

ERRATUM NOTICE
Important: There has been an erratum issued for this article. Read more …
Summary
Beskrevet her er en protokoll som muliggjør den kolorimetriske kvantifiseringen av mengden mat spist innen et definert tidsintervall av Drosophila melanogaster larver utsatt for dietter av forskjellig makronæringsstoffkvalitet. Disse analysene utføres i sammenheng med en nevronal termogenetisk skjerm.
Abstract
Foraging og fôringsatferd gjør det mulig for dyr å få tilgang til kilder til energi og næringsstoffer som er avgjørende for deres utvikling, helse og kondisjon. Å undersøke nevronal regulering av disse atferdene er avgjørende for forståelsen av de fysiologiske og molekylære mekanismene som ligger til grunn for ernæringsmessig homeostase. Bruken av genetisk gjennomførbare dyremodeller som ormer, fluer og fisk letter i stor grad denne typen studier. I det siste tiåret har fruktfluen Drosophila melanogaster blitt brukt som en kraftig dyremodell av nevrobiologer som undersøker nevronal kontroll av fôring og foraging atferd. Selv om de utvilsomt er verdifulle, undersøker de fleste studier voksne fluer. Her beskriver vi en protokoll som utnytter det enklere larvenervesystemet for å undersøke nevronale substrater som kontrollerer fôringsatferd når larver blir utsatt for dietter som er forskjellige i protein- og karbohydratinnholdet. Våre metoder er basert på en kvantitativ kolorimetrisk no-choice fôringsanalyse, utført i sammenheng med en nevronal termogenetisk aktiveringsskjerm. Som en avlesning ble mengden mat spist av larver over et 1 h intervall brukt når den ble utsatt for en av de tre fargestoffmerkede diettene som varierer i forholdet mellom protein og karbohydrater (P:C). Effekten av denne protokollen er demonstrert i sammenheng med en nevrogenetisk skjerm i larval Drosophila, ved å identifisere kandidat nevronale populasjoner som regulerer mengden mat spist i dietter av forskjellig makronæringsstoffkvalitet. Vi var også i stand til å klassifisere og gruppere genotypene som ble testet i fenotypiske klasser. I tillegg til en kort gjennomgang av de tilgjengelige metodene i litteraturen, diskuteres fordelene og begrensningene ved disse metodene, og det gis også noen forslag om hvordan denne protokollen kan tilpasses andre spesifikke eksperimenter.
Introduction
Alle dyr er avhengige av et balansert kosthold for å skaffe seg de nødvendige mengdene næringsstoffer for overlevelse, vekst og reproduksjon1. Valget av hva og hvor mye å spise påvirkes av en rekke interagerende faktorer relatert til dyrets indre tilstand, som metthetsnivået og miljøforhold, for eksempel matkvalitet2,3,4,5. Protein og karbohydrater er to store makronæringsstoffer, og det balanserte inntaket er avgjørende for å opprettholde dyrenes fysiologiske prosesser. Derfor er forståelsen av nevrale mekanismer som kontrollerer fôringsatferd og opprettholder et balansert inntak av disse makronæringsstoffene ekstremt relevant. Dette skyldes at livshistorieegenskaper som levetid, fecundity og metabolsk helse påvirkes direkte av nivåene avproteininntaksinntak 6,7,8,9,10.
Bruken av enklere mer gjennomførbare organismer som viser evolusjonært bevarte fôringsvaner med komplekse dyr, inkludert pattedyr, er avgjørende for denne typen studier. Det er viktig at disse enklere dyremodellene gir en god mulighet til å dissekere komplekse biologiske spørsmål i en kostbar, etisk og teknisk mer effektiv sammenheng. I de siste tiårene har Drosophila, med sin kraftige genetiske verktøykasse, intrikate og stereotype oppførsel og bevart arkitektur av perifere og næringssenkende mekanismer med pattedyr, vært en fruktbar modell for atferdsmessige nevrobiologer11. Til syvende og sist er håpet at ved å forstå hvordan matinntaket er regulert i dette dyret, med et enklere nervesystem, kan vi da begynne å løsne nevronale funksjonsfeil som ligger til grunn for menneskelige spiseforstyrrelser.
Studien av nevronale substrater for fôringsatferd er dypt avhengig av å kunne måle dyrenes matinntak samtidig som de manipulerer nevronaktiviteten. På grunn av de minimale mengdene mat som inntas, er kvantifisering av mengden mat spist av fluer ekstremt utfordrende, og alle tilgjengelige metoder gir betydelige begrensninger. Dermed er gullstandarden å bruke en kombinasjon av komplementære metoder12. Voksne fluer har blitt historisk favorisert som en genetisk og atferdsmessig modell. Likevel tilbyr Drosophila larver også muligheter til å undersøke nevronale substrater som koder fôringsadferd. Larval sentralnervesystemet (CNS), med rundt 12.000 nevroner, er betydelig mindre komplekst enn den voksne, som inneholder ca. 150.000 nevroner. Denne lavere kompleksiteten er ikke bare numerisk, men også funksjonell, siden larvatferd er avhengig av enklere lokomotivfunksjoner og sensoriske systemer. Til tross for den tilsynelatende enkelheten i nervesystemet, viser larver fortsatt fullstendig fôringsatferd, og noen metoder for å kvantifisere matinntak i Drosophila larver har blitt beskrevet5,13,14,15. Ved å parre seg med manipulasjoner av nevronaktivitet, kan Drosophila larver utgjøre en svært gjennomførbar modell for å forstå nevral regulering av matinntaket.
Gitt her er en detaljert protokoll for å kvantifisere matinntak i larver utsatt for dietter av forskjellig makronæringsstoffkvalitet. Diettene, såkalte makronæringsstoffbalanseringsdiett, var forskjellig i protein- og karbohydratinnholdet, spesielt med hensyn til forholdet mellom protein og karbohydrat (P:C): 1:1 (proteinrikt kosthold), 1:4 (mellomliggende kosthold) og 1:16 (proteinfattig kosthold), som vist i figur 1A. Kort sagt ble det etablert en kvantitativ no-choice fôringsanalyse ved hjelp av disse tre isocaloric sukrose-gjær (SY)-baserte dietter farget med et blått matfargestoff. Fordi gjærekstrakt og sukrose ble brukt som protein- og karbohydratkilder, og begge inneholder karbohydrater, ble variasjon i P:C-forholdene oppnådd ved å endre balansen mellom disse to komponentene, som tidligere beskrevet16 og som angitt i figur 1B. En skjematisk oversikt over protokollen, som viser de viktigste eksperimentelle trinnene, er tilgjengelig i figur 2.
Denne protokollen ble etablert med sikte på å undersøke rollen til spesifikke nevronpopulasjoner om regulering av larvefôringsnivåer i dietter av forskjellige P: C-forhold og i sammenheng med en termogenetisk nevronskjerm. Et godt karakterisert nevrogenetisk verktøy ble brukt fra TRP-familien (Transient Receptor Potential): Drosophila Transient Receptor Potential channel (dTRPA1), som er en temperatur- og spenningsportert kationkanal, slik at avfyring av handlingspotensialer når omgivelsestemperaturen stiger over 25 °C17. For å uttrykke dTRPA1-transgene, benyttet vi oss av Gal4-linjene basert på cis-regulatoriske regioner fra Drosophila-genomet, etablert i Rubin-laboratoriet, i sammenheng med FlyLight-prosjektet ved Janelia Research Campus18,19.
