


Summary
Apresentamos uma metodologia para estabelecer os requisitos de polinização das cultivares de damasco(Prunus armeniaca L.) combinando a determinação da auto-(in)compatibilidade por microscopia de fluorescência com a identificação do S-genótipo pela análise pcr.
Abstract
A autoincompatibilidade em Rosaceae é determinada por um Sistema de Autoincompatibilidade Gametofitic (GSI) que é controlado principalmente pelo lócus multiloelico S. No damasco, a determinação das relações de auto e inter-(in)compatibilidade é cada vez mais importante, uma vez que a liberação de um importante número de novas cultivares resultou no aumento de cultivares com requisitos desconhecidos de polinização. Aqui, descrevemos uma metodologia que combina a determinação da auto-(in)compatibilidade por polinizações manuais e microscopia com a identificação do genótipo Spela análise pcr. Para a determinação de auto-(in)compatibilidade, foram coletadas flores em fase de balão de cada cultivar no campo, polinizado à mão em laboratório, fixado e manchado com azul anilina para observação do comportamento do tubo de pólen sob a microscopia de fluorescência. Para o estabelecimento de relações de incompatibilidade entre cultivares, o DNA Sde cada cultivar foi extraído de folhas jovens e S-alelos foram identificados pela PCR. Essa abordagem permite estabelecer grupos de incompatibilidade e elucidar relações de incompatibilidade entre cultivares, o que fornece informações valiosas para escolher polinizadores adequados na concepção de novos pomares e selecionar pais apropriados em programas de reprodução.
Introduction
A autoincompatibilidade é uma estratégia de floração das plantas para prevenir a auto polinização e promover o outcrossing1. Em Rosaceae, esse mecanismo é determinado por um Sistema de Autoincompatibilidade Gametofitic (GSI) que é controlado principalmente pelo lócus multitélico S2. No estilo, o gene RNase codifica o determinante S-stylar, um RNase3, enquanto uma proteína F-box, que determina o determinante do pólen S,é codificada pelo gene SFB 4. A interação de autoincompatibilidade ocorre através da inibição do crescimento do tubo de pólen ao longo do estilo que impede a fertilização do ovule5,6.
No damasco, uma renovação varietal ocorreu em todo o mundo nas últimas duas décadas7,8. Essa introdução de um importante número de novas cultivares, de diferentes programas de reprodução pública e privada, resultou no aumento de cultivares de damasco com requisitos desconhecidos de polinização8.
Diferentes metodologias têm sido usadas para determinar os requisitos de polinização em damasco. No campo, a auto-compatibilidade pode ser estabelecida por polinização controlada em árvores enjauladas ou em flores emasculadas e,posteriormente,registrando o percentual de conjunto de frutas9,10,,11,12. Além disso, polinizações controladas têm sido realizadas em laboratório pela cultura semi-in vivo de flores e análise do comportamento do tubo de pólen sob microscopia de fluorescência8,,13,,14,,15,,16,17. Recentemente, técnicas moleculares, como análise e sequenciamento de PCR, permitiram a caracterização de relações de incompatibilidade com base no estudo dos genes RNase e SFB 18,19. No damasco, foram relatados 33 S-alelos(S1 a S20, S22 a S30, S52, S53, Sv, Sx), incluindo um alelo relacionado com auto-compatibilidade(Sc)12, 18,,18,20,,21,,22,,23,,24. Até agora, 26 grupos de incompatibilidade foram estabelecidos nesta espécie de acordo com o S-genótipo8,9,17,25,26,27. Cultivares com os mesmos alelos Ssão intercompatíveis, enquanto cultivares com pelo menos um s-alelodiferente e, consequentemente, alocados em diferentes grupos incompatíveis, são intercompatíveis.
Para definir os requisitos de polinização das cultivares de damasco, descrevemos uma metodologia que combina a determinação da auto-(in)compatibilidade por microscopia de fluorescência com a identificação do genótipo Spela análise pcr em cultivares de damasco. Essa abordagem permite estabelecer grupos de incompatibilidade e elucidar relações de incompatibilidade entre cultivares.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. Determinação de auto-(in)compatibilidade
- Prove as flores no campo. É necessário coletar as flores no estágio do balão(Figura 1A),correspondente ao estágio 58 na escala BBCH para damasco28, para evitar polinização prévia indesejada.
-
Polinizações auto e cruzadas em laboratório
- Retire os anteratos das flores no estágio do balão e coloque-os em um pedaço de papel para secar à temperatura do laboratório.
- Após 24h, peneirar os grãos de pólen utilizando uma malha fina (0,26 mm)(Figura 1B).
- Emascular um grupo de 30 flores no mesmo estágio de balão para cada auto-polinização e polinização cruzada e colocar os pistils na espuma florista na água em temperatura laboratorial(Figura 1C).
- Polinize as pistils com a ajuda de um pincel com pólen de flores da mesma cultivar 24 h após a emasculação. Além disso, poliniza outro conjunto de pistils de cada cultivar com pólen a partir de flores de um polinizador compatível como controle(Figura 1D).
- Após 72 h, fixar os pistils em uma solução fixativa de etanol/ácido acético (3:1) por pelo menos 24 h a 4 °C29. Em seguida, descarte o fixador e adicione 75% de etanol garantindo que as amostras estejam completamente submersas na solução. As amostras podem ser conservadas nesta solução a 4 °C até o uso8,17,30,31,32.
-
Avaliação da viabilidade do pólen através da germinação in vitro de pólen
- Para preparar o meio de germinação, peso 25 g de sacarose, 0,075 g de ácido bórico (H3BO3) e 0,075 g de nitrato de cálcio (Ca(NO3)2)33.
- Adicione os componentes do meio em 250 mL de água destilada e dissolva completamente.
- Solidifique o meio adicionando 2 g de agarose e misture girando.
- Verifique o pH do meio usando um medidor de pH e ajuste o valor para 7.0 com solução NaOH ou HCl.
- Autoclave a mistura para esterilizar o meio.
- Depois de autoclaving, esfrie o meio e distribua-o em pratos petri em um capô de fluxo laminar estéril.
- Espalhe os grãos de pólen das mesmas cultivares utilizadas para as polinizações controladas no meio de germinação solidificada do pólen e observe-os sob o microscópio após 24 horas6.
NOTA: Para esterilizar a coifa de fluxo laminar, limpe a superfície com 70% de etanol e ligue a lâmpada UV durante 10 minutos. - Guarde as placas de Petri em uma geladeira a 4 °C até usar.
