

Summary
Представляем методологию определения требований к опылинию сортов абрикоса(Prunus armeniaca L.), сочетающих определение самостоятельной (в) совместимости флуоресценции микроскопией с идентификацией S-генотипа с помощью анализа ПЦР.
Abstract
Самосовместимость в розацеи определяется гаметофитической системой самосовместимости (GSI), которая в основном контролируется мультиаллеловым локусом S. В абрикосе определение отношений между собой и между собой приобретает все большее значение, поскольку высвобождение значительного числа новых сортов привело к увеличению сортов с неизвестными требованиями к опылению. Здесь мы описываем методологию, которая сочетает в себе определение самостоятельности (в) совместимости путем S-ручных опыления и микроскопии с определением S-генотипа путем анализа ПЦР. Для самостоятельного (в) определение совместимости, цветы на воздушном шаре этапе от каждого сорта были собраны в поле, ручной опыляется в лаборатории, фиксированной, и окрашенные с анилиновым синим для наблюдения поведения трубки пыльцы под флуоресценции микроскопии. Для установления несовместимости отношений между сортами, ДНК из каждого сорта Sбыла извлечена из молодых листьев и S-аллели были определены ПЦР. Такой подход позволяет создавать группы несовместимости и выясывать несовместимость отношений между сортами, что дает ценную информацию для выбора подходящих опылителей при проектировании новых садов и выбора соответствующих родителей в программах разведения.
Introduction
Самосовместимость является стратегия цветения растений для предотвращения самоопыления и содействия outcrossing1. В Rosaceae этот механизм определяется гаметофитической системой самосовместимости (GSI), которая в основном контролируется многоаллюровым локусом S2. В стиле, ген RNase кодирует S-stylar determinant, RNase3, в то время как белок F-box, который определяет S-пыльцудетерминант, кодифицирован геном SFB 4. Взаимодействие с автосмешазнанием происходит через ингибирование роста пыльцевой трубки по стилю, предотвращающее оплодотворение яйцеклетки5,,6.
В абрикосе, разнообразие обновления состоялась во всем мире в последние два десятилетия7,8. Это введение значительного количества новых сортов, из различных государственных и частных программ разведения, привело к увеличению сортов абрикоса с неизвестными требованиями опыления8.
Различные методологии были использованы для определения требований опыления в абрикосе. В поле, самостоятельной (в) совместимость может быть установлена контролируемых опыляний в клетке деревьев или в выхолощенных цветов, а затем записи процент фруктов набор9,10,11,12. Кроме того, в лаборатории были проведены контролируемые опыления полуинфицейной культурой цветов и анализ поведения пыльцы трубки под флуоресценцией микроскопии8,,13,,14,,15,,16,17. В последнее время молекулярные методы, такие как анализ пЦР и секвенирование, позволили характеристику несовместимости отношений на основе изучения генов RNase и SFB 18,19. В абрикосе, тридцать три S-аллелей были зарегистрированы (S1 до S20, S22 до S30, S52, S53, SV, Sх), в том числе один аллель, связанные с само совместимости (Sc)12,18,20,21,22,23,24. До сих пор, 26 групп несовместимости были стабилизированных в этом виде в соответствии сS-генотип8,9,17,25,26,27. Сорта с теми Sже S-аллелями несовместимы, в то время как сорта с по крайней мере с одним разным S-аллельом и, следовательно, распределенные в разных несовместимых группах, являются взаимосвязанными. S
Для определения требований к опыления абрикосовых сортов мы описываем методологию, которая сочетает в себе определение самостоятельной (в) совместимости флуоресценции микроскопии с определением S-генотипапутем анализа ПЦР в абрикосовых сортах. Такой подход позволяет создавать группы несовместимости и выяснение взаимосвязей между сортами.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. Определение самоуправства (в)совместимости
- Попробуйте цветы в поле. Необходимо собрать цветы на воздушном шаре этапе(рисунок 1А), соответствующие стадии 58 по шкале BBCH для абрикоса28, чтобы избежать нежелательных предыдущих опыления.
-
Само- и перекрестное опыляния в лаборатории
- Удалите пыльниты цветов на этапе воздушного шара и поместите их на лист бумаги, чтобы высохнуть при лабораторной температуре.
- После 24 ч, сито пыльцы зерна с помощью тонкой сетки (0,26 мм) (Рисунок 1B).
- Измождите группу из 30 цветов на одной и той же стадии воздушного шара для каждого самоопыления и перекрестного опыления и место ристилов на флорист пены в воде при лабораторной температуре(Рисунок 1C).
- Рука опыляет пестилицы с помощью кисти с пыльцой из цветов того же сорта 24 ч после выхолощенности. Кроме того, опылять другой набор пестиков каждого сорта с пыльцой из цветов совместимого опылителя в качестве контроля(рисунок 1D).
- После 72 ч, исправить пестилы в фиксаторный раствор этанола / уксусной кислоты (3:1) по крайней мере 24 ч при 4 кк29. Затем отбросьте фиксатор и добавьте 75% этанола, гарантируя, что образцы полностью погружены в раствор. Образцы могут быть сохранены в этом растворе при 4 градусов по Цельсию до использования8,,17,30,31,32.
-
Оценка жизнеспособности пыльцы через прорастание пыльцы in vitro
- Для подготовки среды прорастания, вес 25 г сахарозы, 0,075 г борной кислоты (H3BO3)и 0,075 г нитрата кальция (Ca(NO3)2)33.
- Добавьте компоненты среды в 250 мл дистиллированной воды и полностью растворить.
- Затвердеть среды добавив 2 г агарозы и перемешать, закрученного.
- Проверьте рН среды с помощью рН-метра и отрегулируйте значение до 7,0 с помощью решения NaOH или HCl.
- Автоклав смеси для стерилизации среды.
- После автоклавирования, охладить среду и распределить его в петри блюда в стерильных ламинар потока капот.
- Разбросите пыльцевые зерна тех же сортов, используемых для контролируемых опылений в затвердевшей среде прорастания пыльцы и наблюдайте их под микроскопом после 24 ч6.
ПРИМЕЧАНИЕ: Для стерилизации ламинарного вытяжки потока, очистить поверхность с 70% этанола и включите УФ лампы в течение 10 мин. - Храните блюда Петри в холодильнике при температуре 4 градусов по Цельсию до использования.
