We developed a novel method to generate 3D spheroids with an in vitro heterotypic cell coculture system that mimics the in vivo tumor microenvironment. Fibroblasts are derived from mouse skin fibroblasts. Skin fibroblasts were generated as described above and were characterized as α-SMA+/Vimentin+/FSP-1+ cells. Human metastatic melanoma cells (C8161) were cultured in W489 medium as described32. To visualize and distinguish fibroblasts from tumor cells, fibroblasts and melanoma cells were pretransduced with GFP/lentivirus and DsRed/lentivirus, respectively34,36, before cell coculture.
Figure 1 shows an example of multicellular 3D spheroids formed by coculturing melanoma cells and fibroblasts. Melanoma cells cultured in the absence of fibroblasts did not form typical 3D spheroids, although some melanoma cells form 2D clusters/aggregates with the extended culture. The average size of the spheroids was approximately 170–360 μm in diameter (mean = 275, SD = 37) on day ~5−7. Using time-lapse imaging, we observed that fibroblasts and tumor cells interacted in coculture and started to form 3D spheroids at around 36 h of coculture as shown in Video 1. Time-lapse imaging recorded the dynamic process of cell-cell interaction and the initial phase of spheroid formation at ~4–52 h of coculture. The peak of 3D spheroid formation occurred around day ~5−7. The formed 3D spheroids were composed of fibroblasts and melanoma cells, where the majority (~80%) were tumor cells. Video 2 shows the dynamic process of single cultured melanoma cells (DsRed+/C8161) in the formation of cell cluster/aggregates starting from ~4–52 h in single culture. The formation of 2D clusters/aggregates peaked around day ~7–10. Video 3 and Video 4 show the structures of a 3D spheroid and a 2D tumor cell cluster visualized by confocal microscopy, respectively. The 3D spheroid and the 2D tumor cell clusters were examined by confocal microscopy on day 7 of cell coculture. Video 5 shows that the 3D spheroids were suspended in the culture medium and mobile, while Video 6 shows that the 2D tumor cell cluster was attached to the culture plate and immobile. Suspension in the medium is a feature of 3D spheroids that distinguishes them from 2D clusters. When the medium in the cell culture dish or well is disturbed by either dropping or gently pipetting the culture medium, the suspended 3D spheroids move, whereas 2D cell clusters are immobile. Only a few single dead cells are mobile.
Figure 2A shows an example of this 3D model serving as a unique platform to study tumor-stroma interactions and to elucidate how intracellular Notch1 signaling pathway activity in CAF regulates cancer stem/initiating cells and spheroid formation. Two pairs of fibroblasts (Fb) isolated from the skin of Gain-Of-Function Notch1 (GOFNotch1: Fsp1.Cre+/-;ROSALSL-N1IC+/+) mice versus their counterpart control (GOFctrl : FSP1.Cre-/-;ROSALSL-N1IC+/+) mice and Loss-Of-Function Notch1 (LOFNotch1: Fsp1.Cre+/-;Notch1LoxP/LoxP+/+) mice versus their counterpart control (LOFctrl : FSP1.Cre-/-; Notch1LoxP/LoxP+/+) mice35, respectively. All fibroblasts were transduced by GFP/lentivirus and cocultured with C8161 melanoma cells pretransduced with DsRed/lentivirus. Time-lapse imaging shows that Fb-GOFNotch1 stopped C8161 melanoma cells from forming 3D spheroids compared to Fb-GOFctrl during the first ~4-52 h of cell coculture. In contrast, Fb-LOFNotch1 promoted formation of 3D spheroids by C8161 melanoma cells compared to Fb-LOFctrl. Figure 2B, top, shows representative images of 3D spheroids formed on day 7 of cell coculture with different fibroblasts carrying out varied Notch pathway activities. Figure 2B, bottom, shows the quantitative data on the average size of 3D spheroids formed on day 7 of cell coculture with different fibroblasts carrying varied Notch pathway activities.
Figure 3 shows an example of this 3D model being used to test the drug response of cancer stem/initiating cells. Cancer stem/initiating cells have been shown to be responsible for drug resistance and cancer recurrence. Therefore, evaluating drug response using this 3D model can better reveal a potential drug's clinical efficacy for cancer treatment. The C8161 melanoma cells rely on active MAPK signaling for cell growth and invasion. They also express high levels of CDK4/Kit, but do not carry a BRAF mutation. To test the drug response of cancer stem/initiating cells towards the MAPK inhibitor using this 3D model, we cocultured C8161 melanoma cells and fibroblasts in 24 well plates. PD0325901 (see Table of Materials), a MAPK inhibitor, was prepared in a serial dilution at a range of concentrations from 1 nM, 2.5 nM, 5 nM, 10 nM, and 25 nM. The PD0325901 was added to the cell cocultures when cell mixtures were plated. Untreated cocultured cells were used as control. We evaluated the spheroid-forming ability of the cell cocultures under different drug concentrations and compared it with the untreated control. Figure 3A shows representative images of 3D spheroids formed on day 5 of cell coculture under different drug concentrations. Figure 3B is the quantitative data of the average size per spheroid and numbers of 3D spheroids formed per low power field (LPF x 4) on day 5 of cell coculture under different drug concentrations.