-
Observações de microscopia
- Lave os pistils três vezes por 1h com água destilada e deixe-os em 5% de sulfito de sódio a 4 °C. Após 24 h, autoclave-os a 1 kg/cm2 durante 10 min em sulfito de sódio para suavizar os tecidos34.
- Coloque os pistils autoclavados sobre um escorregador de vidro e, com a ajuda de um bisturi, remova as casas trica ao redor do ovário para obter uma melhor visualização dos tubos de pólen. Em seguida, esmague os pistils com um copo de cobertura.
- Prepare 0,1% (v/v) mancha azul anilina: misture 0,1 mL de azul anilina em 100 mL de 0,1 N linfóteo de potássio tribasic (K3PO4). Aplique uma gota de azul anilina sobre os preparativos para manchar os depoimentos de callose durante o crescimento do tubo de pólen.
- Observe os tubos de pólen ao longo do estilo por um microscópio com epifluorescência UV usando 340-380 bandpass e 425 filtros de longopass.
2. Extração de DNA
- Amostra 2-3 folhas no campo na primavera. Recomenda-se a amostra das folhas em estágios jovens, uma vez que o DNA obtido é de maior qualidade e níveis mais baixos de compostos fenólicos em comparação com as folhas antigas.
- Extrair DNA genômico seguindo as etapas descritas em um kit comercialmente disponível (ver Tabela de Materiais).
- Analise a quantidade e a qualidade das concentrações de DNA utilizando espectrofotômetro UV-vis (260 nm).
3. Identificação de alelo
-
Configuração das reações do PCR
- Prepare uma diluição de 50 ng/μL em água destilada de cada amostra de extração de DNA.
- Descongele os reagentes PCR lentamente e mantenha-os no gelo. Deixe o DNA polimerase no congelador até que seja necessário.
- Prepare as reações de amplificação usando as diferentes combinações de primers. Crie o mix de reação pcr combinando os componentes na Tabela 1. Vortex a reação PCR mistura bem e distribui o volume indicado para as diferentes combinações de primers para cada poço da placa PCR. Em seguida, adicione 1 μL da diluição de DNA em cada poço.
- Coloque a placa PCR no termociclador e execute o programa PCR correspondente mostrado na Tabela 1.
- Analise os fragmentos amplificados. Há principalmente duas maneiras diferentes de analisar os fragmentos amplificados do PCR: eletroforese capilar (CE) com primers fluorescentes rotulados ou como amplificadores visuais de eletroforese de gel agarose com primers não rotulados.
-
Eletroforese capilar
- Para preparar o tampão de carga, misture 35 μL de formamide deionizado com 0,45 μL de padrão de dimensionamento rotulado. Vórtice o reagente para misturar bem, e depois dispense 35,5 μL no poço da placa do leitor.
- Adicione 1 μL do produto PCR no poço. Além disso, adicione uma gota de óleo mineral para evitar a evaporação da água.
- Prepare a placa de separação adicionando tampão de separação.
- Use o software comercial incluído com o analisador genético (ver Tabela de Materiais). Crie uma nova placa de amostra e guarde os nomes da amostra para todos os poços na placa.
- Selecione o método de análise. Neste caso, desnaturar as amostras a 90 °C para 120 s, injetar a 2,0 kV para 30 s e separar a 6,0 kV por 35 min.
- Insira as duas placas no analisador genético. Encha a matriz capilar com água destilada.
- Carregue o gel de poliacrilamida linear patenteado (LPA). Finalmente, clique em Executar.
-
Eletroforese de gel
- Prepare um gel de 1% de agarose adicionando 1,5 g de agarose grau de biologia molecular em 150 mL de 1x TAE (Tris-acetato-EDTA) tampão de execução (40 mM Tris, ácido acético de 20 mM e 1 mM EDTA no pH 8.0). Dissolva a ágarose por aquecimento de micro-ondas por 2-3 min.
- Para visualizar o DNA, adicione 4 μL de uma mancha de ácido nucleico (ver Tabela de Materiais) e misture suavemente.
- Adicione um pente de gel, com poços suficientes para escadas, controles e amostras, em uma bandeja de gel. Em seguida, despeje lentamente a mistura no meio da bandeja de gel e evite bolhas.
- Deixe o gel esfriar por 30-45 min em temperatura ambiente até que o gel se solidifique completamente. Introduza o gel na câmara de eletroforese, remova o pente de gel e encha a câmara com tampão TAE suficiente para cobrir o gel.
NOTA: Verifique a colocação do gel. Os poços devem ser colocados perto do polo negativo, uma vez que o DNA carregado negativamente migra para o cátodo. - Adicione 5 μL de tampão de carga (0,1% (v/v) azul bromofenol) aos produtos PCR e misture bem.
- Para estimar o tamanho das bandas, carregue 5 μL de escada de peso molecular de DNA (ver Tabela de Materiais).
- Coloque as amostras nos poços adicionais do gel.
- Uma vez que todas as amostras e a escada de peso molecular de DNA são carregadas, execute o gel a 90 V por 1-1,5 h, até que a linha de corante azul esteja aproximadamente a 75% do comprimento do gel.
- Visualize as bandas em um transilluminador para ácidos nucleicos.
-
Eletroforese capilar
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Estudos de polinização em damasco exigem o uso de flores na fase final do balão um dia antes da anestesia (Figura 1A). Esta etapa é considerada a mais favorável tanto para a coleta de pólen quanto para pistil, uma vez que as estruturas florais estão quase maduras, mas a deshiscência anther ainda não ocorreu. Isso evita a interferência de pólen indesejado, não só de pólen da mesma flor, mas também de outras flores, uma vez que as pétalas fechadas impedem a chegada de insetos portadores de pólen externo. Os grãos de pólen são facilmente peneirados através de uma malha fina(Figura 1B) de anteras deshisced previamente colocados em um pedaço de papel por 24 horas à temperatura ambiente ou com leve calor extra. Da mesma forma, os pistils são obtidos a partir de flores no estágio de balão após a remoção de pétalas, sepalas e estamens com a ajuda de pinças ou unhas(Figura 1C). Pistils podem ser auto-polinizados com uma escova fina(Figura 1D).