Selv om protokollen, her beskrevet, er etablert i sammenheng med en aktiveringsskjerm, kan den enkelt tilpasses av eksperimentatoren til andre spesifikke behov eller interesser, nemlig å utføre en undertrykkelsesskjerm ved hjelp av den temperaturfølsomme nevronale lyddemperen ShibireTS20, i tillegg til dTRPA1. Denne og andre tilpasninger diskuteres i protokoll- og diskusjonsseksjonene.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. Tilberedning av sukrose-gjær (SY) dietter
- Vei alle de tørre ingrediensene (agar, gjær, sukrose) for makronæringsstoffbalansering og L3 oppdrett dietter. Mengdene i gram for hver av ingrediensene som trengs for å tilberede 1 liter mat, er angitt i figur 1B.
MERK: Ta hensyn til at ca. 13 ml mat er nødvendig for å fylle en 60 mm Petri-tallerken. - Løs opp alle ingrediensene i sterilt destillert vann (bruk ca. 50% av det totale volumet av vann som trengs for å tilberede maten) og rør mediet i 5-10 min.
- Autoklav i 50 min.
- Etter å ha latt mediene avkjøles, legg til nipagin- og propionsyreløsninger til diettene, ved en endelig konsentrasjon (v / v) på henholdsvis 3% og 0,3%. Til makronæringsstoffet balansere dietter, legg blå mat fargestoff til en endelig konsentrasjon (v / v) på 1%. Fullfør de totale volumene med destillert vann.
- Hell forsiktig mat dietter til 60 mm Petri retter, slik at mengden mat helles er omtrent den samme i hver av platene. Merk platene med P: C-forholdene til diettene.
MERK: Forbered makronæringsstoffet som balanserer dietter på fôringsanalysens dag. Hvis det ikke er mulig, oppbevar de tilberedte diettene ved 4 °C, i en forseglet beholder, i maksimalt 3 dager. Lengre lagringsperioder gjør dietten for tørr og hard, og larvene kan ikke grave inn i mediet.
2. Genetisk kryss av foreldrelinjer
MERK: Bruk Gal4/UAS-systemet21 til å sette opp de genetiske kryssene. I denne protokollen, for å aktivere nevronfunksjon i spesifikke nevronale populasjoner, ble kvinnelige jomfruer av UAS dTRPA1 linje17 brukt og krysset til menn fra Janelia Gal4-linjene (Figur 2A). Den genetiske kontrollen som ble brukt var avkommet til et kryss mellom dTRPA1-linjen og en "tom GAL4" -linje, som bærer Gal4 i vektoren som brukes til å generere Rubin Gal4-samlingen, men uten regulatorisk fragment til stede (attP2)22. For å fremme nevronundertrykkingen kan en UAS-linjekoding ShibireTS20 brukes i stedet for dTRPA1.
- Sett opp 60 mm embryooppsamlingsbur med L3 oppdrettsdiettplater, supplert med litt aktiv gjærpasta.
- Overfør de voksne UAS dTRPA1 kvinnelige jomfruer og Janelia Gal4 menn, i alderen 5-8 dager, til embryo samling bur og la parring skje i 24-48 h, ved 25 °C, med 60% fuktighet og en 12:12 lys-mørk syklus (Figur 2A). For 60 mm embryooppsamlingsbur, bruk rundt 100 jomfruhunner og 30 menn per kors.
- På slutten av parringsperioden, fjern og kast L3 oppdrett diettplater som brukes i de genetiske kryssene. Erstatt dem for ferske L3 oppdrett diettplater, for å utføre egget legger og larval iscenesettelse.
3. Tilberedning av tredje-instar larver (L3)
- Overfør de parrede voksne fluene til friske L3 oppdrettsdiettplater og la eggleggingen skje i 3-4 timer, ved 25 °C (Figur 2B). Pass på at alle platene er merket med genotypen, P: C-forholdet mellom dietten og datoen for egget.
MERK: For å spare tid, utfør eggleggingen direkte inn i L3 oppdrettsdiett, som unngår ekstra håndtering av eggene. Ved småskala genetiske screeninger kan optimalisering av eggleggingen oppnås ved å bruke eplejuice agarplater. - På slutten av eggleggingsperioden, fjern platene fra burene og dekk dem med plastlokk. I tilfelle gjærekstrakt brukes til å supplere L3-oppdrettsplatene, må du sørge for å fjerne all gjenværende gjær i enden av eggleggingen. Dette er viktig for å unngå ikke-jevn fôring under larveveksten.
MERK: De parrede voksne kan overføres til friske L3 oppdrett diettplater, slik at flere egglegg utføres, og mer eksperimentelle larver kan oppnås. Påfølgende egg-lays kan utføres med de samme voksne i løpet av en hel arbeidsuke. - Beregn antall egg per tallerken og hold larvetettheten til maksimalt 200 embryoer per plate. Denne beregningen kan gjøres ved å telle antall embryoer i en fjerdedel av platen.
MERK: En overfylt plate vil forsinke larveutviklingen og påvirke larvefôringsatferd. - Inkuber L3-oppdrettsplatene ved 18 °C (tillatt temperatur), 60 % fuktighet og en lys mørk syklus på 12:12, og la larvene vokse i 9 dager (Figur 2B).
- På den niende dagen etter egglegging (AEL) samler tre grupper på 10 L3 fra hver av genotypene (og for replikeringene) som skal testes. I tillegg samler grupper på 10 L3 for "nullfarget mat" -kontrollen. Forsikre deg om at larvesamlingen gjøres i tilsvarende tidsperioder på dagen som brukes til å gjøre egglegget (f.eks. hvis eggleggingen skjedde mellom 10.00-14.00, samle larvene i samme tidsperiode 9 dager AEL) og utføres, så forsiktig som mulig, ved å bruke tang #5 eller fjærvekt tang. Overfør larvene direkte som angitt i neste trinn (3,6).
MERK: Kontrolldyrene med null fargestoffer er larver som i fôringsanalysen får mat uten blåfarge. Denne kontrollen er viktig for å fjerne bakgrunnsabsorberingen av th larval ekstrakter. - Overfør de oppsamlede eksperimentelle larver til plastfat vekt båter som inneholder 1 ml vann. Kontroller at L3 er samlet inn, og ikke L2, ved å følge instruksjonene i figur 3.
MERK: Samlingen av L3 til plastbåter som inneholder vann eller 1x fosfatbufret saltvann (PBS), er viktig for å holde larver godt hydrert før fôringsanalysen starter. Dette er spesielt viktig hvis flere eksperimentelle L3-grupper fra forskjellige genotyper samles samtidig. Hold oversikt over innsamlingsrekkefølgen for hver gruppe, slik at forskjeller i varigheten av matmangel for hver gruppe minimeres. Bruken av plastbåter i dette trinnet letter det valgfrie trinn 4.3, da det gjør det mulig for larver å flyte direkte inn i vannbadet.
4. Termogenetisk aktivering og ikke-valgbar fôringsanalyse
MERK: Det anbefales å utføre fôringsanalysene på omtrent samme tid på dagen for å minimere mulige variasjoner knyttet til døgnrytmen. Kjør også alltid kontrolleksperimentene (avkommet til den "tomme Gal4" -linjen krysset til UAS dTRPA1 og "nullfarget mat" larver), parallelt med genotypene av interesse.
- Sett opp en inkubator til 30 °C (ikke tillatt temperatur) og hold høye fuktighetsnivåer (minst 65 %) for å unngå larvaldehydrering under analysen.
- Før du starter fôringsanalysen, må du likevekte analyseplatenes temperatur ved å varme dem opp ved 30 °C i 30 minutter.
- (Valgfritt) Varmsjokk de eksperimentelle larver i 2 min i et 37 ° C vannbad. Utfør dette trinnet med dyrene i plastvektbåtene som inneholder litt vann.
