the genome of the tardigrade Hypsibius dujardini. Proceedings of the National Academy of Sciences of the United States of America. 113 (18), 5053-5058 (2016).
- Boothby, T. C., Goldstein, B., et al. Reply to Bemm et al. and Arakawa: Identifying foreign genes in independent Hypsibius dujardini genome assemblies. Proceedings of the National Academy of Sciences of the United States of America. 113 (22), E3058-E3061 (2016).
- Boothby, T. C., et al. Evidence for extensive horizontal gene transfer from the draft genome of a tardigrade. Proceedings of the National Academy of Sciences of the United States of America. 112 (52), 15976-15981 (2015).
- Arakawa, K. No evidence for extensive horizontal gene transfer from the draft genome of a tardigrade. Proceedings of the National Academy of Sciences of the United States of America. 113 (22), E3057 (2016).
- Arakawa, K., Yoshida, Y., Tomita, M. Genome sequencing of a single tardigrade Hypsibius dujardini individual. Scientific Data. 3, 160063 (2016).
- Yoshida, Y., et al. Comparative genomics of the tardigrades Hypsibius dujardini and Ramazzottius varieornatus. PLoS Biology. 15 (7), e2002266 (2017).
- He, F. Total RNA Extraction from C. elegans. Bio-protocol. Bio101, e47 (2011).
- Andrews, S. FastQC a quality-control tool for high-throughput sequence data. , http://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (2015).
- Hashimoto, T., et al. Extremotolerant tardigrade genome and improved radiotolerance of human cultured cells by tardigrade-unique protein. Nature Communications. 7, 12808 (2016).
- Zimin, A. V., et al. The MaSuRCA genome assembler. Bioinformatics. 29 (21), 2669-2677 (2013).
- Li, H., Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics. 25 (14), 1754-1760 (2009).
- Okonechnikov, K., Conesa, A., Garcia-Alcalde, F. Qualimap 2: advanced multi-sample quality control for high-throughput sequencing data. Bioinformatics. 32 (2), 292-294 (2016).
- Horikawa, D. D., et al. Establishment of a rearing system of the extremotolerant tardigrade Ramazzottius varieornatus: a new model animal for astrobiology. Astrobiology. 8 (3), 549-556 (2008).

{kind=link}