As flores hermafrodroditas de damasco têm cinco sepalas vermelhas escuras, cinco pétalas brancas (Figura 1A),um único pistil(Figura 2A) e 25-30 estamens. O pistil tem três estruturas principais: estigma, estilo e ovário. O ovário tem dois óvulos, e a fertilização de pelo menos um deles é necessária para a criação de frutas. Durante a polinização, os insetos, principalmente as abelhas, transferem grãos de pólen para o estigma (Figura 1A), onde germinam(Figura 2B) dentro de 24h após a polinização. Um tubo de pólen é produzido a partir de cada grão de pólen germinatório, que cresce através das estruturas pistil para chegar ao ovário após 3-4 dias e fertilizar um dos dois ovulos após cerca de 7 dias. Em cultivares auto-incompatíveis em que o alelo S do grão de pólen é o mesmo que um dos dois pistils, o tubo de pólen deixa de crescer no estilo superior, impedindo a fertilização(Figura 2C). No entanto, os tubos de pólen de uma cultivar compatível, com um alelo S diferente, podem crescer através do estilo (Figura 2D),alcançar o ovário(Figura 2E) e fertilizar um dos dois ovulos.
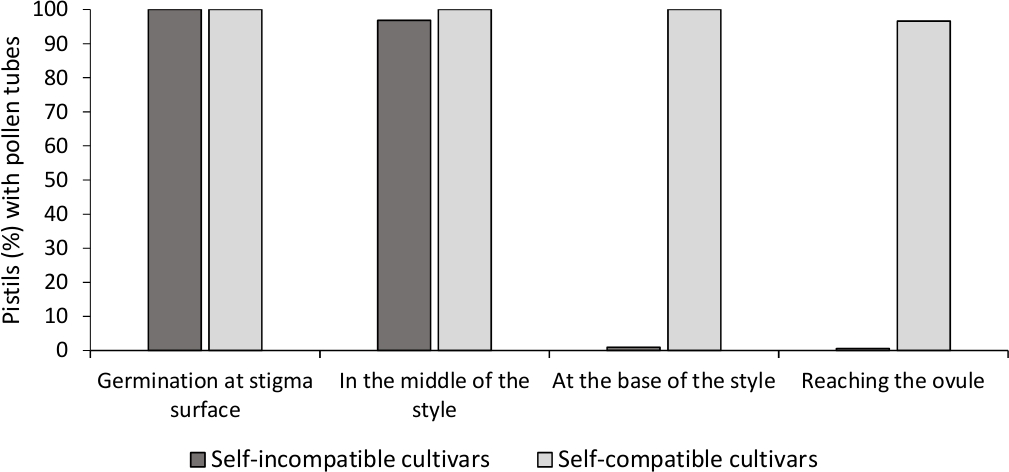
A análise da germinação in vitro de pólen mostrou boa viabilidade de pólen em todas as cultivares aqui analisadas, uma vez que a maioria dos tubos de pólen eram mais longos do que o comprimento do grão de pólen após 24 h no meio da cultura. Os grãos de pólen germinados foram observados na superfície do estigma(Figura 2B)em pistils de todas as polinização, indicando polinização adequada(Figura 3).
Para determinar a auto-compatibilidade para cada cultivar, observou-se o comportamento do tubo de pólen em polinizações auto e cruzadas feitas em condições controladas em laboratório sob microscopia de fluorescência. O crescimento do tubo de pólen foi registrado ao longo do estilo em todos os pistils examinados. As cultivares foram consideradas autocompatíveis quando o crescimento do tubo de pólen foi preso ao longo do estilo na maioria dos pistils auto-polinizados(Figura 2C, Figura 3) e autocompatíveis quando pelo menos um tubo de pólen atingiu a base do estilo na maioria dos pistils examinados(Figura 2E, Figura 3).
O estudo do S-locuspela análise pcr permitiu caracterizar o genótipo Sde cada cultivar. Em primeiro lugar, os selados Sforam identificados pela amplificação do primeiro intron S-RNase utilizando os primers SRc-F/SRc-R (Tabela 2). O tamanho dos fragmentos amplificados foi analisado por eletroforese capilar(Figura 4A) e foi utilizado para classificar os genótipos analisados em seu grupo de incompatibilidade correspondente (I.G.) (Tabela 3).
Alguns pares de alelos, como S1 e S7 ou S6 e S9,apresentaram tamanhos de fragmentos semelhantes para o primeiro intron. Assim, a diferenciação desses alelos foi feita ampliando uma região do segundo intron da RNase com os primers Pru-C2/PruC4R, SHLM1/SHLM2 e SHLM3/SHLM4 (Tabela 2). A combinação de primer PruC2/PruC4R foi usada para distinguir entre S6 e S9. Para S6, foi amplificado um fragmento de 1300 bp, enquanto um fragmento de cerca de 700 bp foi observado para o alelo S9 ( Figura4B, Tabela 3). Os primers específicos SHLM1/SHLM2 e SHLM3/SHLM4 amplificaram um fragmento de aproximadamente 650 bp no alelo S1 e 413 bp no alelo S7 ( Figura4C, Tabela 3).
Os primers AprFBC8-(F/R) que amplificam as regiões variáveis V2 e HVb do gene SFB foram usados para distinguir alelos Sc e S8, uma vez que ambos os alelos apresentam sequência RNase idêntica. O alelo S8 mostrou um fragmento de PCR de aproximadamente 150 bp, enquanto um fragmento de 500 bp correspondia ao alelo Sc ( Figura4D,Tabela 3). Uma vez determinado o S-genótipo para todas as cultivares, cultivares auto-incompatíveis foram atribuídas aos seus grupos de incompatibilidade correspondentes com base em seus S-alelos(Tabela 3).