MERK: Målet med dette trinnet er å intensivere nevronaktiveringen ved å fremme avfyring av nevronene siden begynnelsen av fôringsanalysen. - Hold flere tidtakere klare til 1 time. Antall tidtakere som skal brukes avhenger av antall eksperimentelle grupper som testes og på eksperimentets ferdighetsnivå på håndtering av larver.
MERK: Bruk av flere tidtakere er avgjørende for å holde analysens varighet konsistent for alle genotypene. - Tøm forsiktig vannet fra plastbåtene, og bruk en fuktet myk børste og overfør L3-gruppene forsiktig fra båtene til midten av analyseplatene. Sett platenes lokk tilbake og start en timer for hver plate (eller gruppe plater) for å opprettholde nøyaktige 1-timers fôringsøkter.
- La larvene mate i 1 time, ved 30 °C, i mørket (Figur 2C).
MERK: Ytelsen til analysen i mørket er viktig for å kontrollere for forskjeller i de visuelle signalene på tvers av dietter, da diettene vil variere i toner, selv om de inneholder samme fargekonsentrasjon. - Stopp fôringsanalysen ved å overføre platene til et isbad. Trykk ned isen så mye som mulig for å gi en stabil overflate for platene.
MERK: Kalde temperaturer vil fremme slutten av fôringen ved å hemme grave- og graveatferd. De fleste larver vil overflate matplatene etter noen minutter, noe som letter utvinningen i de følgende trinnene.
5. Mat fargestoff ekstraksjon
- Forbered 2 ml mikrorør for hver gruppe på 10 L3 testet, som inneholder omtrent de samme mengdene 0,5 mm glassperler (nok til å fylle den nederste delen av mikrorøret) og 300 μL iskald metanol. Hold mikrorørene i kulden ved hjelp av en benkkjøler.
FORSIKTIG: Metanol er svært brannfarlig og giftig. Følg alle sikkerhetsprosedyrer som anbefales for håndtering av dette reagenset, inkludert arbeid i et godt ventilert område og bruk av nitrilhansker.
MERK: Bruk av metanol er viktig for å fikse larveprøvene og unngå melaniseringsreaksjoner i kutikylen. - Bruk #5 eller fjærvekt-tang, forsiktig gjenopprette gruppene på 10 L3 fra fôringsanalyseplatene og overfør dem til lokkene på analyseplatene som inneholder litt vann. Skyll larvene for å fjerne matrester på kroppen mens du forsiktig håndterer larver for å unngå skader. Hold oversikt over antall larver gjenvunnet for hver genotype per replikering, slik at gjennomsnittlig mengde matinntak per larve kan kvantifiseres.
MERK: Skadede larver skal kastes da de vil ha melanisert kutikol, som er uegnet for kolorimetrisk kvantifisering. - Overfør L3-gruppene til 2 ml mikrorør tilberedt i 5.1.
- Lyse larvalvevet for å trekke ut matfargen fra tarmene ved hjelp av en mekanisk lysismetode ved hjelp av en vevlyser og glassperler tilsatt i trinn 5.1. (Hvis en vevslyser ikke er tilgjengelig, bruk en homogeniserende pestle). Fortrinnsvis utfører du dette trinnet ved 4 °C (figur 2D).
MERK: Varigheten av dette trinnet vil avhenge av utstyret som brukes. Ved hjelp av en konvensjonell vevs lyser, 1 min ekstraksjon er tilstrekkelig. I tilfelle tidsbegrensninger, kan protokollen settes på pause på slutten av dette trinnet og fortsette senere. Oppbevar prøvene ved -20 °C. - Overfør ekstraktene for å rengjøre 1,5 ml mikrorør, ved å invertere 2 ml mikrorør direkte til de nye 1,5 ml mikrorørene. Hvis det utføres forsiktig, vil de fleste glassperlene holde seg på bunnen av 2 ml mikrotube.
- Fjern cellulært rusk ved å sentrifugere ekstraktene, med maksimal hastighet i 10 minutter, ved 4 °C.
- Samle supernatantene for å rengjøre 1,5 ml mikrorør. Hvis cellulært rusk fortsatt er synlig i supernatantene, gjentar du trinn 5.6 og 5.7.
6. Kolorimetrisk kvantifisering av matforbruket
- Forbered standardløsninger, for å generere en kalibreringskurve, ved å utføre serielle 1: 2 fortynninger i metanol av en startblå fargestoffløsning. Bruk bare metanol som tom. Konsentrasjonen av standardene er avhengig av dyrenes nivåer av matinntak.
MERK: Når det gjelder pilotskjermen som presenteres her, da fargekonsentrasjonene oppnådd for larveekstraktene varierte fra 0,02 til 1,93 μL / ml, ble den brukt en standardkurve oppnådd ved å måle absorbansene til 8 serielle fortynninger av en 2 μL / ml blåfarget løsning i metanol. Om nødvendig, øke eller redusere konsentrasjonen av disse løsningene, avhengig av fargekonsentrasjonen av eksperimentelle prøver. - Overfør 100 μL av de eksperimentelle prøvene (oppnådd i trinn 5.7), standarder og blanke (trinn 6.1) til brønnene på en 96-brønns mikroplate og mål absorbansen ved 600 nm, ved hjelp av en plateleser (Figur 2E). For å fjerne bakgrunnsabsorberingen, mål absorbansen av ekstrakter hentet fra larver matet på mat uten blåfarget som en "null" for larvalekstraktene ("nullfarget matkontroll).
- Generer en standardkurve og korreler absorbansverdiene som er oppnådd for prøvene fra hver eksperimentell larvalgruppe med mengden matinntak (volum i ml). Finn det gjennomsnittlige matforbruket per larve ved å ta hensyn til antall larver samlet for hver gruppe i trinn 5.2
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Drosophila larver regulerer proteininntaket på bekostning av inntak av overflødige karbohydrater23 (skjematisk tomt i figur 2E). Faktisk har denne prioriteringen av proteininntak blitt observert hos mange andre dyr og kalles proteinet som utnytter24,25.
Ved å dra nytte av denne robuste fôringsadferdsresponsen ble en atferdsbasert skjerm designet med sikte på å identifisere nevronale populasjoner som er involvert i makronæringsstoffbalansering. En no-choice fôringsanalyse ble etablert, som besto av å la grupper på L3 (10 personer per gruppe) mate ad libitum i 1 time og under nevronale termogenetiske aktiveringsbetingelser ved hjelp av dTRPA1, i tre isocaloric (248 Cal/L) matfargede dietter som inneholder spesifikke P:C-forhold (1:1, 1:4 og 1:16) (Figur 1 og Figur 2C). Som en avlesning ble gjennomsnittlig mengde mat spist i makronæringsstoffet dietter av forskjellige P: C-forhold brukt. Ved å dra nytte av Gal4 / UAS-systemet21 og bruke noen av Janelia Gal4-linjene fra FlyLight Project18,19, ble uttrykket av dTRPA1 indusert i spesifikke nevronpopulasjoner.
Med metodene beskrevet i denne protokollen var vi i stand til å kvantifisere den relative mengden makronæringsstoffer som forbrukes, når det gjelder P: C-forhold, for dyr under termogenetisk aktivering av spesifikke nevronale populasjoner i larvalnervesystemet. Denne eksperimentelle tilnærmingen viste at aktivering av distinkte populasjoner av nevroner betydelig påvirket makronæringsstoffbalansering i tredje-instar larver (Figur 4, Tabell 1). Fôringsmønsteret som observeres for kontrollinjen (attP2), viser effektiviteten av metoden ved å vise en forventet kompenserende økning av matinntaket av larver testet i lavere P:C-forholdsdiett (grå prikker og linje i figur 4). Videre ble det funnet et betydelig samspill mellom genotypene og dietten, noe som betyr at termogenetisk aktivering av spesifikke nevronpopulasjoner endrer måten larver regulerer matinntaket på som svar på diettens makronæringsstoffkvalitet.
