


ERRATUM NOTICE
Important: There has been an erratum issued for this article. Read more …
Summary
이 프로토콜은 사용자가 Seahorse 세포외 플럭스 분석을 사용하여 3D 암 세포주 유래 구상체에서 미토콘드리아 에너지 대사를 조사하는 데 도움이됩니다.
Abstract
구상체라고 불리는 3차원 (3D) 세포 응집체는 최근 몇 년 동안 시험관 내 세포 배양의 최전선이되었습니다. 세포를 2차원, 단세포 단층(2D 배양)으로 배양하는 것과는 달리, 스페로이드 세포 배양은 세포외 매트릭스 단백질의 발현, 세포 신호전달, 유전자 발현, 단백질 생산, 분화 및 증식을 포함하여 생체 내에 존재하는 생리적 세포 구조 및 특성을 촉진, 조절 및 지원한다. 3D 배양의 중요성은 종양학, 당뇨병, 줄기 세포 생물학 및 조직 공학을 포함한 많은 연구 분야에서 인정 받고 있습니다. 지난 십 년 동안 구상체를 생산하고 신진 대사 기능과 운명을 평가하기 위해 개선 된 방법이 개발되었습니다.
세포외 플럭스(XF) 분석기는 XF24 섬 포획 플레이트 또는 XFe96 스페로이드 마이크로플레이트를 사용하여 구상체와 같은 3D 미세조직에서 미토콘드리아 기능을 탐색하는 데 사용되었습니다. 그러나, XF 기술을 이용한 구상체에서 미토콘드리아 에너지 대사를 조사하는 뚜렷한 프로토콜과 최적화는 상세히 설명되지 않았다. 이 백서에서는 XFe96 XF 분석기가 있는 스페로이드 마이크로플레이트를 사용하여 단일 3D 구상체에서 미토콘드리아 에너지 대사를 조사하기 위한 상세한 프로토콜을 제공합니다. XF 기술은 서로 다른 암 세포주를 사용하여 크기가 다른 것뿐만 아니라 다른 부피, 세포 번호, DNA 함량 및 유형의 3D 구상체에서 세포 호흡을 구별 할 수있는 것으로 입증되었습니다.
올리고마이신, BAM15, 로테논 및 안티마이신 A의 최적의 미토콘드리아 이펙터 화합물 농도는 3D 구상체에서 미토콘드리아 에너지 대사의 특정 파라미터를 조사하는 데 사용됩니다. 이 백서에서는 또한 구상체에서 얻은 데이터를 정상화하는 방법에 대해 논의하고 XF 기술을 사용하여 스페로이드 대사를 탐구 할 때 고려해야 할 많은 고려 사항을 다룹니다. 이 프로토콜은 고급 시험관 내 스페로이드 모델에 대한 연구를 추진하는 데 도움이됩니다.
Introduction
생물학적 연구에서 시험관 내 모델의 발전은 지난 20 년 동안 빠르게 진행되었습니다. 이러한 모델에는 이제 오르간-온-칩 양식, 오가노이드 및 3D 미세조직 구상체가 포함되며, 이들 모두는 시험관내 및 생체내 연구 사이의 번역을 개선하기 위한 공통적인 초점이 되었다. 고급 시험관 내 모델, 특히 구상체의 사용은 조직 공학, 줄기 세포 연구, 암 및 질병 생물학 1,2,3,4,5,6,7 및 유전 독성 학 8,9,10, 나노 물질 독성학 11을 포함한 안전성 테스트를 포함한 여러 연구 분야에 걸쳐 있습니다. 12,13,14, 및 약물 안전성 및 효능 시험8,15,16,17,18,19.
정상 세포 형태학은 생물학적 표현형과 활성에 매우 중요하다. 세포를 3D 미세조직 구상체로 배양하면 세포가 형태학, 표현형 기능 및 아키텍처를 채택할 수 있으며, 이는 생체 내에서 관찰되는 것과 비슷하지만 고전적인 단층 세포 배양 기술로 포획하기는 어렵습니다. 생체내 및 시험관내 둘 다에서 , 세포 기능은 세포 미세환경에 의해 직접적으로 영향을 받는데, 이는 세포 통신 및 프로그래밍에 한정되지 않는다(예를 들어, 세포-세포 접합 형성, 세포 틈새를 형성할 기회); 즉각적인 환경에서 호르몬 및 성장 인자에 대한 세포 노출 (예를 들어, 염증 반응의 일부로서 세포 사이토카인 노출); 물리적 및 화학적 매트릭스의 조성 (예를 들어, 세포가 뻣뻣한 조직 배양 플라스틱 또는 탄성 조직 환경에서 성장하는지 여부); 그리고 가장 중요한 것은 세포 신진 대사가 영양과 산소에 대한 접근뿐만 아니라 젖산과 같은 대사 폐기물의 처리에 의해 어떻게 영향을 받는지입니다.