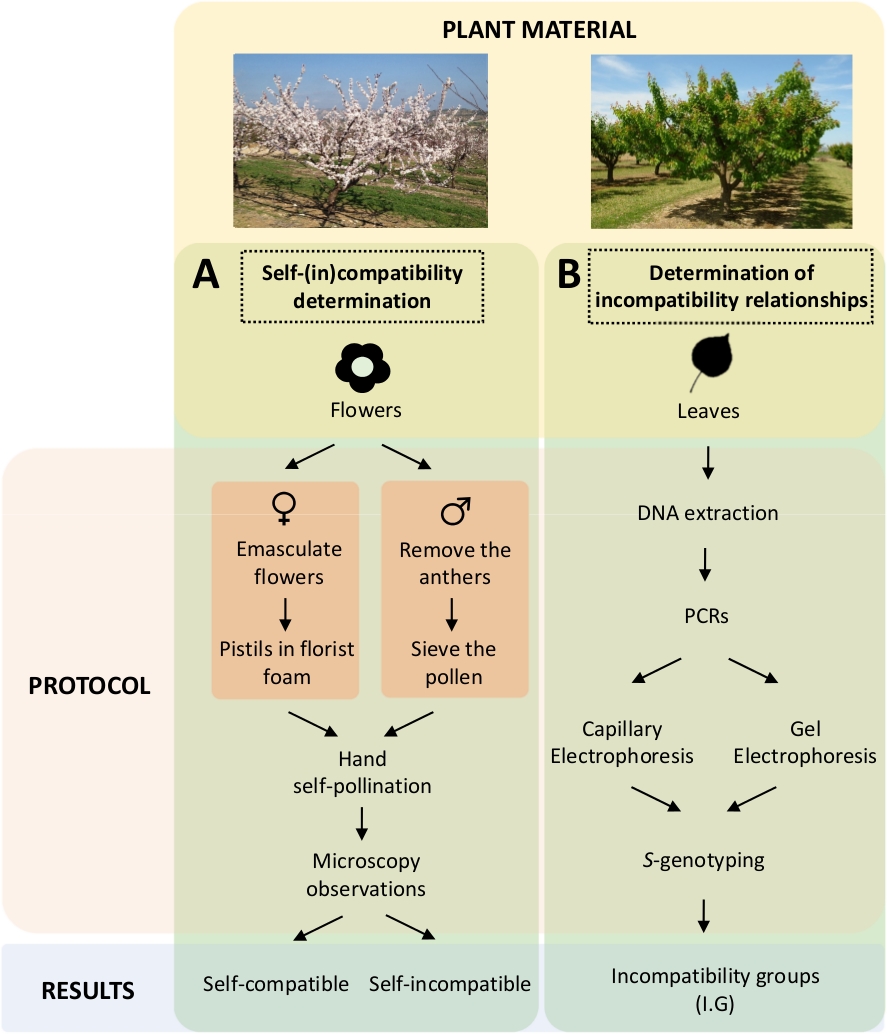
Essa abordagem requer determinar a auto-(compatibilidade de cada cultivar por auto-polinização controlada e cruzada em laboratório (Figura 5A) concomitantemente com a caracterização do S-genótipo por análise genética(Figura 5B). Como resultado, podem ser determinados os requisitos de polinização de cada cultivar e as relações de incompatibilidade entre as cultivares de damasco.

Figura 1. Configuração experimental para a determinação da auto-(in)compatibilidade no damasco.
(A) Flores no estágio do balão (flechas pretas) no campo. (B) Peneira de grãos de pólen usando uma malha fina. (C) Pistils colocados na espuma florista na água. (D) Polinização manual dos pistils com a ajuda de um pincel. Clique aqui para ver uma versão maior desta figura.

Figura 2. Representação diagramática das relações de incompatibilidade gametofítica nas flores de damasco.
(A) Em Autoincompatibilidade Gametofística (GSI), tanto os grãos de pólen compatíveis quanto incompatíveis germinam sobre o estigma. O grão de pólen carrega um dos dois S-alelosdo genótipo original, neste caso s1 ou S2. Se o S-alelos do grão de pólen corresponder a um dos dois S-alelosdo pistil, neste caso S1S3, o crescimento do tubo de pólen é inibido no um terço superior do estilo. (B) Germinação de grãos de pólen na superfície do estigma. (C) Tubo de pólen preso no estilo indicando um comportamento incompatível. (D) Tubos de pólen crescendo ao longo do estilo. (E) Tubos de pólen na base do estilo indicando um comportamento compatível. Barras de escala, 100 μm. Clique aqui para ver uma versão maior desta figura.

Figura 3. Resultados representativos da germinação de pólen e crescimento do tubo de pólen através do estilo para cultivares autocompatíveis e auto-incompatíveis após auto-polinização.
Percentual de pistils com grãos de pólen germinando na superfície do estigma, com tubos de pólen na metade do estilo, na base do estilo, e atingindo o ovulo. Clique aqui para ver uma versão maior desta figura.

Figura 4. Amplificação de fragmentos PCR usando cinco combinações de pares de primer para a identificação de alelos S.
(A) Saída do analisador genético para os primers SRc-(F/R) mostrando o tamanho dos dois fragmentos amplificados da primeira região intron da RNase correspondente aos alelos S. (B) Amplificação PCR utilizando os primers PruC2/PruC4R para a identificação dos alelos S6 e S9. (C) Produtos PCR obtidos utilizando os primers específicos SHLM1 e SHLM2 para a diferenciação do alelo S11 e SHLM3 e SHLM4 para distinguir o alelo S7. (D) amplificação PCR com os primers AprFBC8-(F/R) para identificação de alelos Sc e S8. MI: 1 kb escada de DNA. MII: Escada de DNA de 100 bp. Clique aqui para ver uma versão maior desta figura.

Figura 5. Esquema do projeto experimental para elucidar as relações de compatibilidade auto e inter-(in) em cultivares de damasco.
(A) Fluxo de trabalho de determinação de auto-(in)compatibilidade por polinizações controladas em laboratório. (B) Fluxo de trabalho da identificação de S-alelopor abordagens moleculares. Clique aqui para ver uma versão maior desta figura.