-
Наблюдения за микроскопией
- Вымойте пистилы три раза в течение 1 ч с дистиллированной водой и оставить их в 5% сульфита натрия при 4 градусах Цельсия. После 24 ч, автоклав их на 1 кг / см2 в течение 10 мин в сульфите натрия, чтобы смягчить ткани34.
- Поместите автоклавированные пестилы на стеклянную горку и, с помощью скальпеля, удалить трихом вокруг яичника, чтобы получить лучшую визуализацию пыльцы труб. Затем раздавите пестилы крышкой.
- Подготовка 0,1% (v/v) анилин синий пятно: смешать 0,1 мл анилин синий в 100 мл 0,1 N фосфат калия трибаза (K3PO4). Нанесите каплю анилинового синего на препараты, чтобы пятно калезы осаждения во время роста пыльцы трубки.
- Наблюдайте за пыльцевыми трубками по стилю с помощью микроскопа с УФ-эпифлуоресценцией с помощью 340-380 полос и 425 фильтров длиннопропасса.
2. Извлечение ДНК
- Образец 2-3 листья в поле весной. Рекомендуется пробовать листья на молодых стадиях, так как полученная ДНК имеет более высокое качество и более низкие уровни фенольных соединений по сравнению со старыми листьями.
- Извлеките геномную ДНК после шагов, описанных в коммерчески доступном комплекте (см. Таблицу Материалов).
- Проанализируйте количество и качество концентраций ДНК с помощью УФ-визави спектрофотометра (260 нм).
3. SS-аллельная идентификация
-
Настройка реакции ПЦР
- Подготовьте разбавление 50 нг/мл в дистиллированной воде каждого образца извлечения ДНК.
- Оттепель из РЦК реагентов медленно и держать их на льду. Оставьте полимеразу ДНК в морозильной камере до тех пор, пока это не понадобится.
- Подготовьте реакции усиления с использованием различных комбинаций грунтовок. Создайте смесь реакции ПЦР, объединив компоненты в таблице 1. Vortex реакции ПЦР хорошо перемешать и распределить объем, указанный для различных комбинаций грунтовок для каждого колодца ПЦР пластины. Затем добавьте 1 МЛ разбавления ДНК в каждом колодце.
- Поместите пластину ПЦР в термоциклер и запустите соответствующую программу ПЦР, показанную в таблице 1.
- Проанализируйте усиленные фрагменты. Есть в основном два различных способа анализа ПЦР усиленные фрагменты: капиллярный электрофорез (CE) с флуоресцентной этикеткой грунтовки или как визуализировать ампликоны агароз гель электрофорез с не-маркированные грунтовки.
-
Капилляр электрофорез
- Чтобы подготовить погрузочный буфер, смешайте 35 мл деионизированного формамида с 0,45 мл маркированного стандарта размеров. Vortex реагент хорошо перемешать, а затем обойтись 35,5 л в колодец читателя пластины.
- Добавьте 1 л продукта ПЦР в скважину. Кроме того, добавьте каплю минерального масла, чтобы предотвратить испарение воды.
- Подготовьте разделительную пластину, добавляя буфер разделения.
- Используйте коммерческое программное обеспечение, включенное в анализатор генов (см. Таблицу Материалов). Создайте новую пластину образца и сохраните имена образцов для всех скважин на пластине.
- Выберите метод анализа. В этом случае денатурируете образцы при 90 градусах по Цельсию на 120 с, вводят при 2,0 кВ на 30 с, и разделите на 6,0 кВ в течение 35 мин.
- Вставьте две пластины в ген анализатор. Заполните капиллярный массив дистиллированной водой.
- Загрузите запатентованный линейный гель полиакриламида (LPA). Наконец, нажмите Run.
-
Гель Электрофорез
- Подготовка 1% агарозного геля, добавляя 1,5 г молекулярной биологии класса агарозы в 150 мл 1x TAE (Трис-ацетат-ЭДТА) электрофорез работает буфер (40 мМ Трис, 20 мМ уксусной кислоты, и 1 мМ EDTA при pH 8.0). Растворите агарозу путем микроволнового нагрева на 2-3 мин.
- Чтобы визуализировать ДНК, добавьте 4 мл пятна нуклеиновой кислоты (см. Таблицу материалов) и аккуратно перемешайте.
- Добавьте гелевый гребень, с достаточным количеством колодцев для лестниц, элементов управления и образцов, в гелевый лоток. Затем медленно налейте смесь в середину геля лоток и избежать пузырьков.
- Пусть гель остыть в течение 30-45 мин при комнатной температуре, пока гель полностью затвердев. Введите гель в электрофорезной камере, удалите гелевый гребень и заполните камеру достаточным 1x буфером TAE, чтобы покрыть гель.
ПРИМЕЧАНИЕ: Проверьте размещение геля. Скважины должны быть помещены близко к отрицательному полюсу, так как отрицательно заряженная ДНК мигрирует в сторону катода. - Добавьте 5 мл погрузочного буфера (0,1% (v/v) бромофенол синий) в продукты PCR и хорошо перемешайте.
- Для оценки размера полос, загрузите 5 МЛ молекулярной лестницы веса ДНК (см. Таблица Материалов).
- Загрузите образцы в дополнительные колодцы геля.
- После того, как все образцы и ДНК молекулярной лестнице веса загружены, запустить гель на 90 В за 1-1,5 ч, пока синий краситель линии примерно на 75% длина геля.
- Визуализируйте полосы в трансиллюмиторе для нуклеиновых кислот.
-
Капилляр электрофорез
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Исследования опыления в абрикосе требуют использования цветов на поздней стадии воздушного шара за день до антеза(Рисунок 1А). Эта стадия считается наиболее благоприятной как для пыльцы, так и для коллекции пистилов, так как цветочные структуры почти зрелые, но атер дехисценции еще не произошло. Это предотвращает вмешательство нежелательной пыльцы, не только пыльцы из того же цветка, но и от других цветов, так как закрытые лепестки препятствуют прибытию насекомых, несущих внешнюю пыльцу. Зерна пыльцы легко просеиваются через тонкую сетку(рисунок 1B) из дехискаированных пыльных, ранее размещенных на листе бумаги в течение 24 ч при комнатной температуре или с небольшим дополнительным теплом. Кроме того, пестилы получаются из цветов на этапе воздушного шара после удаления лепестков, чашелистиков и тычинок с помощью пинцета или ногтей(рисунок 1C). Pistils может быть самостоятельной и кросс-опыляется с тонкой кистью (Рисунок 1D).