Fôringsmønstrene til genotypene som er testet i de tre makronæringsstoffbalanseringsdiettene (1:1, 1:4, 1:16), vises av de fargede prikkene og linjene i figur 4, og den statistiske analysen er tilgjengelig i tabell 1.
I aktiveringsskjermen ble totalt 36 Janelia Gal4-linjer kjent for å være sparsomt uttrykt i larvenervesystemet testet. Ved hjelp av lineære regresjonsmodeller bestemte vi hvilke genotyper som viste betydelig forskjellig matinntak med referanse til de genetiske kontrolldyrene. Disse forskjellene inkluderte enten forskjeller i absolutt mengde mat spist på tvers av alle dietter, eller forskjeller i makronæringsstoffets balanserespons (skråning av responsen på de forskjellige P: C-forholdene i diettene).
På tvers av alle tre diettene spiste R12E06 betydelig mer mat enn kontrolldyr. I tillegg overkompenserte det økningen i matinntaket på mellomliggende og lave protein dietter, som indikert av en betydelig forskjell i interaksjonsbegrepet mellom matinntak og P: C-forholdet mellom dietten (Tabell 1). R22H01 spiste betydelig mer enn kontroller, men var ikke forskjellig i makronæringsmiddelbalanseringsresponsen (Tabell 1). R14B11, R19G11, R21B06, R29C02 og R48F09 larver spiste små mengder mat og mistet evnen til å kompensere for den dårlige makronæringsstoffkvaliteten til dietten som er tilgjengelig (som indikert av de betydelige interaksjonsvilkårene mellom matinntak og P: C-forholdet mellom dietten, tabell 1). Til slutt spiste R45D11 larver betydelig mer i det proteinrike kostholdet som inneholder et P:C-forhold på 1:1 enn i mellomliggende og i proteinfattige dietter (1:4 og 1:16), som er det motsatte av hva man forventer på de lave protein diettene.
Derfor tillot metodene våre oss å klassifisere de eksperimentelle larver, fra hver genotype, til fenotypiske klasser relatert til den totale mengden mat spist og evne til å prioritere proteininntak ved å overforbruke i dietter med lavt P:C-forhold. Fem fenotypiske klasser ble etablert for de eksperimentelle dyrene (Figur 5): 1 – "Spis mye" (mer enn kontrolldyrene) og overkompensat for proteinfortynning; 2 – "Spis mye, men kompenser normalt"; 3 – "Spis lite (mindre enn kontrollen), men kompenser"; 4 – "Spis lite og ikke kompenser"; 5 – "Spis avvikende" (mer i proteinrike og mellomliggende dietter enn i proteinfattig diett). I tillegg, for hver av disse fenotypiske klassene og genotypene, viser vi GFP-mønstrene i sentralnervesystemene til tredje-instar larver. Denne informasjonen ble hentet fra de offentlig tilgjengelige bildedataene i FlyLight Project online-plattformen, der man kan få tilgang til uttrykksmønstrene til alle Rubin Gal4-linjene av interesse26.

Figur 1: Sukrose-gjær diettene (SY) som brukes i vår protokoll. (A) De blå prikkene representerer det isocaloriske (248 kalorier/l) makronæringsstoffbalanseringsdiett som brukes i fôringsanalysen, som varierer i forholdet mellom protein og karbohydrat (P:C): 1:1, 1:4 og 1:16. Beige dot representerer dietten som brukes til å bake de eksperimentelle tredje-instar larver (L3), som inneholdt et P: C-forhold på 1:2 og en kaloritetthet på 495 kalorier / L. (B) Detaljert sammensetning og næringsinformasjon om sukrose-gjær (SY) basert dietter. Komponentene er de samme for alle dietter: agar, sukrose og gjær. Mengden i gram av komponentene som trengs for å forberede 1 liter diett, vises. Merk at 1% (v / v) blå fargestoff må legges til makronæringsstoffet balansere dietter og til L3 oppdrett diett nipagin og propionsyre løsninger må legges til en endelig konsentrasjon (v / v) på henholdsvis 3% og 0,3%. Klikk her for å se en større versjon av denne figuren.

Figur 2: Skjematisk fremstilling av hovedtrinnene i vår protokoll (A) Genetisk kryss av foreldrelinjer som utnytter Gal4/UAS-systemet. Krysningen mellom Rubin Gal4-linjene og UAS-linjekodingen dTRPA1 tillater termogenetisk aktivering av spesifikke nevronpopulasjoner i larval sentralnervesystemet. (B) Forberedelse av eksperimentelle tredje-instar larver (L3). Foreldrehunnene fikk lov til å legge egg i 3-4 timer, og larveoppsamlingen skjer ved tillatt temperatur (18 °C) i 9 dager. Valgfritt er varmesjokket ved 37 °C i 2 minutter før fôringsanalysen. (C) Termogenetisk aktivering av nevronfunksjonen og ikke-valgbar fôringsanalyse i 1 time ved ikke-tillatt temperatur (30 °C). Tre grupper på 10 eksperimentelle L3 fra hver genotype fikk lov til å mate i hver av de makronæringsstoffbalanserende diettene som inneholder spesifikke protein- til karbohydrater (P:C) forhold (1:1, 1:4 og 1:16). (D) Mat fargestoff ekstraksjon. Mekanisk lysis av larver, ved hjelp av en vevslys, for å trekke ut det blå matfargen. (E) Tallantifisering av matinntak. Kolorimetrisk kvantifisering av gjennomsnittlig mengde mat spist per larve ved å kvantifisere matfargekonsentrasjonen i larveekstraktene. Absorbansen av eksperimentelle prøver, standarder og "null" ble målt til 600 nm (blå), ved hjelp av en 96-brønns plateleser. Klikk her for å se en større versjon av denne figuren.

Figur 3: Forskjeller mellom andre (L2) og tredje-instars Drosophila larver (L3). L2 og L3 kan lett skilles ved observasjon av spirakler under et stereomikroskop. De fremre spirakler av L2 er klubblignende, mens i L3 er forgrenet. Andre egenskaper kan bidra til å skille de to instars, men er subjektive og mindre pålitelige. De bakre spirakler av L3 har en mørk oransje ring på spissen, som mangler eller er svakt til stede i L2. Luftrøret er tykkere i L3 larver. Illustrasjon: Marisa Oliveira. Klikk her for å se en større versjon av denne figuren.

Figur 4: Mengde mat spist per larve under nevronale termogenetiske aktiveringsforhold i tre makronæringsstoffer som balanserer dietter som inneholder spesifikke protein-til-karbohydrater (P:C)-forhold. Gjennomsnittlige nivåer av mengde mat spist per larve (ml) i 3 makronæringsstoff balansere dietter som inneholder de spesifikke P: C-forholdene på 1: 1, 1: 4 og 1: 16. Grupper på 10 tredje-instar larver, fra hver genotype, fikk lov til å mate i løpet av 1 time, under nevronale termogenetiske aktiveringsforhold, ved hjelp av dTRPA1, ved 30 °C. Genotypene som er testet (larvalavkom fra de genetiske kryssene mellom Rubin Gal4-linjene og UAS dTRPA1-linjen) er indikert av prikker og linjer i forskjellige farger. Som en genetisk kontroll (indikert i grått) ble larvalavkommet fra et kryss mellom den "tomme Gal4" -linjen (attP2) og UAS dTRPA1 brukt. Navnene som ble gitt til genotypene, angitt i forklaringen, var relatert til "Rubin GAL4" -linjene som ble brukt. Klikk her for å se en større versjon av denne figuren.