| PCR Master Mix | Condições do termciclador | ||||||
| Componentes | Concentração Final | Reação de 15 μL | Passo de ciclo | Temperatura | Tempo | Ciclos | |
| 10x NH4 Tampão de Reação | 10x | 1,5 μL | Desnaturação inicial | 94 °C | 3 min. | 1 | |
| Solução mgcl2 de 50 mM | 25 mM | 1,2 μL | Desnaturação | 94 °C | 1 min | 35 | |
| 100 mM dNTP | 2,5 mM | 0,6 μL | Recozimento | 55 °C | 1 min | ||
| Primer SRc-F | 10 μM | 0,6 μL | Extensão | 72 °C | 3 min. | ||
| Primer SRc-R | 10 μM | 0,6 μL | Extensão Final | 72 °C | 5 min. | 1 | |
| 500 U Taq DNA Polymerase | 0.5 U | 0,2 μL | 4 °C | Segurar | |||
| H2O | 8,3 μL | ||||||
| Componentes | Concentração Final | Reação de 25 μL | Passo de ciclo | Temperatura | Tempo | Ciclos | |
| Tampão PCR 10x | 10x | 2,5 μL | Desnaturação inicial | 94 °C | 2 min. | 1 | |
| 5x solução Q | 5x | 5 μL | Desnaturação | 94 °C | 10 s | 10 | |
| 100 mM dNTP | 2,5 mM | 0,5 μL | Recozimento | 55 °C | 2 min. | ||
| Primer PruC2 | 10 μM | 0,2 μL | Extensão | 68 °C | 2 min. | ||
| Primer C4R | 10 μM | 0,2 μL | Desnaturação | 94 °C | 10 s | 25 | |
| 250 U Taq DNA Polymerase | 10 U | 0,13 μL | Recozimento | 58 °C | 2 min. | ||
| H2O | 15,5 μL | Extensão* | 68 °C | 2 min. | |||
| Extensão Final | 72 °C | 5 min. | 1 | ||||
| 4 °C | Segurar | ||||||
| * com 10 s adicionados a cada ciclo à etapa de extensão de 68 %C. | |||||||
| Componentes | Concentração Final | Reação de 25 μL | Passo de ciclo | Temperatura | Tempo | Ciclos | |
| Tampão PCR 10x | 10x | 2,5 μL | Desnaturação inicial | 94 °C | 2 min. | 1 | |
| 5x solução Q | 5x | 5 μL | Desnaturação | 94 °C | 30 s | 35 | |
| 100 mM dNTP | 2,5 mM | 0,5 μL | Recozimento | 62 °C | 1,5 min. | ||
| Primer SHLM1 | 10 μM | 0,2 μL | Extensão | 72 °C | 2 min. | ||
| Primer SHLM2 | 10 μM | 0,2 μL | Extensão Final | 72 °C | 5 min. | 1 | |
| 250 U Taq DNA Polymerase | 10 U | 0,13 μL | 4 °C | Segurar | |||
| H2O | 15,5 μL | ||||||
| Componentes | Concentração Final | Reação de 20 μL | Passo de ciclo | Temperatura | Tempo | Ciclos | |
| 5x buffer PCR | 5x | 4 μL | Desnaturação inicial | 98 °C | 30 s | 1 | |
| dNTP | 2,5 mM | 1,6 μL | Desnaturação | 98 °C | 10 s | 35 | |
| Primer SHLM3 | 10 μM | 1 μL | Recozimento | 51 °C | 30 s | ||
| Primer SHLM4 | 10 μM | 1 μL | Extensão | 72 °C | 1 min | ||
| 100 U DNA Polymerase | 5 U | 0,2 μL | Extensão Final | 72 °C | 5 min. | 1 | |
| H2O | 12,4 μL | 4 °C | Segurar | ||||
| Componentes | Concentração Final | Reação de 25 μL | Passo de ciclo | Temperatura | Tempo | Ciclos | |
| Tampão PCR 10x | 10x | 2,5 μL | Desnaturação inicial | 94 °C | 2 min. | 1 | |
| 100 mM dNTP | 2,5 mM | 2 μL | Desnaturação | 94 °C | 30 s | 35 | |
| Primer FBC8-F | 10 μM | 1 μL | Recozimento | 55 °C | 1,5 min. | ||
| Primer FBC8-R | 10 μM | 1 μL | Extensão | 72 °C | 2 min. | ||
| 250 U Taq DNA Polymerase | 10 U | 0,125 μL | Extensão Final | 72 °C | 5 min. | 1 | |
| H2O | 17,4 μL | 4 °C | Segurar | ||||
Mesa 1. Condições de reação e ciclismo para diferentes combinações de primer usadas neste protocolo.
| Primers | Seqüência | Referência |
| SRc-F | 5'-CTCGCTTTCCTTGTTCTTGC-3' | 18 |
| SRc-R | 5'-GGCCATTGTTGCACCCCTTG-3' | 18 |
| Pru-C2 | 5'-CTTTGGCCAAGTAATTATTCAACC-3' | 35 |
| Pru-C4R | 5'-GGATGTGGTACGATTGAAGCG-3' | 35 |
| SHLM1-F | 5'-GGTGGAGGTGATAAGGTAGCC-3' | 17 |
| SHLM2-R | 5'-GGCTGCATAAGGAAGCTGTAGG-3' | 17 |
| SHLM3-F | 5'-TATATCTTACTCTTTGGC-3' | 17 |
| SHLM4-R | 5'-CACTATGATAATGTGTATG-3' | 17 |
| AprFBC8-F | 5'-CATGGAAAAAGCTGACTTATGG-3' | 26 |
| AprFBC8-R | 5'-GCCTCTAATGTCATCTCTCTTAG-3' | 26 |
Mesa 2. Primers utilizados neste protocolo, sequência e referência para caracterização de S-genótipoem Prunus armeniaca.
| Cultivar | SRc-(F/R) (bp) | PruC2/PruC4R (bp) | SHLM1/SHLM2 (bp) | SHLM3/SHLM4 (bp) | AprFBC8-(F/R) (bp) | S-Genótipo | Grupo de incompatibilidade (I.G) |
| Berço Maravilha8 | 420, 420 | 749, 1386 | S6S9 | Viii | |||
| Berço Mágico8 | 334, 420 | 749 | S2S9 | Xx | |||
| Goldstrike8 | 334, 420 | 749 | S2S9 | - | |||
| T06917 | 334, 408 | 650 | S1S2 | Eu | |||
| T12017 | 334, 408 | 650 | S1S2 | - | |||
| C-6 | 334, 408 | 413 | S2S7 | Iv | |||
| Cooper Cot8 | 274, 408 | 650 | S1S3 | XVIII | |||
| Apriqueen | 358, 358 | 500 | ScSc | - | |||
| Bergecot8 | 334, 358 | 500 | S2Sc | - | |||
| Blush primavera8 | 274, 358 | 150 | S3S8 | XXI |
Mesa 3. S-genotipagem de cultivares de damasco com cinco pares de primer usados neste protocolo e atribuição de grupo de incompatibilidade. Os diferentes tamanhos de produtos de reação em cadeia de polimerase de alelosS-alelos amplificados usando primers SRc-(F/R), PruC2/PruC4R, SHLM1/ SHLM2, SHLM3/SHLM4 e AprFBC8-(F/R) são mostrados na tabela.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Tradicionalmente, a maioria das cultivares europeias de damasco comercial eram autocompatíveis36. No entanto, o uso de cultivares autocompatíveis norte-americanas como pais em programas de reprodução nas últimas décadas resultou na liberação de um número crescente de novas cultivares auto-incompatíveis com requisitos de polinização desconhecidos7,,8,,37. Assim, a determinação das relações de auto e inter-(in)compatibilidade nas cultivares de damasco é cada vez mais importante. Isso se acentua nas áreas onde o frio do inverno está diminuindo, uma vez que as variações de alto ano a ano na época da floração estão prevenindo a coincidência na floração de cultivares e seus polinizadores em muitos casos, especialmente em cultivares com altas exigências de refrigeração38. A metodologia aqui descrita, combinando polinização manual, microscopia e análises genéticas tem sido muito útil para determinar a auto(in)compatibilidade de cada cultivar e estabelecer suas potenciais cultivares polinizadores.