Figure 1: Formation of 3D spheroids and 2D clusters. (A) Representative image of 3D spheroids formed by coculture of human C8161 melanoma cells and mouse skin fibroblasts. The 3D spheroids were photographed on day 7 of the coculture of melanoma cells and fibroblasts. The average sizes of the spheroids was ~170–360 μm in diameter (mean = 275, SD = 37) on day ~5−7. The average number of spheroids was 18–26 (20.5 ± 3.6) per low power field (LPF x4). (B) Representative image of 2D tumor cell clusters formed by single culture of C8161 melanoma cells. The 2D melanoma cell clusters were photographed on day 7 of single culture of melanoma cells. Please click here to view a larger version of this figure.

Figure 2: Elucidation of role of intracellular Notch1 signaling pathway activity in CAF in regulating cancer stem/initiating cells using the 3D spheroid model. (A) Intracellular Notch1 signaling pathway activity in CAF determined the formation of spheroids by melanoma cells in cell coculture. Time-lapse video shows that Fb-GOFNotch1 stopped the formation of 3D spheroids by the C8161 melanoma cells, while Fb-LOFNotch1 promoted the formation of more 3D spheroids by the C8161 melanoma cells during the first 4–52 h of cell coculture. (B) Top: Representative images of 3D spheroids formed on day 7 of cell coculture with different fibroblasts carrying varied Notch pathway functions. Bottom: The quantitative data of the average size (diameter [μm]/spheroid) of the 3D spheroids formed on day 7 of cell coculture with different fibroblasts carrying varied Notch pathway activities. The two-tail student's t-test was used for statistical analysis. Data are expressed as mean ± standard deviation (SD). Please click here to view a larger version of this figure.

Figure 3: Assessment of drug response of cancer stem/initiating cells using the 3D spheroid model. (A) Representative images of 3D spheroids formed on day 5 of cell coculture under different drug concentrations. (B) The quantitative data of the average size (diameter [μm]/spheroid) and the number of 3D spheroids per low power field (LPF x 4) formed on day 5 of cell coculture under different drug concentrations. Quantitative data are expressed as mean ± standard deviation (SD). Please click here to view a larger version of this figure.
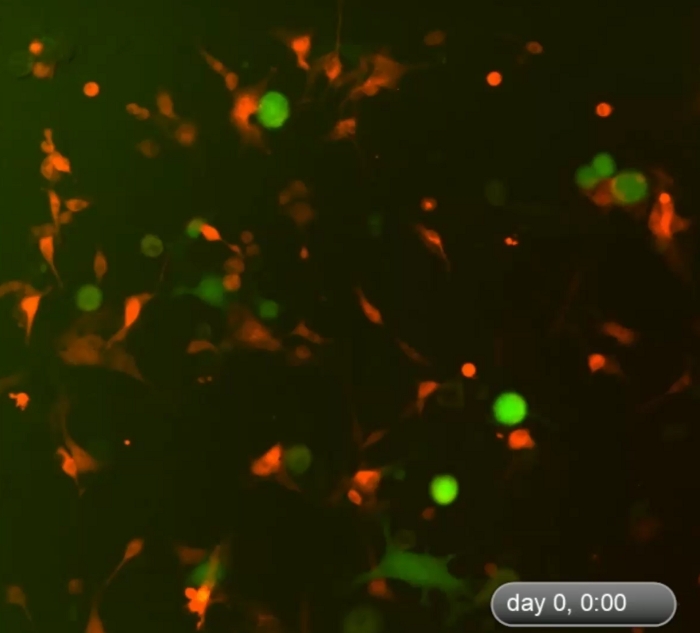
Video 1: Dynamic process of formation of 3D spheroids in the early phase of cell coculture. Time-lapse imaging shows dynamic cell-cell interactions between fibroblasts and tumor cells in the coculture and formation of 3D spheroids during the first ~4–52 h of cell coculture. The cells started to form 3D spheroids around 48 h after the start of coculture. The peak of 3D spheroid formation occurred on day ~5−7 (not displayed here). The 3D spheroids were composed of fibroblasts and melanoma cells, where the majority (~80%) were tumor cells. Please click here to download this video.

Video 2: Dynamic process of formation of 2D clusters in the early phase of cell coculture. Time-lapse imaging shows the dynamic process of 2D clusters formed by C8161melanoma cells in single culture. Formation of 2D clusters occurred around day ~7–10. Time-lapse imaging records the period of ~4–52 h in single culture of DsRed+/C8161 melanoma cells. Please click here to download this video.

Video 3: Architecture and rotation of a 3D spheroid as visualized by confocal microscopy. Architecture and rotation of a 3D spheroid. Green and red lasers were used to scan the spheroids formed on day 7 in cell coculture. The scan area was determined under a 10x objective. The scan starts from the bottom to the top of the spheroid at a 1 µm z-step. The 3D spheroid rotation movie was created using Fiji software. Please click here to download this video.

Video 4: Architecture and rotation a 2D tumor cell cluster as visualized by confocal microscopy. Architecture and rotation of a 2D cluster. Confocal images of cell clusters were taken on day 7 of melanoma cell single culture. The scan area was determined under a 10x objective. The scan starts from the bottom to the top of the spheroid at a 1 µm z-step. The 2D cluster rotation movie was created using Fiji software. Please click here to download this video.

Video 5: Movement of 3D spheroids. The 3D spheroids were suspended in the culture medium and did not adhere to the culture dish/well. When still medium in the cell culture well was disturbed by gentle pipetting, the suspended 3D spheroids moved. Please click here to download this video.

Video 6: Steadfastness of 2D tumor cell clusters. The 2D tumor cell cluster was anchored to the culture plate and immobile in spite of disturbance of the culture medium. A few single dead cells were mobile. Please click here to download this video.