Figur 5: Gruppere linjene som er testet i 5 hovedfenotypiske klasser. De fenotypiske klassene indikert av tall var basert på kombinasjonen av fenotypene som ble observert når det gjelder total mengde mat spist og evne til å opprettholde proteininntaksprioriteringsresponsen: 1 - spis mye (mer enn kontrolldyrene) og var i stand til å kompensere for proteinfortynning ved å overspise; 2 – spis mye og var ikke i stand til å kompensere; 3 - spis lite (mindre enn kontrollen), men kompenser; 4 – spis lite og var ikke i stand til å kompensere; og 5 - en ekstra fenotypisk klasse, som ble kalt "avvikende", der larvene ikke oppførte seg som forventet som forventet som svar på makronæringsstofffortynning av proteininnhold i kostholdet, og spiste mer i proteinrike og mellomliggende dietter enn i proteinfattig diett. For hver genotype vises GFP-uttrykksmønsteret i sentralnervesystemene til tredje-instar larver. Disse bildedataene for Rubin Gal4-linjene som ble brukt i denne analysen, ble hentet fra den offentlig tilgjengelige FlyLight Project online plattform26. Klikk her for å se en større versjon av denne figuren.
| Anova-tabell (type II-tester) | |||||
| Respons: Konsentrasjon/L3 | |||||
| Sum kv. | Df | F-verdi | Pr(>F) | ||
| mat | 0.086832 | 1 | 113.5358 | < 2,2e-16 | *** |
| Genotype | 0.078443 | 10 | 10.2567 | 9.762e-15 | *** |
| Mat : Genotype | 0.064038 | 10 | 8.3733 | 6.416e-12 | *** |
| Rester | 0.215673 | 282 | |||
| Betydningskoder: 0 ***0,001 **0,01 *0,05. 0.1 ' ' 1 | |||||
| Sammendragstabell (koeffisienter nedenfor sammenlignes med genotypen attP-kontroll): | |||||
| taksere | Std. feil | t verdi | Pr(>|t|) | ||
| (Skjæringspunkt) | 0.064245 | 0.004316 | 14.886 | < 2e-16 | *** |
| mat | -0.058117 | 0.007206 | -8.066 | 2.10e-14 | *** |
| Genotype R12E06 | 0.040243 | 0.008961 | 4.491 | 1.03e-05 | *** |
| Genotype R14B11 | -0.053347 | 0.014361 | -3.715 | 0.000245 | *** |
| Genotype R19G11 | -0.044880 | 0.010788 | -4.160 | 4.23e-05 | *** |
| Genotype R21B06 | -0.051912 | 0.009363 | -5.544 | 6.79e-08 | *** |
| Genotype R22H01 | 0.017682 | 0.007296 | 2.423 | 0.016004 | * |
| Genotype R29C02 | -0.043102 | 0.011113 | -3.879 | 0.000131 | *** |
| Genotype R40D06 | -0.005341 | 0.009876 | -0.541 | 0.589102 | |
| Genotype R45C03 | 0.004064 | 0.009876 | 0.412 | 0.680997 | |
| Genotype R45D11 | -0.052579 | 0.009876 | -5.324 | 2.08e-07 | *** |
| Genotype R48F09 | -0.044612 | 0.011362 | -3.926 | 0.000108 | *** |
| Mat : Genotype R12E06 | -0.037763 | 0.015440 | -2.446 | 0.015067 | * |
| Mat : Genotype R14B11 | 0.058054 | 0.027100 | 2.142 | 0.033031 | * |
| Mat : Genotype R19G11 | 0.051532 | 0.017726 | 2.907 | 0.003937 | ** |
| Mat : Genotype R21B06 | 0.054403 | 0.015689 | 3.467 | 0.000607 | *** |
| Mat : Genotype R22H01 | -0.020863 | 0.012377 | -1.686 | 0.092979 | . |
| Mat : Genotype R29C02 | 0.048996 | 0.018714 | 2.618 | 0.009317 | ** |
| Mat : Genotype R40D06 | 0.003804 | 0.016550 | 0.230 | 0.818371 | |
| Mat : Genotype R45C03 | 0.034117 | 0.016550 | 2.061 | 0.040177 | * |
| Mat : Genotype R45D11 | 0.090661 | 0.016550 | 5.478 | 9.53e-08 | *** |
| Mat : Genotype R48F09 | 0.051184 | 0.019045 | 2.688 | 0.007625 | ** |
| Betydningskoder: 0 ***0,001 **0,01 *0,05. 0.1 ' ' 1 | |||||
| Gjenværende standardfeil: 0,02765 på 282 frihetsgrader | |||||
| Multippel R-kvadrert: 0,516, justert R-kvadrert: 0,4799 | |||||
| F-statistikk: 14,31 på 21 og 282 DF, p-verdi: < 2,2e-16 | |||||
Tabell 1: ANOVA-tabell for effekten av nevronal termogenetisk aktivering og makronæringsstoffkvalitet på dietten som er tilgjengelig på mengden matinntak. En lineær modell ble montert for å bestemme genotypene som viser en fôringsatferd som er vesentlig forskjellig fra kontrolldyrene.
| Genotype | Assosiert gen | opprinnelse | BDSC-lagernummer |
| w[*] ; P{UAS-TrpA1(B). K}attP2 / TM6B, Tb[1] | Bloomington | 26264 | |
| w[1118] ; P{GAL4.1Uw}attP2 | Janelia | 68384 | |
| w[1118] ; P{GMR12E06-GAL4}attP2 | nett (CG11450) | Janelia | Na |
| w[1118] ; P{GMR14B11-GAL4}attP2 / TM3, Sb[1] | dnc (CG32498) | Janelia | 49255 |
| w[1118] ; P{GMR19G11-GAL4}attP2 | CG33696 | Janelia | 48864 |
| w[1118] ; P{GMR21B06-GAL4}attP2 | oa2 (CG6919) | Janelia | 49857 |
| w[1118] ; P{GMR22H01-GAL4}attP2 | fru (CG14307) | Janelia | 49001 |
| w[1118] ; P{GMR29C02-GAL4}attP2 | Ptp69D (CG10975) | Janelia | 48088 |
| w[1118] ; P{GMR40D06-GAL4}attP2 | cnc (CG17894) | Janelia | 48616 |
| w[1118] ; P{GMR45C03-GAL4}attP2 | kni (CG4717) | Janelia | 47936 |
| w[1118] ; P{GMR45D11-GAL4}attP2 | pnt (CG17077) | Janelia | 49563 |
| w[1118] ; P{GMR48F09-GAL4}attP2 | dpr8 (CG32600) | Janelia | 50377 |
Tabell 2: Drosophila linjer som brukes i dette arbeidet. Detaljert informasjon om alle linjene som brukes: kodenavn, genotype, tilknyttet gen, opprinnelse og Bloomington Drosophila Stock Center (BDSC)-nummer.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Med denne protokollen kan man teste larvens evne under termogenetisk aktivering av spesifikke nevronale populasjoner for å regulere inntaksnivåene av protein og karbohydrater, to store makronæringsstoffer, når de blir utsatt for dietter av forskjellig P:C-sammensetning. Denne metoden ble testet i sammenheng med en larval foreløpig screening med sikte på å identifisere nevronale populasjoner knyttet til kontroll av matinntak på tvers av dietter av forskjellig makronæringsstoffkvalitet. Dette arbeidet bidrar også til å demonstrere at Drosophila larver er verdifulle dyremodeller for å undersøke det nevronale grunnlaget for fôringsatferd knyttet til næringshusostase.