Os requisitos de polinização podem ser determinados através de experimentos de controle de campo nas condições do pomar11,39. No entanto, a exposição a fatores externos, incluindo condições adversas meteorológicas, pode causar falha na polinização10, o que pode resultar em diagnósticos errôneos de autoincompatibilidade. A metodologia aqui descrita permite avaliar a auto-(in)compatibilidade com mais precisão por observações de microscopia de flores polinizados à mão em condições controladas por laboratório, evitando influência ambiental. Além disso, essa abordagem permite analisar um maior número de cultivares por ano, uma vez que apenas um pequeno número de flores é necessário em vez de várias árvores adultas para cada cultivar que são necessárias em experimentos de campo40.
Relações de incompatibilidade podem ser estabelecidas combinando polinizações manuais e microscopia14. No entanto, as polinizações só podem ser realizadas por um curto período durante a estação de floração na primavera, e são necessárias árvores adultas perto do laboratório, uma vez que a vida útil das flores coletadas é muito curta. Assim, o número de relações de incompatibilidade que podem ser analisadas por polinizações manuais controladas em cada estação é muito baixo. A caracterização dos genes codificados pelo lócus Spermitiu o desenvolvimento de métodos baseados em PCR para genotipagem de s-alelo18,41. Essa abordagem acelera a identificação de S-alelo, uma vez que não requer flores, e os experimentos podem ser realizados com qualquer tecido vegetativo42. S Isso amplia o período durante o qual o material vegetal, geralmente folhas jovens, pode ser coletado43. Além disso, as folhas podem ser liofilizadas ou congeladas, de modo que a análise possa ser feita em qualquer época do ano, ao contrário das polinizações que só podem ser feitas em flores frescas durante a estação de floração44. Um benefício adicional é que as folhas podem ser coletadas de árvores jovens antes mesmo de entrar em idade de floração, facilitando a coleta de amostras e a obtenção precoce dos resultados45.
A análise genética permite uma melhor diferenciação dos alelos de autoincompatibilidade, uma vez que fornece resultados precisos dos tamanhos de fragmento amplificados21,46. Até o momento, foram identificados 33 S-alelosno damasco12,18,20,21,22,23,24, que permitiram estabelecer 36 grupos de incompatibilidade com base no S-genótipo8,,9,,17,25,,26,27. Por outro lado, uma desvantagem dessa metodologia é que diferentes alelos do mesmo tamanho ou mutações podem ser erroneamente identificados como o mesmo alelo. Assim, os alelos Sc e S8 são idênticos para a sequência RNase, mas uma inserção de 358 bps é encontrada no gene SFB de Sc19. Da mesma forma, a primeira região intron dos alelos S1 e S7 são idênticas e são indistinguíveis usando os primers SRc-F/SRc-R. Além disso, vários homólogos, como S6 e S528 ou S20 e S55, e S7, S13 (EF062341) e S4617,foram encontrados porque alguns desses alelos foram parcialmente sequenciados ou por falhas durante a amplificação da PCR e, consequentemente, mais trabalhos são necessários para distingui-los corretamente.
A análise de PCR e o sequenciamento S-RNase são adequados para estabelecer relações de incompatibilidade através da identificação de S-alelose da alocação de cultivares em seu correspondente Grupo de Incompatibilidade8,,17,,26,,27. No entanto, essa metodologia tem a limitação de impedir a determinação da auto-(in)compatibilidade para cultivares particulares de damasco. A auto-compatibilidade (SC) tem sido associada a alelos Sparticulares em outras espécies de Prunus 47, como amêndoa(Sf)48,,49 ou cereja doce(S4')50,51. No entanto, no damasco, o alelo Sc, que tem sido associado à SC21,pode ser erroneamente identificado como S8, um alelo auto-incompatível19,22, e possíveis mutações não ligadas ao lócus S, como o M-locus12,52, conferenciando SC foram identificados. Recentemente, Mo M-locus foi genótipo usando marcadores SSR12. Portanto, a identificação genética de SC para genótipos de damasco precisa de mais pesquisas e, para evitar erros devido a fatores não ligados ao lócus S, neste trabalho a caracterização da auto-(in)compatibilidade tem sido determinada também por fenotipar o comportamento dos tubos de pólen através do pistil das flores auto-polinizado.
A metodologia aqui descrita combina a determinação da auto-(in)compatibilidade por polinizações manuais em condições laboratoriais com a observação subsequente do comportamento dos tubos de pólen Sno pistil das auto-polinizações controladas sob a microscopia de fluorescência e a identificação do S-genótipo pela análise pcr permite estabelecer os requisitos de polinização dos cultivares de damasco. Isso fornece uma informação valiosa para os produtores e criadores, uma vez que permite estabelecer as relações de incompatibilidade entre cultivares para escolher polinizadores adequados no projeto de novos pomares, bem como selecionar pais apropriados para projetar novas cruzes em programas de criação de damascos.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Os autores não têm nada a revelar.
Acknowledgments
Esta pesquisa foi financiada pelo Ministerio de Ciencia, Innovación y Universidades-European Regional Development Fund, União Europeia (AGL2016-77267-R e AGL2015-74071-JIN); Instituto Nacional de Investigação y Tecnología Agraria y Alimentaria (RFP2015-00015-00, RTA2017-00003-000); Gobierno de Aragón-European Social Fund, União Europeia (Grupo Consolidado A12_17R), Fundação Biodiversidad e Agroseguro S.A.