Гермафродитические цветы абрикоса имеют пять темно-красных чашелистиков, пять белых лепестков(рисунок 1А),один пестик(рисунок 2A)и 25-30 тычинок. Пистил имеет три основные структуры: стигма, стиль и яичник. Яичник имеет два яйцеклетки, и оплодотворение по крайней мере один из них требуется для установки плодов. Во время опыления насекомые, в основном пчелы, переносят пыльцу зерна в клеймо(рисунок 1А),где они прорастают(рисунок 2B)в пределах 24 ч после опыления. Пыльца трубки производится из каждого прорастающего зерна пыльцы, которая растет через структуры пестила, чтобы достичь яичника после 3-4 дней и удобрять один из двух яйцеклеток примерно через 7 дней. В самоухотных сортах, в которых аллель S пыльцы зерна такой же, как один из двух пестилов, пыльца трубки перестает расти в верхнем стиле, предотвращая оплодотворение (Рисунок 2C). Тем не менее, пыльца трубки из совместимого сорта, с другой аллель S, может расти через стиль (Рисунок 2D), достичь яичника (Рисунок 2E) и удобрять один из двух овул.
Анализ прорастания пыльцы in vitro показал хорошую жизнеспособность пыльцы во всех сортах, проанализированных здесь, так как большинство пыльцевых труб были длиннее, чем длина зерна пыльцы после 24 ч в среде культуры. Проминированные пыльцы зерна наблюдались на поверхности стигмы(Рисунок 2B) в пестиликах от всех опылений, что указывает на адекватное опыление(рисунок 3).
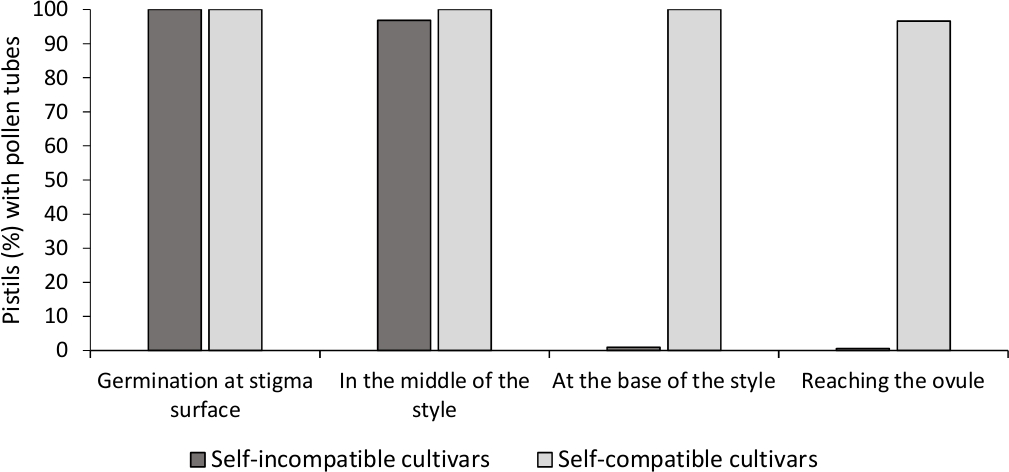
Для определения само-(в) совместимости для каждого сорта, поведение пыльцы трубки в само- и перекрестного опыления, сделанные в лабораторно контролируемых условиях наблюдалось под флуоресценции микроскопии. Рост пыльцы трубки был записан вдоль стиля во всех изученных пестиков. Cultivars были рассмотрены как самоподготовленные, когда рост пыльцы трубки был арестован по стилю в большинстве самоопиленных пестиков(Рисунок 2C, Рисунок 3) и самосовместимы, когда по крайней мере одна пыльца трубки достигли основания стиля в большинстве из пистилов рассмотрены (Рисунок 2E, Рисунок 3).
Изучение S-локусас помощью анализа ПЦР позволило охарактеризовать S-генотипкаждого сорта. Во-первых, S-аллелибыли определены путем усиления первого альтрона S-RNase с использованием грунтовок SRc-F/SRc-R(Таблица 2). Размер усиленных фрагментов был проанализирован капиллярным электрофорезом(рисунок 4A)и использовался для классификации генотипов, проанализированных в соответствующей группе несовместимости (I.G.) (Таблица 3).
Некоторые пары аллелей, такие как S1 и S7 или S6 и S9,показали аналогичные размеры фрагментов для первого интрона. Таким образом, дифференциация этих аллелей была сделана путем усиления области второго трона RNase с грунтовками Pru-C2/PruC4R, SHLM1/SHLM2 и SHLM3/SHLM4(Таблица 2). Комбинация грунтовок PruC2/PruC4R использовалась для различения S6 и S9. Для S6, фрагмент 1300 bp был усилен в то время как фрагмент около 700 bp наблюдался для аллеля S9 (Рисунок 4B, Таблица 3). Специфические грунтовки SHLM1/SHLM2 и SHLM3/SHLM4 усилили фрагмент примерно 650 б.п. в аллеле S1 и 413 bp в аллеле S7 (рисунок 4C, Таблица 3).
Праймеры AprFBC8-(F/R), которые усиливают переменные области V2 и HVb гена SFB, были использованы для различения аллелей Sc и S8, так как оба аллеля показывают идентичную последовательность RNase. Аллель S8 показал ПЦР-фрагмент примерно 150 bp, в то время как фрагмент 500 bp соответствовал аллельу Sc (рисунок 4D,таблица 3). После Sопределения S-генотипа для всех сортов, несовместимые с собой сорта были отнесены к соответствующим группам несовместимости на основе их S-аллелей(Таблица 3).