I tillegg til informasjonen som er gitt som notater i protokollen, ønsker vi å diskutere noen viktige aspekter ytterligere. Som i enhver atferdsanalyse må det treffes tiltak av eksperimentet for å minimere variasjonen knyttet til dyreadferd. Et svært viktig aspekt som man bør huske på er relatert til viktigheten av å skaffe utviklingssynkroniserte dyr. Bruken av tidlige L3 larver som er godt synkronisert i utviklingsstadiet, vil redusere atferdsvariasjonene som dyrene viser under fôringsanalysen27. Synkronisering av larver oppnås ved korte egglegg og ved å kontrollere tettheten av larver i kulturene. Ikke bruk lengre perioder med egglegging enn de vi angir i protokollen (3-4 timer). Også å kontrollere larvetettheten til maksimalt 200 dyr per plate vil unngå utviklingsforsinkelser og eliminere ytterligere variasjon i fôringsadferd. Vær oppmerksom på at det første egget lå etter parring, må kastes for å opprettholde homogenitet og oppnå bedre synkronisert larvutvikling. Hunnene har befruktet egg i ovidukt og lagt dem i ulike utviklingsstadier, noe som gjør det vanskelig å opprettholde ensartethet blant larvesamling. Det er viktig at minst den første timen eggoppsamlingsplaten kastes før den endelige samlingen. Vær så snill, ta hensyn til at stresset som er forårsaket til dyrene under larvalhåndtering, også kan påvirke atferden negativt. Prøv å være så skånsom som mulig, ved å bruke en myk og vannfuktet børste. Husk til slutt at et høyt antall repliker genererer et mer pålitelig datasett.
Som i enhver eksperimentell protokoll, presenterer metodene våre noen begrensninger. Bruk av en kolorimetrisk metode for å kvantifisere matinntaket basert på opphopning av et matfargestoff i dyrenes tarm innebærer noen forholdsregler knyttet til analysens varighet. For voksne fluer ble det vist at det er en betydelig risiko for å nå en jevn tilstand for fargeakkumulering, der egestionhastigheten tilsvarer inntaket, og reduserer nøyaktigheten av metoden28. Selv om det ikke er bevis for at dette skjer i larver, bestemte vi oss for å utføre en fôringsanalyse med en maksimal varighet på 60 min. Denne varigheten er praktisk og kompatibel med skjerm med høy gjennomstrømning. Hvis du holder den totale varigheten av protokollen så kort som mulig, kan du også fullføre alle trinnene i avsnitt 4, 5 og 6 på en arbeidsdag. Hvis det er nødvendig å endre varigheten av fôringsanalysen, bør analysens varigheter fra 60 til 120 minutter tillate en effektiv kvantifisering av matinntaket på tvers av genotyper, som tidligere vist29. Følsomheten til matdødsmetoder er også relativt lav når små mengder mat forbrukes, noe som reduserer oppløsningen blant genotyper som viser svært lave nivåer av matinntak. Vi setter opp våre fôringsanalyser ved hjelp av et no-choice paradigme. Bare en dietttype er tilgjengelig for hver eksperimentell gruppe larver, som ikke tillater dyr å selvstendig regulere nivåene av proteiner og karbohydrater forbruk. Videre, fordi vi bruker kjemisk udefinerte dietter, er det vanskelig å holde kontroll over næringskonsentrasjoner som direkte kan påvirke mønstrene for larvfôring. For å overvinne disse problemene, eller for å bekrefte og ytterligere dissekere treff som finnes på en foreløpig skjerm, kan det være lurt å vurdere muligheten for å etablere en presis og kontrollert eksperimentell ernæringsmessig kontekst, ved å bruke definerte syntetiske (holidiske) medier10 og sette opp matvalganalyser som tidligere beskrevet30. Når du bruker en protokoll som involverer termogenetisk nevronal modulasjon, er det viktig å vurdere at de nødvendige temperaturskiftene direkte kan påvirke dyrenes atferdsutganger. En komplementær bruk av optogenetiske tilnærminger ville være interessant å kontrollere for temperaturinduserte falske positiver, men bruken av optogenetikk i sammenheng med larvefôringsanalyser er teknisk utfordrende, siden fôringslarver tilbringer mesteparten av tiden gravd i matsubstratet.
Likevel kan flere styrker av vår eksperimentelle tilnærming listes opp. Enkelheten og relativt høy gjennomstrømning av vår metode tillater kvantifisering av matinntak for flere genotyper når de utsettes for forskjellige ernæringsmessige forhold. Fôringsatferd i larvestadiet er lettere kvantifiserbar enn hos voksne fluer, noe som muliggjør generering av bedre funksjonelle avlesninger. Det er også mindre utfordrende å etablere fôringsanalyser som ligner det naturlige miljøet i larver enn det er for voksne, som det tidligere har blitt diskutert31. Videre, sammenlignet med andre tidligere etablerte metoder for å kvantifisere fôring i larver, nemlig de som er basert på manuell telling av antall munnkroksammentrekninger i løpet av en viss periode32, muliggjør vår kolorimetriske metode genetiske screeningstudier på større skalaer. Noen andre metoder er ganske enkelt basert på å score andelen larver med farget mat i tarmene, og tillater ikke en nøyaktig kvantifisering av matinntaksnivåene33,34. Når det gjelder nevrogenetisk kontroll av nevronfunksjon, sikrer det faktum at TRPA1-transgene er inaktiv ved 18 °C at nevronaktivitet ikke påvirkes gjennom larvutvikling. Dette sikrer at den eksperimentelle nevronaktiveringen vil bli utført utelukkende under fôringsanalysen og ikke under larvutviklingen. I tillegg vil vi nevne, en gang til, at vår protokoll lett kan tilpasses eksperimentets spesifikke behov og interesser. For eksempel kan undertrykkelsen av nevronfunksjonen, i stedet for aktivering, lett oppnås ved å erstatte dTRPA1 for en UAS-linje som koder den temperaturfølsomme nevron lyddemperen ShibireTS20. Også, hvis fôringsnivåene utstilt av eksperimentelle larver er svært lave, noe som gjør det vanskelig å kvantifisere matinntaket, er det mulig å utføre et ekstra trinn på 30 min larval sult før fôringsanalysen (før trinnene i avsnitt 4 i protokollen), som tidligere beskrevet15. Dette matmangeltrinnet kan være spesielt interessant hvis du undersøker moduler av sultdrevet oppførsel. Til slutt, i tidligere studier, ved hjelp av kvantitative kolorimetriske metoder, ble det vist at merking av mat med blå fargestoff ikke har noen innflytelse på fôring12. Likevel tror vi at bruk av komplementære, mer nøyaktige og sensitive metoder, som radiomerking av maten12, i mer avanserte stadier av en studie, med sikte på å bekrefte eller ytterligere dissekere treff funnet i foreløpige stadier, ville være et godt supplement til vår metode og bør vurderes av eksperimentatoren. Av alle disse grunnene tror vi på attraktiviteten til metodene våre for å utføre genetiske skjermer (spesielt primære skjermer) med sikte på å identifisere nevronale populasjoner som er involvert i montering av nevronkretser som koder fôringsatferd.
Som et siste notat vil vi nevne det faktum at tusenvis av larval Gal4-linjer etablert i Janelia Research Campus er offentlig tilgjengelige, på Bloomington Drosophila Stock Center og en stor mengde informasjon om larval26 og voksne19 CNS-uttrykksmønstre er også offentlig tilgjengelig på FlyLight Image Database (http://www.janelia.org/gal4-gen1). Disse ressursene gjør det mulig å utarbeide putative strukturfunksjons neuronale kart over nevronene som regulerer fôringsadferd i Drosophila larver. Dette er mulig ved å integrere fenotypisk informasjon generert i nevronskjermer med uttrykksmønstrene til driverne som brukes. Vi mener at metodene våre utgjør en gyldig tilnærming for å generere foreløpige nevronkart for fôringsatferd knyttet til makronæringsstoffbalansering i Drosophila-hjernen.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Forfatterne har ingenting å avsløre.