Materials
| Name | Company | Catalog Number | Comments |
| Agarose D1 Low EEO | Conda | 8010.22 | |
| BIOTAQ DNA Polymerase kit | Bioline | BIO-21060 | |
| Bright field microscope | Leica Microsystems | DM2500 | |
| CEQ System Software | Beckman Coulter | ||
| DNeasy Plant Mini Kit | QIAGEN | 69106 | |
| dNTP Set, 4 x 25 µmol | Bioline | BIO-39025 | |
| GenomeLab DNA Size Standard Kit - 400 | Beckman Coulter | 608098 | |
| GenomeLab GeXP Genetic Analysis System | Beckman Coulter | ||
| GenomeLab Separation Buffer | Beckman Coulter | 608012 | |
| GenomeLab Separation Gel LPA-1 | Beckman Coulter | 391438 | |
| HyperLadder 100bp | Bioline | BIO-33029 | |
| HyperLadder 1kb | Bioline | BIO-33025 | |
| Image Analysis System | Leica Microsystems | ||
| Molecular Imager VersaDoc MP 4000 system | Bio-Rad | 170-8640 | |
| NanoDrop One Spectrophotometer | Thermo Fisher Scientific | 13-400-518 | |
| pH-Meter BASIC 20 | Crison | ||
| Phusion High-Fidelity PCR Kit | Thermo Fisher Scientific | F553S | |
| Power Pack P 25 T | Biometra | ||
| Primer Forward | Isogen Life Science | ||
| Primer Reverse | Isogen Life Science | ||
| Quantity One Software | Bio-Rad | ||
| Stereoscopic microscope | Leica Microsystems | MZ-16 | |
| Sub-Cell GT | Bio-Rad | ||
| SYBR Safe DNA Gel Stain | Thermo Fisher Scientific | S33102 | |
| T100 Thermal Cycler | Bio-Rad | 1861096 | |
| Taq DNA Polymerase | QIAGEN | 201203 | |
| Vertical Stand Autoclave | JP Selecta |
References
- Silva, N. F., Goring, D. R.
- Charlesworth, D., Vekemans, X., Castric, V., Glémin, S. Plant self-incompatibility systems: A molecular evolutionary perspective. New phytologist. 168, 61-69 (2005).
- Tao, R., et al. Identification of stylar RNases associated with gametophytic self-incompatibility in almond (Prunus dulcis). Plant and Cell Physiology. 38, 304-311 (1997).
- Ushijima, K., et al. Structural and transcriptional analysis of the self-incompatibility locus of almond: Identification of a pollen-expressed F-box gene with haplotype-specific polymorphism. The Plant cell. 15, 771-781 (2003).
- Bedinger, P. A., Broz, A. K., Tovar-Mendez, A., McClure, B. Pollen-Pistil Interactions and Their Role in Mate Selection. Plant Physiology. 173, 79-90 (2017).
- Guerra, M. E., Rodrigo, J. Japanese plum pollination: A review. Scientia Horticulturae. 197, 674-686 (2015).
- Zhebentyayeva, T., Ledbetter, C., Burgos, L., Llacer, G. Apricot. Fruit Breeding. Badenes, M. L., Byrne, D. , Springer. Boston, MA. 415-458 (2012).
- Herrera, S., Lora, J., Hormaza, J. I., Herrero, M., Rodrigo, J. Optimizing Production in the New Generation of Apricot Cultivars: Self-incompatibility, S-RNase Allele Identification, and Incompatibility Group Assignment. Frontiers in Plant Science. 9, 527 (2018).
- Egea, J., Burgos, L. Detecting Cross-incompatibility of Three North American Apricot Cultivars and Establishing the First Incompatibility Group in Apricot. Journal of the American Society for Horticultural Science. 121, 1002-1005 (1996).
- Rodrigo, J., Herrero, M. Effects of pre-blossom temperatures on flower development and fruit set in apricot. Scientia Horticulturae. 92, 125-135 (2002).
- Julian, C., Herrero, M., Rodrigo, J. Flower bud differentiation and development in fruiting and non-fruiting shoots in relation to fruit set in apricot (Prunus armeniaca). Trees. 24, 833-841 (2010).
- Muñoz-Sanz, J. V., Zuriaga, E., López, I., Badenes, M. L., Romero, C. Self-(in)compatibility in apricot germplasm is controlled by two major loci, S and M. BMC Plant Biology. 17, 82 (2017).
- Burgos, L., Berenguer, T., Egea, J. Self- and Cross-compatibility among Apricot Cultivars. HortScience. 28, 148-150 (1993).
- Rodrigo, J., Herrero, M. Evaluation of pollination as the cause of erratic fruit set in apricot "Moniqui". Journal of Horticultural Science. 71, 801-805 (1996).
- Milatović, D., Nikolić, D., Krška, B. Testing of self-(in)compatibility in apricot cultivars from European breeding programmes. Horticultural Science. 40 (2), 65-71 (2013).
- Milatović, D., Nikolić, D., Fotirić-Aksić, M., Radović, A. Testing of self-(in)compatibility in apricot cultivars using fluorescence microscopy. Acta Scientiarum Polonorum, Hortorum Cultus. 12 (6), 103-113 (2013).
- Herrera, S., Rodrigo, J., Hormaza, J. I., Lora, J. Identification of Self-Incompatibility Alleles by Specific PCR Analysis and S-RNase Sequencing in Apricot. Int J Mol Sci. 19, 3612 (2018).
- Romero, C., et al. Analysis of the S-locus structure in Prunus armeniaca L. Identification of S-haplotype specific S-RNase and F-box genes. Plant Molecular Biology. 56, 145-157 (2004).
- Halász, J., Pedryc, A., Hegedus, A. Origin and dissemination of the pollen-part mutated SC haplotype which confers self-compatibility in apricot (Prunus armeniaca). New Phytologist. 176, 792-803 (2007).
- Halász, J., Hegedus, A., Hermán, R., Stefanovits-Bányai, É, Pedryc, A. New self-incompatibility alleles in apricot (Prunus armeniaca L.) revealed by stylar ribonuclease assay and S-PCR analysis. Euphytica. 145, 57-66 (2005).
- Vilanova, S., Romero, C., Llacer, G., Badenes, M. L., Burgos, L. Identification of Self-(in)compatibility Alleles in Apricot by PCR and Sequence Analysis. Journal of the American Society for Horticultural Science. 130, 893-898 (2005).
- Feng, J., et al. Detection and transcript expression of S-RNase gene associated with self-incompatibility in apricot (Prunus armeniaca L.). Molecular Biology Reports. 33, 215-221 (2006).