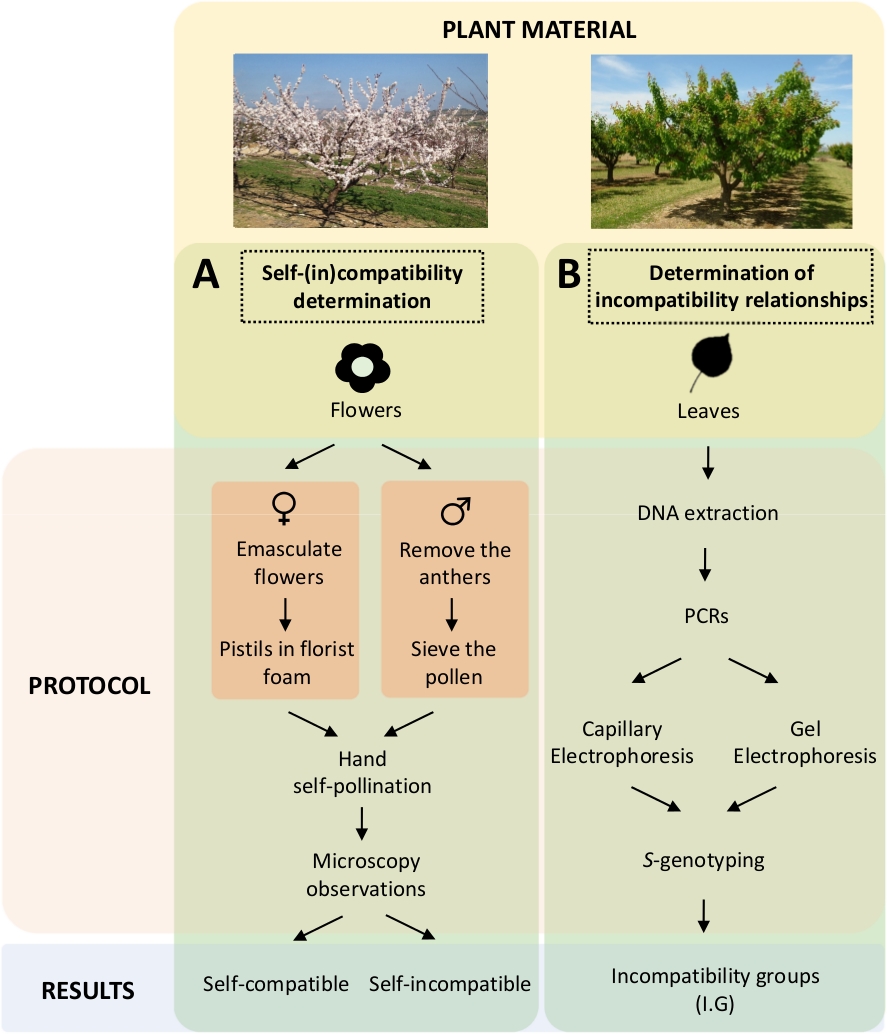
Этот подход требует определения самостоятельной (в) совместимости каждого сорта контролируемых само- и перекрестного опыляния в лаборатории(Рисунок 5А) одновременно с характеристикой S-генотипа по генетическому анализу(рисунок 5B). S В результате могут быть определены требования к опылиму каждого сорта и несовместимость между абрикосовыми сортами.

Рисунок 1. Экспериментальный создан для определения самостоятельной (в) совместимости в абрикосе.
(A) Цветы на воздушном шаре этапе (черные стрелки) в поле. (B) Сито пыльцы зерна с использованием тонкой сетки. (C) Pistils помещается на флористической пены в воде. (D) Ручное опыление пестили с помощью кисти. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой фигуры.

Рисунок 2. Схематическое представление игровых фитофитических взаимосвязей в абрикосовых цветах.
(A) В гаметофитической самосовместимости (GSI), как совместимые и несовместимые зерна пыльцы прорастают на стигму. Пыльца зерна несет один из двух S-аллелейоригинального генотипа, в данном случае либо S1 или S2. Если S-аллельпыльцы зерна соответствует одному из двух S-аллелейпестил, в этом случае S1S3, рост пыльцы трубки тормозится в верхней одной трети стиля. (B) Прорастание пыльцы зерна на поверхности стигмы. (C) Пыльца трубки арестован в стиле, указывая на несовместимое поведение. (D) Пыльца трубы растут по стилю. (E) Пыльца трубы в основании стиля, указывающие на совместимое поведение. Масштабные бары, 100 мкм. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой фигуры.

Рисунок 3. Репрезентативные результаты прорастания пыльцы и роста пыльцы трубки через стиль для самосовместимывания и самоотвердивания сортов после самоопыля.
Процент пестилов с пыльцой зерна прорастающих на поверхности стигмы, с пыльцой труб на полпути стиль, в основании стиля, и достижения яйцеклетки. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой фигуры.

Рисунок 4. Усиление фрагмента ПЦР с использованием пяти комбинаций основных пар для идентификации S-аллелей. S
(A) Выход анализатора гена для праймеров SRc-(F/R), показывающих размер двух усиленных фрагментов первой интронации RNase, соответствующей S-аллелям. S (B) усиление ПЦР с использованием грунтовок PruC2/PruC4R для идентификации S6 и S9 аллелей. (C) ПЦР продукты, полученные с использованием конкретных грунтовок SHLM1 и SHLM2 для дифференциации аллеля S1 и SHLM3 и SHLM4, чтобы отличить S7 аллель. (D) УСИЛЕНИЕ ПЦР с aprFBC8-(F/ R) грунтовки для идентификации Sc и S8 аллелей. MI: 1 кб ДНК Лестница. MII: 100 bp ДНК Лестница. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой фигуры.

Рисунок 5. Схема экспериментального дизайна для выяснения отношений совместимости между собой и между собой и между собой в абрикосовых сортах.
() Рабочийпроцесс самостоятельного определения совместимости контролируемых опылихов в лаборатории. (B) Рабочий процесс идентификации S-аллеляс помощью молекулярных подходов. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой фигуры.
