The workflow of EM preparation by magnetic separation-assisted high-speed homogenization is shown in Figure 1. Cells internalize 10 nm polylysine-modified IONPs, which are specifically accumulated in endosomes via endocytosis (Figure 3A). After being treated with hypotonic buffer and homogenized, the IONP-loaded endosomes are released from the cells and subsequently collected by magnetic separation. The isolated endosomes are further reconstituted into monodisperse nanovesicles, also known as EMs, by high-speed homogenization. We explored multiple key parameters (e.g., homogenization speed and time) to identify optimized EM preparation conditions (Figure 2). Finally, a homogenization speed of 140 x g for 5 min was chosen as the optimized condition by considering the particle size and yield of produced EMs. Free IONPs and IONP-loaded EMs are eventually removed from the final product solution by a second round of magnetic separation to obtain IONP-free EMs. The method produces highly uniform and monodispersed nanovesicles from parental cell endosomes, sharing the same biological origin as native EVs.
To compare EVs obtained by ultracentrifugation with EMs generated by this method, BMSC and 293T were prepared for EVs and EMs. The diameter and morphology of EMs were analyzed by NTA and TEM. The morphology of BMSC-EMs has the feature of a typical bowl-shaped vesicle-like structure and is delimited by a lipid bilayer (Figure 3A). As analyzed by NTA, both BMSC-EMs and 293T-EMs have a similar hydrodynamic diameter to native EVs (BMSC-EVs and 293T-EVs) (Figure 3B). The BMSC-EMs yields of high-speed homogenization were 8.16 × 108-1.42 × 109/1 × 106cells, and the yield of native EVs prepared by ultracentrifugation was only 7.2 × 107-1.12 × 108/1 × 106cells. Similarly, the 293T-EMs yields of high-speed homogenization were 3.71 × 108-7.58 × 108/1 × 106cells, which reaches up to approximately 100-fold higher than those of native 293T-EVs prepared by the conventional ultracentrifuge method (≈5.5 × 106/1 × 106 cells) (Figure 4A).
Moreover, western blotting results showed that BMSC-EMs contain the same protein biomarkers as EVs (CD63 and Annexin). Both EMs and EVs are negative for Calnexin expression, suggesting that EMs produced by this method had almost no plasma membrane contamination (Figure 3C). There is no significant difference in protein concentration between EM and EV, BMSC-EMs and BMSC-EVs exhibited similar total protein concentrations, 11.15 µg/1 × 109 particles and 14.71 µg/1 × 109 particles via the BCA protein assay kit. Moreover, 293T-EMs and 293T-EVs exhibited total protein concentrations of approximately 31.8 µg/1 × 109 particles and 9.95 µg/1 × 109 particles, respectively (Figure 4B). These results indicate that EMs have a similar protein composition as native EVs. To detect whether EMs can be endocytosed, PKH26-labeled EMs and EVs were co-incubated with BMSC and ID8 at a concentration of 1 × 109 particles/mL for 8 h, and the cells were observed under confocal fluorescence microscopy to confirm that EMs could be readily taken up by the cells for action as well as EVs (Figure 5).

Figure 1: Schematic diagram of the magnetic-assisted high-speed homogenization method. Step 1: Cells internalize IONPs into endosomes through endocytosis. Step 2: Collect organelles, including IONP-loaded endosomes, after hypotonic treatment and homogenization. Step 3: Purify IONP-loaded endosomes by magnetic separation. Step 4: The endosomes are homogenized and reconstituted into monodisperse nanovesicles. Step 5: Remove free IONPs and IONP-loaded EMs by magnetic separation to collect IONP-free EMs. Please click here to view a larger version of this figure.

Figure 2: The optimization of EM preparation conditions. (A) The diameter and PDI of BMSC-EMs in response to homogenization speed changes when time is set at 5 min were analyzed by DLS. (B) The diameter and PDI of BMSC-EMs in response to homogenization time changes when the homogenization speed is set at 140 x g were analyzed by DLS. (C) The diameter and PDI of BMSC-EMs at different storage time points were analyzed by DLS. (D) The diameter and concentration of BMSC-EMs in response to homogenization speed changes when time is set at 5 min were analyzed by NTA. (E) The diameter and concentration of BMSC-EMs in response to the homogenization time change when the homogenization speed is set at 140 x g were analyzed by NTA. (F) The diameter and yield of BMSC-EMs in response to BMSC cells co-incubation with IONPs of different concentrations were analyzed by NTA. ****p < 0.0001. Please click here to view a larger version of this figure.