- Zhang, L., et al. Identification of self-incompatibility (S-) genotypes of Chinese apricot cultivars. Euphytica. 160, 241-248 (2008).
- Wu, J., et al. Identification of S-haplotype-specific S-RNase and SFB alleles in native Chinese apricot (Prunus armeniaca L). Journal of Horticultural Science and Biotechnology. 84, 645-652 (2009).
- Szabó, Z., Nyéki, J. Blossoming, fructification and combination of apricot varieties. Acta Horticulturae. 293, 295-302 (1991).
- Halász, J., Pedryc, A., Ercisli, S., Yilmaz, K. U., Hegedűs, A. S-genotyping supports the genetic relationships between Turkish and Hungarian apricot germplasm. Journal of the American Society for Horticultural Science. 135, 410-417 (2010).
- Lachkar, A., et al. Identification of self-(in)compatibility S-alleles and new cross-incompatibility groups in Tunisian apricot (Prunus armeniaca L.) cultivars. The Journal of Horticultural Science and Biotechnology. 88, 497-501 (2013).
- Pérez-Pastor, A., Ruiz-Sánchez, M. C., Domingo, R., Torrecillas, A. Growth and phenological stages of Búlida apricot trees in South-East. Agronomie. 24, 93-100 (2004).
- Williams, J. H., Friedman, W. E., Arnold, M. L. Developmental selection within the angiosperm style: using gamete DNA to visualize interspecific pollen competition. Proceedings of the National Academy of Sciences of the United States of America. 96, 9201-9206 (1999).
- Julian, C., Herrero, M., Rodrigo, J. Anther meiosis time is related to winter cold temperatures in apricot (Prunus armeniaca L.). Environmental and Experimental Botany. 100, 20-25 (2014).
- Guerra, M. E., López-Corrales, M., Wünsch, A., Rodrigo, J. Lack of Fruit Set Caused by Ovule Degeneration in Japanese Plum. Journal of the American Society for Horticultural Science. 136 (6), 375-381 (2011).
- Guerra, M. E., Wünsch, A., López-Corrales, M., Rodrigo, J. Flower Emasculation as the Cause for Lack of Fruit Set in Japanese Plum Crosses. Journal of the American Society for Horticultural Science. 135 (6), 556-562 (2010).
- Hormaza, J. I., Pinney, K., Polito, V. S. Correlation in the tolerance to ozone between sporophytes and male gametophytes of several fruit and nut tree species (Rosaceae). Sexual Plant Reproduction. 9, 44-48 (1996).
- Alcaraz, M. L., Hormaza, J. I., Rodrigo, J. Pistil Starch Reserves at Anthesis Correlate with Final Flower Fate in Avocado (Persea americana). PLOS ONE. 8 (10), 78467 (2013).
- Tao, R., et al. Molecular typing of S-alleles through Identification, Characterization and cDNA cloning for S-RNases in Sweet Cherry. Journal of the American Society for Horticultural Science. 124, 224-233 (1999).
- Burgos, L., et al. The self-compatibility trait of the main apricot cultivars and new selections from breeding programmes. Journal of Horticultural Science. 72, 147-154 (1997).
- Hormaza, J. I., Yamane, H., Rodrigo, J. Apricot. Genome Mapping and Molecular Breeding in Plants, Volume 4 Fruits and Nuts. Kole, C. , Springer. New York, NY. 171-187 (2007).
- Benmoussa, H., Ghrab, M., Ben Mimoun, M., Luedeling, E. Chilling and heat requirements for local and foreign almond (Prunus dulcis Mill.) cultivars in a warm Mediterranean location based on 30 years of phenology records. Agricultural and Forest Meteorology. 239, 34-46 (2017).
- Rodrigo, J., Herrero, M., Hormaza, J. I. Pistil traits and flower fate in apricot (Prunus armeniaca). Annals of Applied Biology. 154, 365-375 (2009).
- Williams, R. R. Techniques used in fruit-set experiments. Towards Regulated Cropping. Williams, R. R., Wilson, D. , Grower Books. London. 57-61 (1970).
- Sutherland, B. G., Robbins, T. P., Tobutt, K. R. Primers amplifying a range of Prunus S-alleles. Plant Breeding. 123, 582-584 (2004).
- Murray, M. G., Thompson, W. F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Research. 8, 4321-4325 (1980).
- Porebski, S., Bailey, L. G., Baum, B. R. Modification of a CTAB DNA Extraction Protocol for Plants Containing High Polysaccharide and Polyphenol Components. Plant Molecular Biology Reporter. 15 (1), 8-15 (1997).
- Rogers, S. O., Bendich, A. J. Extraction of DNA from milligram amounts of fresh, herbarium and mummified plant tissues. Plant Molecular Biology. 5 (2), 69-76 (1985).
- Hormaza, J. I. Molecular characterization and similarity relationships among apricot (Prunus armeniaca L.) genotypes using simple sequence repeats. Theoretical and Applied Genetics. 104, 321-328 (2002).
- Sonneveld, T., Tobutt, K. R., Robbins, T. P. Allele-specific PCR detection of sweet cherry self-incompatibility (S) alleles S1 to S16 using consensus and allele-specific primers. Theoretical and Applied Genetics. 107, 1059-1070 (2003).
- Hegedus, A., Lénárt, J., Halász, J. Sexual incompatibility in Rosaceae fruit tree species: molecular interactions and evolutionary dynamics. Biologia Plantarum. 56 (2), 201-209 (2012).
- Fernández i Martí, A., Gradziel, T. M., Socias i Company, R. Methylation of the Sf locus in almond is associated with S-RNase loss of function. Plant Molecular Biology. 86, 681-689 (2014).
- Company, R. S. i, Kodad, O., Martí, A. F. i, Alonso, J. M. Mutations conferring self-compatibility in Prunus species: From deletions and insertions to epigenetic alterations. Scientia Horticulturae. 192, 125-131 (2015).
- Boskovic, R., Tobutt, K. R. Correlation of stylar ribonuclease zymograms with incompatibility alleles in sweet cherry. Euphytica. 90, 245-250 (1996).
- Cachi, A. M., Wünsch, A. S-genotyping of sweet cherry varieties from Spain and S-locus diversity in Europe. Euphytica. 197 (2), 229-236 (2014).
- Zuriaga, E., et al. An S-locus Independent Pollen Factor Confers Self-Compatibility in "Katy" Apricot. PLoS ONE. 8 (1), 53947 (2013).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}