| ПЦР Мастер Микс | Термоциклер условия | ||||||
| Компоненты | Окончательная концентрация | Реакция 15 МЛ | Шаг цикла | Температура | Время | Циклов | |
| 10x NH4 Реакция Буфер | 10x | 1,5 л | Первоначальная денатурация | 94 КК | 3 мин. | 1 | |
| 50 mM MgCl2 Решение | 25 мМ | 1,2 л | Денатурирование | 94 КК | 1 мин. | 35 | |
| 100 мМ дНТП | 2,5 мМ | 0,6 л | Отжига | 55 градусов по Цельсию | 1 мин. | ||
| Праймер SRc-F | 10 МКМ | 0,6 л | Расширение | 72 КК | 3 мин. | ||
| Праймер SRc-R | 10 МКМ | 0,6 л | Окончательное расширение | 72 КК | 5 мин. | 1 | |
| 500 U Так ДНК полимераза | 0,5 U | 0,2 л | 4 КК | Держать | |||
| H2O | 8,3 л | ||||||
| Компоненты | Окончательная концентрация | Реакция 25 МЛ | Шаг цикла | Температура | Время | Циклов | |
| 10x буфер ПЦР | 10x | 2,5 л | Первоначальная денатурация | 94 КК | 2 мин | 1 | |
| 5x-решение | 5x | 5 л | Денатурирование | 94 КК | 10 с | 10 | |
| 100 мМ дНТП | 2,5 мМ | 0,5 л | Отжига | 55 градусов по Цельсию | 2 мин | ||
| Праймер ПруК2 | 10 МКМ | 0,2 л | Расширение | 68 градусов по Цельсию | 2 мин | ||
| Праймер C4R | 10 МКМ | 0,2 л | Денатурирование | 94 КК | 10 с | 25 | |
| 250 U Так ДНК полимераза | 10 U | 0,13 л | Отжига | 58 градусов по Цельсию | 2 мин | ||
| H2O | 15,5 л | Расширение | 68 градусов по Цельсию | 2 мин | |||
| Окончательное расширение | 72 КК | 5 мин. | 1 | ||||
| 4 КК | Держать | ||||||
| с 10 с добавил каждый цикл на 68 %C шаг расширения. | |||||||
| Компоненты | Окончательная концентрация | Реакция 25 МЛ | Шаг цикла | Температура | Время | Циклов | |
| 10x буфер ПЦР | 10x | 2,5 л | Первоначальная денатурация | 94 КК | 2 мин | 1 | |
| 5x-решение | 5x | 5 л | Денатурирование | 94 КК | 30 с | 35 | |
| 100 мМ дНТП | 2,5 мМ | 0,5 л | Отжига | 62 КК | 1,5 мин. | ||
| Праймер SHLM1 | 10 МКМ | 0,2 л | Расширение | 72 КК | 2 мин | ||
| Праймер SHLM2 | 10 МКМ | 0,2 л | Окончательное расширение | 72 КК | 5 мин. | 1 | |
| 250 U Так ДНК полимераза | 10 U | 0,13 л | 4 КК | Держать | |||
| H2O | 15,5 л | ||||||
| Компоненты | Окончательная концентрация | Реакция 20 МЛ | Шаг цикла | Температура | Время | Циклов | |
| 5x ПЦР Буфер | 5x | 4 л | Первоначальная денатурация | 98 градусов по Цельсию | 30 с | 1 | |
| dNTP | 2,5 мМ | 1,6 л | Денатурирование | 98 градусов по Цельсию | 10 с | 35 | |
| Праймер SHLM3 | 10 МКМ | 1 л/л | Отжига | 51 кк | 30 с | ||
| Праймер SHLM4 | 10 МКМ | 1 л/л | Расширение | 72 КК | 1 мин. | ||
| 100 U полимераза ДНК | 5 U | 0,2 л | Окончательное расширение | 72 КК | 5 мин. | 1 | |
| H2O | 12,4 л | 4 КК | Держать | ||||
| Компоненты | Окончательная концентрация | Реакция 25 МЛ | Шаг цикла | Температура | Время | Циклов | |
| 10x буфер ПЦР | 10x | 2,5 л | Первоначальная денатурация | 94 КК | 2 мин | 1 | |
| 100 мМ дНТП | 2,5 мМ | 2 л | Денатурирование | 94 КК | 30 с | 35 | |
| Праймер FBC8-F | 10 МКМ | 1 л/л | Отжига | 55 градусов по Цельсию | 1,5 мин. | ||
| Праймер ФБК8-Р | 10 МКМ | 1 л/л | Расширение | 72 КК | 2 мин | ||
| 250 U Так ДНК полимераза | 10 U | 0,125 л | Окончательное расширение | 72 КК | 5 мин. | 1 | |
| H2O | 17,4 л | 4 КК | Держать | ||||
Таблица 1. Условия реакции и езды на велосипеде для различных комбинаций грунтовки, используемых в этом протоколе.
| Грунтовки | Последовательности | Ссылки |
| SRc-F | 5'-CTCGCTTTTTTTGTTTTTGC-3' | 18 |
| SRc-R | 5'-GGCCATTGTTGCACCCCTTG-3' | 18 |
| Пру-С2 | 5'-CTTTGGCCAAGTAATTATTCAAACC-3' | 35 |
| Пру-C4R | 5'-GGATGTGGTACGATTGAAGCG-3' | 35 |
| ШЛМ1-F | 5'-GGTGGAGGATGATAAGGTAG-3' | 17 |
| SHLM2-R | 5'-GGCTGCATAAGGAAGCTGTAGG-3' | 17 |
| SHLM3-F | 5'-ТАТАТКТТАКТТГТГТ-3' | 17 |
| SHLM4-R | 5'-CACTATGATAATGTG-3' | 17 |
| АпрФБК8-Ф | 5'-CATGGAAAAAGCTGACTTATGG-3' | 26 |
| АпрФБК8-Р | 5'-GCCTCTAATGTCATCTACTACT-3' | 26 |
Таблица 2. Праймеры, используемые в этом протоколе, последовательности и справке для характеристики S-генотипав Prunus armeniaca.
| Сорт | SRc-(F/R) (bp) | PruC2/PruC4R (bp) | SHLM1/SHLM2 (bp) | SHLM3/SHLM4 (bp) | AprFBC8-(F/R) (bp) | S-генотип | Группа несовместимости (I.G) |
| Чудо-кроватка8 | 420, 420 | 749, 1386 | S6S9 | Viii | |||
| Волшебная кроватка8 | 334, 420 | 749 | S2S9 | Xx | |||
| Голдтурик8 | 334, 420 | 749 | S2S9 | - | |||
| T06917 | 334, 408 | 650 | S1S2 | Я | |||
| T12017 | 334, 408 | 650 | S1S2 | - | |||
| C-6 | 334, 408 | 413 | S2S7 | Iv | |||
| Купер Кот8 | 274, 408 | 650 | S1S3 | XVIII | |||
| Априкин | 358, 358 | 500 | ScSc c | - | |||
| Бергекот8 | 334, 358 | 500 | S2Sc | - | |||
| Весенний румянец8 | 274, 358 | 150 | S3S8 | XXI |
Таблица 3. S-генотипирование абрикосовых сортов с пятью праймерными парами, используемыми в этом протоколе и групповой уступкой несовместимости. S В таблице показаны различные Sразмеры полимеразной цепной реакции S-аллелей, усиленные с помощью SRc-(F/R), PruC2/PruC4R, SHLM1/ SHLM2, SHLM3/SHLM4 и aprFBC8-(F/R).