Acknowledgments
Vi vil gjerne takke Instituto Gulbenkian de Ciência (IGC) for å gi oss tilgang til en del av det eksperimentelle utstyret som er beskrevet i denne protokollen. Dette arbeidet ble støttet av Portuguese Foundation for Science and Technology (FCT), LISBOA-01-0145-FEDER-007660, PTDC/NEU- NMC/2459/2014, IF/00697/2014 og La Caixa HR17-00595 til PMD og av australian Research Council Future Fellowship (FT170100259) til CKM.
Materials
| Name | Company | Catalog Number | Comments |
| 1.5 mL microtubes | Sarstedt AG & Co. | 72.690.001 | |
| 10xPBS | Nytech | MB18201 | |
| 2.0 mL microtubes | Sarstedt AG & Co. | 72.695.500 | |
| 60 mm petri dishes | Greiner Bio-one, Austria | 628161 | |
| 96 well microplates | Santa Cruz Biotechnology | SC-204453 | |
| Agar | Pró-vida, Portugal | ||
| Bench cooler | Nalgene, USA | Labtop Cooler 5115-0032 | |
| Blue food dye | Rayner, Billingshurst, UK | ||
| Cell disruption media | Scientific Industries, Inc. | 888-850-6208 | (0.5 mm glass beads) |
| Dish weight boats | Santa Cruz Biotechnology | SC-201606 | |
| Embryo collection cage for 60 mm petri dishes | Flystuff, Scientific Laboratory Supplies, UK | FLY1212 (59-100) | |
| Featherweight forceps | BioQuip Products, USA | 4750 | |
| Fly food for stocks maintenance | 1 L food contains: 10 g Agar, 100 g Yeast Extract, 50 g Sucrose, 30 mL Nipagin, 3 mL propionic acid | ||
| Forceps #5 | Dumont | 0108-5-PS | Standard tips, INOX, 11cm |
| Incubator | LMS Ltd, UK | Series 2, Model 230 | For thermogenetic feeding assay (30?C) |
| Incubator | Percival Scientific, USA | DR36NL | To stage larvae (19?C) |
| Janelia lines | Janelia Research Campus | Detailed information in Table 2 | |
| Macronutrient balancing diets | Composition and nutritional information in Figure 1 | ||
| Methanol | VWR | CAS number: 67-56-1 | |
| Nipagin (Methyl 4-hydroxybenzoate) | Sigma-Aldrich | H5501 | |
| Nitrile gloves | VWR, USA | ||
| Refrigerated centrifuge | Eppendorf, Germany | 5804 R / Serial number: 5805CI364293 | |
| Rubin Gal4 ines | Janelia Research Campus | Stoks available at Bloomington Drosophila Stock Center | |
| ShibireTS UAS line | Bloomington Drosophila Stock Center | BDSC number: 66600 | Provided by Carlos Ribeiro Group |
| Soft brushes | For sorting anaesthetised fruit flies | ||
| Spectrophotometer plate reader | Thermo Fisher Scientific | Multiskan Go 51119300 | |
| Stereo microscope | Nikon | 1016625 | |
| Sucrose | Sidul, Portugal | ||
| Third-instar larvae (L3) rearing diet | Composition and nutritional information in Figure 1 | ||
| Timer | |||
| Tissue lyzer / bead beater | MP Biomedicals, USA | FastPrep-24 6004500 | |
| TRPA1 UAS line | Bloomington Drosophila Stock Center | BDSC number: 26264 | Expresses TrpA1 under UAS control; may be used to activate neurons experimentally at 25 ?C |
| Water bath | Sheldon Manufacturing Inc., USA | W20M-2 / 03068308 / 9021195 | |
| Yeast extract | Pró-vida, Portugal | 51% Protein, 15% Carbohydrate |
References
- Raubenheimer, D. Nature of nutrition - a unifying framework from animal adaptation to human. , (2012).
- Carvahlo, M. J. a, Mirth, C. K. Coordinating morphology with behavior during development: an integrative approach from a fly perspective. Frontiers in Ecology and Evolution. , (2015).
- Steck, K., et al. Internal amino acid state modulates yeast taste neurons to support protein homeostasis in Drosophila. Elife. 7, 31625 (2018).
- Itskov, P. M., Ribeiro, C. The dilemmas of the gourmet fly: the molecular and neuronal mechanisms of feeding and nutrient decision making in Drosophila. Frontiers in Neuroscience. 7, 12 (2013).
- Bjordal, M., Arquier, N., Kniazeff, J., Pin, J. P., Leopold, P. Sensing of amino acids in a dopaminergic circuitry promotes rejection of an incomplete diet in Drosophila. Cell. 156 (3), 510-521 (2014).
- Grandison, R. C., Piper, M. D., Partridge, L. Amino-acid imbalance explains extension of lifespan by dietary restriction in Drosophila. Nature. 462 (7276), 1061-1064 (2009).
- Lee, K. P., et al. Lifespan and reproduction in Drosophila: New insights from nutritional geometry. Proceedings of the National Academy of Sciences of the United States of America. 105 (7), 2498-2503 (2008).
- Levine, M. E., et al. Low protein intake is associated with a major reduction in IGF-1, cancer, and overall mortality in the 65 and younger but not older population. Cell Metabolism. 19 (3), 407-417 (2014).
- Solon-Biet, S. M., et al. The ratio of macronutrients, not caloric intake, dictates cardiometabolic health, aging, and longevity in ad libitum-fed mice. Cell Metabolism. 19 (3), 418-430 (2014).
- Piper, M. D., et al. A holidic medium for Drosophila melanogaster. Nature Methods. 11 (1), 100-105 (2014).
- Jones, W. D. The expanding reach of the GAL4/UAS system into the behavioral neurobiology of Drosophila. BMB Reports. 42 (11), 705-712 (2009).
- Deshpande, S. A., et al. Quantifying Drosophila food intake: comparative analysis of current methodology. Nature Methods. 11 (5), 535-540 (2014).
- Neckameyer, W. S. A trophic role for serotonin in the development of a simple feeding circuit. Developmental Neuroscience. 32 (3), 217-237 (2010).
- Gasque, G., Conway, S., Huang, J., Rao, Y., Vosshall, L. B. Small molecule drug screening in Drosophila identifies the 5HT2A receptor as a feeding modulation target. Scientific Reports. 3, (2013).
- Schoofs, A., et al. Selection of motor programs for suppressing food intake and inducing locomotion in the Drosophila brain. PLoS Biology. 12 (6), 1001893 (2014).
- Pocas, G. M., Crosbie, A. E., Mirth, C. K. When does diet matter? The roles of larval and adult nutrition in regulating adult size traits in Drosophila melanogaster. Journal of Insect Physiology. , 104051 (2020).
- Hamada, F. N., et al. An internal thermal sensor controlling temperature preference in Drosophila. Nature. 454 (7201), 217-220 (2008).
- Pfeiffer, B. D., et al. Tools for neuroanatomy and neurogenetics in Drosophila. Proceedings of the National Academy of Sciences of the United States of America. 105 (28), 9715-9720 (2008).
- Jenett, A., et al. A GAL4-driver line resource for Drosophila neurobiology. Cell Reports. 2 (4), 991-1001 (2012).
- Kitamoto, T. Conditional modification of behavior in Drosophila by targeted expression of a temperature-sensitive shibire allele in defined neurons. Journal of Neurobiology. 47 (2), 81-92 (2001).
- Brand, A. H., Perrimon, N. Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development. 118 (2), 401-415 (1993).