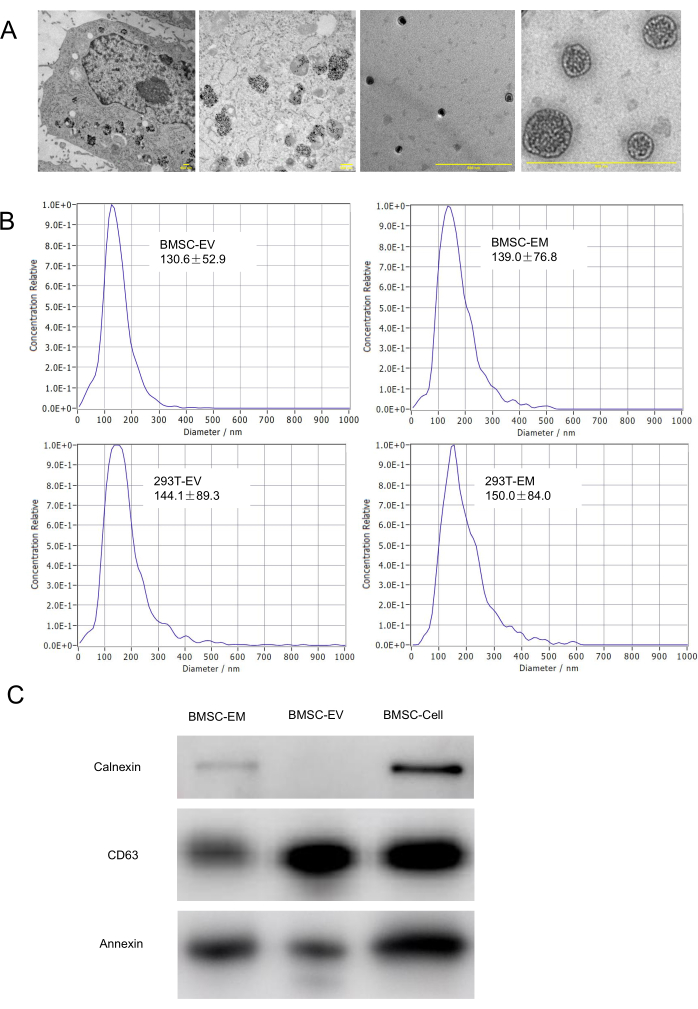
Figure 3: Characterization of EMs. (A) TEM characterization of the morphology of parental cells with endocytosed IONPs, endosomes, and EMs. The scale bars represent 500 nm. (B) Hydrodynamic diameters of BMSC-EM, BMSC-EV, 293T-EM, and 293T-EV are characterized by NTA. (C) Western blot analysis results of BMSC-EMs, BMSC-EVs, and BMSC cell lysates. Please click here to view a larger version of this figure.
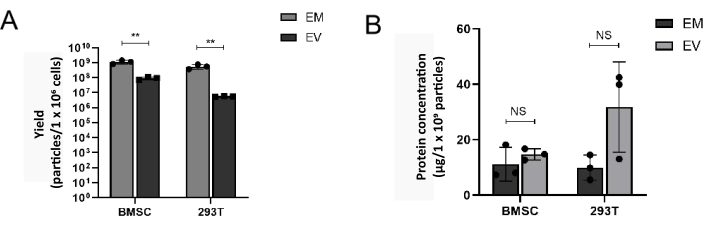
Figure 4: The yield of EMs is higher than EVs. (A) The yield of EMs and EVs prepared from BMSC and 293T was analyzed by NTA. (B) Protein yield of EMs and EVs prepared from BMSC and 293T. **p < 0.01. Please click here to view a larger version of this figure.
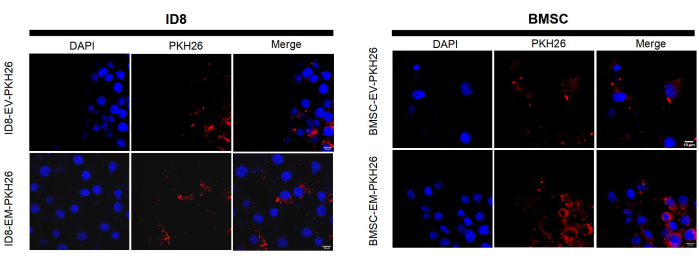
Figure 5: Representative fluorescent microscope images of PKH26-labeled EVs and EMs internalized by BMSC and ID8. The scale bars represent 10 µm. Please click here to view a larger version of this figure.