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Традиционно, большинство коммерческих абрикосовых европейских сортов были совместимы36. Тем не менее, использование североамериканских самоухотных сортов в качестве родителей в программах разведения в последние десятилетия привело к выпуску все большего числа новых несовместимых сортов с неизвестными требованиями опыления7,8,37. Таким образом, определение отношений между собой и между собой совместимости в абрикосовых сортах становится все более важным. Это подчеркивается в тех районах, где зимнее охлаждение уменьшается, так как высокие из года в год вариации во время цветения предотвращают совпадение в цветения сортов и их пыльцы во многих случаях, особенно в сортах с высоким охлаждением требований38. Методология, описанная в настоящем ею, сочетающая в себе опыление рук, микроскопию и генетический анализ, была очень полезна для определения совместимости себя (в) совместимости каждого сорта и установления его потенциальных сортов опылителей.
Требования к опылиму могут быть определены с помощью полевых экспериментов в условиях сада11,39. Однако экспозиция к внешним факторам, включая метеорологические неблагоприятные условия, может привести к провалу опыления10,что может привести к ошибочным диагнозам самосовместимости. Методология, описанная в настоящем документе, позволяет более точно оценить самоотверпальность путем микроскопии наблюдений за от опыляемых вручную цветов в лабораторно контролируемых условиях, избегая воздействия на окружающую среду. Кроме того, такой подход позволяет анализировать большее количество сортов в год, так как требуется лишь небольшое количество цветов вместо нескольких взрослых деревьев для каждого сорта, которые требуются в полевых экспериментах40.
Отношения несовместимости могут быть установлены, сочетая ручные опылиения и микроскопию14. Тем не менее, опыляние может быть выполнено только в течение короткого периода во время цветения весной, и взрослые деревья возле лаборатории необходимы, так как продолжительность жизни собранных цветов очень коротка. Таким образом, количество несовместимости отношений, которые могут быть проанализированы контролируемых ручных опылиения в каждом сезоне очень низка. Характеристика генов, кодируемых SS-locus, позволила развивать методы на основе ПЦР для генотипирования S-аллелей18,,41. Такой подход Sускоряет идентификацию S-аллелей, так как он не требует цветов, а эксперименты могут быть проведены с любой вегетативной тканью42. Это расширяет период, в течение которого растительный материал, как правило, молодые листья, могут быть собраны43. Кроме того, листья могут быть лиофилизированы или заморожены, так что анализ может быть сделано в любое время года, в отличие от опылиения, которые могут быть сделаны только на свежие цветы во время цветения сезона44. Дополнительным преимуществом является то, что листья могут быть собраны из молодых деревьев еще до вступления цветения возраста, облегчая сбор образцов и скорейшего получения результатов45.
Генетический анализ позволяет лучше дифференцировать аллели самосовместимости, так как он обеспечивает точные результаты усиленных размеров фрагментов21,46. На сегодняшний день, тридцать три S-аллелибыли определены в абрикос12,18,20,21,,22,23,24, что позволило установить 36 групп несовместимости на основе S-генотипа8,9,17,25,26,27., С другой стороны, недостаток этой методологии заключается в том, что различные аллели в одном и том же размере диапазона или мутации могут быть ошибочно определены как один и тот же аллель. Таким образом, Sc и S8 аллели идентичны для последовательности RNase, но 358-bp вставка находится в гене SFB Sc19. Аналогичным образом, первая область интрона аллелей S1 и S7 идентична и неотличима с помощью грунтовок SRc-F/SRc-R. Кроме того, были найдены несколько омологий, таких как S6 и S528 или S20 и S55,и S7, S13 (EF062341) и S4617,потому что некоторые из этих аллелей были частично секвенированы или по сбоям во время усиления ПЦР и, следовательно, необходима дальнейшая работа, чтобы правильно различить их.
ПЦР-анализ и С-RNase секвенирования являются адекватными для установления несовместимости отношений путем идентификации S-аллелейи распределения сортов в их соответствующей группе несовместимости8,17,26,27. Однако эта методология имеет ограничение, препятствующее определению совместимости само-(в)для конкретных сортов абрикоса. Само совместимость (SC) была связана с конкретными S-аллелейв других видов Prunus 47, как миндаль (Sf)48,49 или сладкая вишня (S4')50,51. Тем не менее, в абрикосе, Sc аллель, который был связан с SC21, может быть ошибочно определены как S8, несовместимый аллель19,22, и возможные мутации, не связанные с S локус, как M-locus12,52, совещаясь SC были определены. В последнее время M-локусбыл генотипированный с помощью SSR маркеров12. Поэтому генетическая идентификация ГК для абрикосовых генотипов требует дальнейших исследований и, чтобы избежать ошибок из-за факторов, не связанных с локусом S, в этой работе характеристика само-(в) совместимости была определена также фенотипированием поведения пыльцы труб через пистил самоопылиных цветов.
Описанная в настоящем документе методология сочетания определения самоотборности путем ручного опыления в лабораторных условиях с последующим наблюдением за поведением пыльцевых труб в пистиль контролируемых самоопилениях под флуоресценцией микроскопии и идентификации S-генотипа с помощью анализа ПЦР позволяет установить требования опыления абрикосовых сортов. S Это обеспечивает ценную информацию для производителей и заводчиков, так как это позволяет установить несовместимость отношений между сортами, чтобы выбрать подходящие опылители в разработке новых садов, а также выбрать соответствующих родителей для разработки новых крестов в программах разведения абрикосов.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Авторам нечего раскрывать.
Acknowledgments
Это исследование финансировалось Ministerio de Ciencia, Innovaci'n y Universidades-Европейский фонд регионального развития, Европейский союз (AGL2016-77267-R, и AGL2015-74071-JIN); Национальный институт Инвестигазон-и-Текнология Агрария и Алиментария (RFP2015-00015-00, RTA2017-00003-00); Европейский социальный фонд Гобьерно-де-Арагон, Европейский союз (A12_17R A12_17R, Фонд Биодиверсидад и Агросегуро С.А.