- Shirangi, T. R., Stern, D. L., Truman, J. W. Motor control of Drosophila courtship song. Cell Reports. 5 (3), 678-686 (2013).
- Mirth, C. M. J. Food intake and food choice are altered by the developmental transition at critical weight in Drosophila melanogaster. Animal Behaviour. 126, 195-208 (2017).
- Simpson, S. J., Raubenheimer, D.
- Raubenheimer, D., Simpson, S. J. Integrative models of nutrient balancing: application to insects and vertebrates. Nutrition Research Reviews. 10 (1), 151-179 (1997).
- Li, H. H., et al. A GAL4 driver resource for developmental and behavioral studies on the larval CNS of Drosophila. Cell Reports. 8 (3), 897-908 (2014).
- Bhatt, P. K., Neckameyer, W. S. Functional analysis of the larval feeding circuit in Drosophila. Journal of Visualized Experiments. (81), e51062 (2013).
- Wong, R., Piper, M. D. W., Blanc, E., Partridge, L. Pitfalls of measuring feeding rate in the fruit fly Drosophila melanogaster. Nature Methods. 5 (3), 214-215 (2008).
- Almeida-Carvalho, M. J., et al. The Ol1mpiad: concordance of behavioural faculties of stage 1 and stage 3 Drosophila larvae. Journal of Experimental Biology. 220, Pt 13 2452-2475 (2017).
- Rodrigues, M. A., et al. Drosophila melanogaster larvae make nutritional choices that minimize developmental time. Journal of Insect Physiology. 81, 69-80 (2015).
- Wong, R., Piper, M. D., Wertheim, B., Partridge, L.
- Wu, Q., et al. Developmental control of foraging and social behavior by the Drosophila neuropeptide Y-like system. Neuron. 39 (1), 147-161 (2003).
- Wu, Q., Zhang, Y., Xu, J., Shen, P. Regulation of hunger-driven behaviors by neural ribosomal S6 kinase in Drosophila. Proceedings of the National Academy of Sciences of the United States of America. 102 (37), 13289-13294 (2005).
- Lingo, P. R., Zhao, Z., Shen, P. Co-regulation of cold-resistant food acquisition by insulin- and neuropeptide Y-like systems in Drosophila melanogaster. Neuroscience. 148 (2), 371-374 (2007).
Tags
Nevrovitenskap Utgave 160 Drosophila larver termogenetisk nevronskjerm larvfôringsadferd makronæringsstoffbalansering protein karbohydrater matinntaksanalyse matfargestoff kolorimetrisk kvantifiseringErratum
Formal Correction: Erratum: Quantification of Macronutrients Intake in a Thermogenetic Neuronal Screen using Drosophila Larvae
Posted by JoVE Editors on 10/06/2020.
Citeable Link.
An erratum was issued for: Quantification of Macronutrients Intake in a Thermogenetic Neuronal Screen using Drosophila Larvae. A figure was updated.
Figure 1 was updated from:
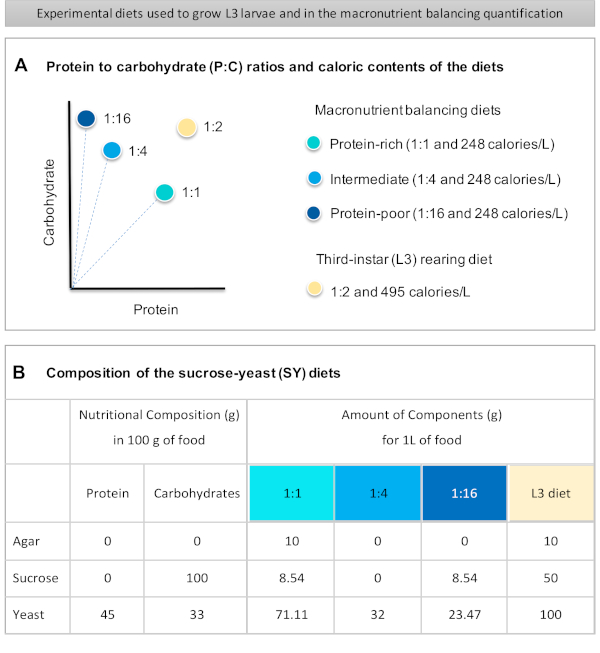
Figure 1: The sucrose-yeast (SY) diets used in our protocol. (A) The blue dots represent the isocaloric (248 calories/L) macronutrient balancing diets used in the feeding assay, which differ in the protein to carbohydrate (P:C) ratios: 1:1, 1:4 and 1:16. The beige dot represents the diet used to rear the experimental third-instar larvae (L3), which contained a P:C ratio of 1:2 and a caloric density of 495 calories/L. (B) Detailed composition and nutritional information of the sucrose-yeast (SY) based diets. The components are the same for all the diets: agar, sucrose and yeast. The amount in grams of the components needed to prepare 1 L of diet is shown. Note that 1% (v/v) of blue dye must be added to the macronutrient balancing diets and to the L3 rearing diet nipagin and propionic acid solutions must be added to a final concentration (v/v) of 3% and 0.3%, respectively. Please click here to view a larger version of this figure.
to:
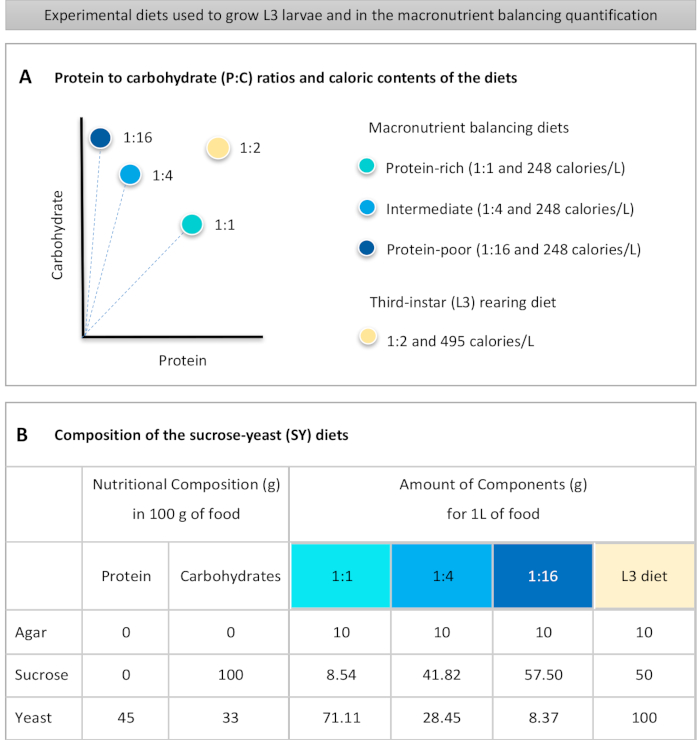
Figure 1: The sucrose-yeast (SY) diets used in our protocol. (A) The blue dots represent the isocaloric (248 calories/L) macronutrient balancing diets used in the feeding assay, which differ in the protein to carbohydrate (P:C) ratios: 1:1, 1:4 and 1:16. The beige dot represents the diet used to rear the experimental third-instar larvae (L3), which contained a P:C ratio of 1:2 and a caloric density of 495 calories/L. (B) Detailed composition and nutritional information of the sucrose-yeast (SY) based diets. The components are the same for all the diets: agar, sucrose and yeast. The amount in grams of the components needed to prepare 1 L of diet is shown. Note that 1% (v/v) of blue dye must be added to the macronutrient balancing diets and to the L3 rearing diet nipagin and propionic acid solutions must be added to a final concentration (v/v) of 3% and 0.3%, respectively. Please click here to view a larger version of this figure.