Materials
| Name | Company | Catalog Number | Comments |
| Agarose D1 Low EEO | Conda | 8010.22 | |
| BIOTAQ DNA Polymerase kit | Bioline | BIO-21060 | |
| Bright field microscope | Leica Microsystems | DM2500 | |
| CEQ System Software | Beckman Coulter | ||
| DNeasy Plant Mini Kit | QIAGEN | 69106 | |
| dNTP Set, 4 x 25 µmol | Bioline | BIO-39025 | |
| GenomeLab DNA Size Standard Kit - 400 | Beckman Coulter | 608098 | |
| GenomeLab GeXP Genetic Analysis System | Beckman Coulter | ||
| GenomeLab Separation Buffer | Beckman Coulter | 608012 | |
| GenomeLab Separation Gel LPA-1 | Beckman Coulter | 391438 | |
| HyperLadder 100bp | Bioline | BIO-33029 | |
| HyperLadder 1kb | Bioline | BIO-33025 | |
| Image Analysis System | Leica Microsystems | ||
| Molecular Imager VersaDoc MP 4000 system | Bio-Rad | 170-8640 | |
| NanoDrop One Spectrophotometer | Thermo Fisher Scientific | 13-400-518 | |
| pH-Meter BASIC 20 | Crison | ||
| Phusion High-Fidelity PCR Kit | Thermo Fisher Scientific | F553S | |
| Power Pack P 25 T | Biometra | ||
| Primer Forward | Isogen Life Science | ||
| Primer Reverse | Isogen Life Science | ||
| Quantity One Software | Bio-Rad | ||
| Stereoscopic microscope | Leica Microsystems | MZ-16 | |
| Sub-Cell GT | Bio-Rad | ||
| SYBR Safe DNA Gel Stain | Thermo Fisher Scientific | S33102 | |
| T100 Thermal Cycler | Bio-Rad | 1861096 | |
| Taq DNA Polymerase | QIAGEN | 201203 | |
| Vertical Stand Autoclave | JP Selecta |
References
- Silva, N. F., Goring, D. R.
- Charlesworth, D., Vekemans, X., Castric, V., Glémin, S. Plant self-incompatibility systems: A molecular evolutionary perspective. New phytologist. 168, 61-69 (2005).
- Tao, R., et al. Identification of stylar RNases associated with gametophytic self-incompatibility in almond (Prunus dulcis). Plant and Cell Physiology. 38, 304-311 (1997).
- Ushijima, K., et al. Structural and transcriptional analysis of the self-incompatibility locus of almond: Identification of a pollen-expressed F-box gene with haplotype-specific polymorphism. The Plant cell. 15, 771-781 (2003).
- Bedinger, P. A., Broz, A. K., Tovar-Mendez, A., McClure, B. Pollen-Pistil Interactions and Their Role in Mate Selection. Plant Physiology. 173, 79-90 (2017).
- Guerra, M. E., Rodrigo, J. Japanese plum pollination: A review. Scientia Horticulturae. 197, 674-686 (2015).
- Zhebentyayeva, T., Ledbetter, C., Burgos, L., Llacer, G. Apricot. Fruit Breeding. Badenes, M. L., Byrne, D. , Springer. Boston, MA. 415-458 (2012).
- Herrera, S., Lora, J., Hormaza, J. I., Herrero, M., Rodrigo, J. Optimizing Production in the New Generation of Apricot Cultivars: Self-incompatibility, S-RNase Allele Identification, and Incompatibility Group Assignment. Frontiers in Plant Science. 9, 527 (2018).
- Egea, J., Burgos, L. Detecting Cross-incompatibility of Three North American Apricot Cultivars and Establishing the First Incompatibility Group in Apricot. Journal of the American Society for Horticultural Science. 121, 1002-1005 (1996).
- Rodrigo, J., Herrero, M. Effects of pre-blossom temperatures on flower development and fruit set in apricot. Scientia Horticulturae. 92, 125-135 (2002).
- Julian, C., Herrero, M., Rodrigo, J. Flower bud differentiation and development in fruiting and non-fruiting shoots in relation to fruit set in apricot (Prunus armeniaca). Trees. 24, 833-841 (2010).
- Muñoz-Sanz, J. V., Zuriaga, E., López, I., Badenes, M. L., Romero, C. Self-(in)compatibility in apricot germplasm is controlled by two major loci, S and M. BMC Plant Biology. 17, 82 (2017).
- Burgos, L., Berenguer, T., Egea, J. Self- and Cross-compatibility among Apricot Cultivars. HortScience. 28, 148-150 (1993).
- Rodrigo, J., Herrero, M. Evaluation of pollination as the cause of erratic fruit set in apricot "Moniqui". Journal of Horticultural Science. 71, 801-805 (1996).
- Milatović, D., Nikolić, D., Krška, B. Testing of self-(in)compatibility in apricot cultivars from European breeding programmes. Horticultural Science. 40 (2), 65-71 (2013).
- Milatović, D., Nikolić, D., Fotirić-Aksić, M., Radović, A. Testing of self-(in)compatibility in apricot cultivars using fluorescence microscopy. Acta Scientiarum Polonorum, Hortorum Cultus. 12 (6), 103-113 (2013).
- Herrera, S., Rodrigo, J., Hormaza, J. I., Lora, J. Identification of Self-Incompatibility Alleles by Specific PCR Analysis and S-RNase Sequencing in Apricot. Int J Mol Sci. 19, 3612 (2018).
- Romero, C., et al. Analysis of the S-locus structure in Prunus armeniaca L. Identification of S-haplotype specific S-RNase and F-box genes. Plant Molecular Biology. 56, 145-157 (2004).
- Halász, J., Pedryc, A., Hegedus, A. Origin and dissemination of the pollen-part mutated SC haplotype which confers self-compatibility in apricot (Prunus armeniaca). New Phytologist. 176, 792-803 (2007).
- Halász, J., Hegedus, A., Hermán, R., Stefanovits-Bányai, É, Pedryc, A. New self-incompatibility alleles in apricot (Prunus armeniaca L.) revealed by stylar ribonuclease assay and S-PCR analysis. Euphytica. 145, 57-66 (2005).
- Vilanova, S., Romero, C., Llacer, G., Badenes, M. L., Burgos, L. Identification of Self-(in)compatibility Alleles in Apricot by PCR and Sequence Analysis. Journal of the American Society for Horticultural Science. 130, 893-898 (2005).
- Feng, J., et al. Detection and transcript expression of S-RNase gene associated with self-incompatibility in apricot (Prunus armeniaca L.). Molecular Biology Reports. 33, 215-221 (2006).
- Zhang, L., et al. Identification of self-incompatibility (S-) genotypes of Chinese apricot cultivars. Euphytica. 160, 241-248 (2008).
- Wu, J., et al. Identification of S-haplotype-specific S-RNase and SFB alleles in native Chinese apricot (Prunus armeniaca L). Journal of Horticultural Science and Biotechnology. 84, 645-652 (2009).
- Szabó, Z., Nyéki, J. Blossoming, fructification and combination of apricot varieties. Acta Horticulturae. 293, 295-302 (1991).
- Halász, J., Pedryc, A., Ercisli, S., Yilmaz, K. U., Hegedűs, A. S-genotyping supports the genetic relationships between Turkish and Hungarian apricot germplasm. Journal of the American Society for Horticultural Science. 135, 410-417 (2010).
- Lachkar, A., et al. Identification of self-(in)compatibility S-alleles and new cross-incompatibility groups in Tunisian apricot (Prunus armeniaca L.) cultivars. The Journal of Horticultural Science and Biotechnology. 88, 497-501 (2013).
- Pérez-Pastor, A., Ruiz-Sánchez, M. C., Domingo, R., Torrecillas, A. Growth and phenological stages of Búlida apricot trees in South-East. Agronomie. 24, 93-100 (2004).
- Williams, J. H., Friedman, W. E., Arnold, M. L. Developmental selection within the angiosperm style: using gamete DNA to visualize interspecific pollen competition. Proceedings of the National Academy of Sciences of the United States of America. 96, 9201-9206 (1999).
- Julian, C., Herrero, M., Rodrigo, J. Anther meiosis time is related to winter cold temperatures in apricot (Prunus armeniaca L.). Environmental and Experimental Botany. 100, 20-25 (2014).
- Guerra, M. E., López-Corrales, M., Wünsch, A., Rodrigo, J. Lack of Fruit Set Caused by Ovule Degeneration in Japanese Plum. Journal of the American Society for Horticultural Science. 136 (6), 375-381 (2011).
- Guerra, M. E., Wünsch, A., López-Corrales, M., Rodrigo, J. Flower Emasculation as the Cause for Lack of Fruit Set in Japanese Plum Crosses. Journal of the American Society for Horticultural Science. 135 (6), 556-562 (2010).
- Hormaza, J. I., Pinney, K., Polito, V. S. Correlation in the tolerance to ozone between sporophytes and male gametophytes of several fruit and nut tree species (Rosaceae). Sexual Plant Reproduction. 9, 44-48 (1996).
- Alcaraz, M. L., Hormaza, J. I., Rodrigo, J. Pistil Starch Reserves at Anthesis Correlate with Final Flower Fate in Avocado (Persea americana). PLOS ONE. 8 (10), 78467 (2013).
- Tao, R., et al. Molecular typing of S-alleles through Identification, Characterization and cDNA cloning for S-RNases in Sweet Cherry. Journal of the American Society for Horticultural Science. 124, 224-233 (1999).
- Burgos, L., et al. The self-compatibility trait of the main apricot cultivars and new selections from breeding programmes. Journal of Horticultural Science. 72, 147-154 (1997).
- Hormaza, J. I., Yamane, H., Rodrigo, J. Apricot. Genome Mapping and Molecular Breeding in Plants, Volume 4 Fruits and Nuts. Kole, C. , Springer. New York, NY. 171-187 (2007).
- Benmoussa, H., Ghrab, M., Ben Mimoun, M., Luedeling, E. Chilling and heat requirements for local and foreign almond (Prunus dulcis Mill.) cultivars in a warm Mediterranean location based on 30 years of phenology records. Agricultural and Forest Meteorology. 239, 34-46 (2017).
- Rodrigo, J., Herrero, M., Hormaza, J. I. Pistil traits and flower fate in apricot (Prunus armeniaca). Annals of Applied Biology. 154, 365-375 (2009).
- Williams, R. R. Techniques used in fruit-set experiments. Towards Regulated Cropping. Williams, R. R., Wilson, D. , Grower Books. London. 57-61 (1970).
- Sutherland, B. G., Robbins, T. P., Tobutt, K. R. Primers amplifying a range of Prunus S-alleles. Plant Breeding. 123, 582-584 (2004).
- Murray, M. G., Thompson, W. F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Research. 8, 4321-4325 (1980).
- Porebski, S., Bailey, L. G., Baum, B. R. Modification of a CTAB DNA Extraction Protocol for Plants Containing High Polysaccharide and Polyphenol Components. Plant Molecular Biology Reporter. 15 (1), 8-15 (1997).
- Rogers, S. O., Bendich, A. J. Extraction of DNA from milligram amounts of fresh, herbarium and mummified plant tissues. Plant Molecular Biology. 5 (2), 69-76 (1985).
- Hormaza, J. I. Molecular characterization and similarity relationships among apricot (Prunus armeniaca L.) genotypes using simple sequence repeats. Theoretical and Applied Genetics. 104, 321-328 (2002).
- Sonneveld, T., Tobutt, K. R., Robbins, T. P. Allele-specific PCR detection of sweet cherry self-incompatibility (S) alleles S1 to S16 using consensus and allele-specific primers. Theoretical and Applied Genetics. 107, 1059-1070 (2003).
- Hegedus, A., Lénárt, J., Halász, J. Sexual incompatibility in Rosaceae fruit tree species: molecular interactions and evolutionary dynamics. Biologia Plantarum. 56 (2), 201-209 (2012).
- Fernández i Martí, A., Gradziel, T. M., Socias i Company, R. Methylation of the Sf locus in almond is associated with S-RNase loss of function. Plant Molecular Biology. 86, 681-689 (2014).
- Company, R. S. i, Kodad, O., Martí, A. F. i, Alonso, J. M. Mutations conferring self-compatibility in Prunus species: From deletions and insertions to epigenetic alterations. Scientia Horticulturae. 192, 125-131 (2015).
- Boskovic, R., Tobutt, K. R. Correlation of stylar ribonuclease zymograms with incompatibility alleles in sweet cherry. Euphytica. 90, 245-250 (1996).
- Cachi, A. M., Wünsch, A. S-genotyping of sweet cherry varieties from Spain and S-locus diversity in Europe. Euphytica. 197 (2), 229-236 (2014).
- Zuriaga, E., et al. An S-locus Independent Pollen Factor Confers Self-Compatibility in "Katy" Apricot. PLoS ONE. 8 (1), 53947 (2013).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}