대사 플럭스 분석은 정의된 시험관내 시스템 내에서 세포 대사를 검사하는 강력한 방법입니다. 특히, XF 기술은 무손상 세포 및 조직의 세포 생물 에너지학의 실시간 변화를 실시간으로 분석 할 수있게합니다. 많은 세포 내 대사 사건이 수초에서 분 정도 내에 발생한다는 것을 감안할 때, 실시간 기능적 접근법은 시험관 내에서 손상되지 않은 세포 및 조직에서 세포 대사 흐름의 실시간 변화를 이해하는 데 가장 중요합니다.
이 논문은 강제 응집 접근법을 사용하여 시험관내 3D 스페로이드 모델로서 암 유래 세포주 A549 (폐 선암종), HepG2/C3A (간세포 암종), MCF-7 (유방 선암종) 및 SK-OV-3 (난소 선암종)을 배양하기 위한 프로토콜을 제공한다 (그림 1). 또한 (i) 애질런트 XFe96 XF 분석기를 사용하여 단일 3D 구상체의 미토콘드리아 에너지 대사를 조사하는 방법을 자세히 설명하고, (ii) 단일 3D 구상체를 사용하여 XF 분석을 최적화하는 방법을 강조하고, (iii) 이 접근법을 사용하여 3D 스페로이드 대사를 조사하는 데 있어 중요한 고려 사항 및 한계에 대해 논의합니다. 가장 중요한 것은,이 논문은 산소 소모율 (OCR)의 계산이 산화 인산화 및 세포 구상체에서 미토콘드리아 기능을 결정할 수있게 해주는 데이터 세트가 수집되는 방법을 설명합니다. 이 프로토콜에 대해 분석되지는 않았지만 세포외 산성화 속도 (ECAR)는 XF 실험에서 OCR 데이터와 함께 측정되는 또 다른 매개 변수입니다. 그러나 ECAR은 종종 XF 데이터 세트에서 잘못 해석되거나 잘못 해석됩니다. 우리는 기술 제조업체의 기본 접근 방식에 따라 ECAR 계산의 한계에 대한 논평을 제공합니다.
Subscription Required. Please recommend JoVE to your librarian.
Protocol

그림 1: 세포 구상체 생성을 위한 그래픽 워크플로우, 세포외 플럭스 분석 및 다운스트림 분석. 네 개의 암 세포주를 단층(A)으로 선택적으로 배양하고, 조직 배양 플라스크로부터 분리하고, 초저 부착 96-웰 마이크로플레이트에 시딩하여 구상체(B)를 형성하였다. A549 폐 암종, HepG2/C3A 간 암종, SK-OV-3 난소 선암종 및 MCF-7 유방 암종 세포를 1 × 10 3-8 × 103 세포/웰에서 시딩하고 7일까지 성장시켜 단일 구상체를 형성하고 지속적인 관찰 및 평면 측정을 통해 스페로이드 시딩 밀도 및 배양 시간을 최적화하였다. 일단 형성되면, 단일 구상체를 무혈청 XF 배지로 세척하고, 폴리-D-리신 (C)으로 예비코팅된 스페로이드 분석 마이크로플레이트에 조심스럽게 시딩하였다. 스페로이드는 해결하기 위해 몇 가지 프로토콜을 사용하여 XFe96 분석기를 사용하여 세포외 플럭스 분석을 실시하였다: (1) 기저 미토콘드리아 호흡 반응을 위한 최적의 스페로이드 크기; (2) 미토콘드리아 호흡기 억제제의 최적화된 적정; (3) 마이크로플레이트 웰 내의 스페로이드 배치의 최적화. (d) 사후 XF 분석, 위상차 현미경 및 스페로이드 DNA 정량화를 데이터 정규화 및 기타 하류 시험관내 분석에 사용하였다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
1. 암세포주를 3D 체외구 상체로 육성
| 세포주 | 묘사 | 배양 배지 | 근원 |
| A549 | 폐 암종 세포주 | RPMI 1640 | 인증된 세포 배양의 유럽 컬렉션 (ECACC) |
| 피루베이트 나트륨 (1 mM) | |||
| 페니실린 - 스트렙토 마이신 - (100 U / mL - 100 mg / mL) | |||
| 10 % (v / v) FBS | |||
| 헵G2/C3A | 간암종 세포주, 모체 HepG2 세포주의 클론 유도체 | 증권 시세 표시기 | 미국 조직 문화 컬렉션 (ATCC) |
| 페니실린 - 스트렙토 마이신 - (100 U / mL - 100 mg / mL) | |||
| 10 % (v / v) FBS | |||
| MCF7 | 유방 선암종 세포주 | RPMI 1640 | 인증된 세포 배양의 유럽 컬렉션 (ECACC) |
| 피루베이트 나트륨 (1 mM) | |||
| 페니실린 - 스트렙토 마이신 - (100 U / mL - 100 mg / mL) | |||
| 10 % (v / v) FBS | |||
| SK-OV-3 | 난소 선암종 세포주 | RPMI 1640 | 인증된 세포 배양의 유럽 컬렉션 (ECACC) |
| 피루베이트 나트륨 (1 mM) | |||
| 페니실린 - 스트렙토 마이신 - (100 U / mL - 100 mg / mL) | |||
| 10 % (v / v) FBS | |||
| 구성 요소 | RPMI 분석 배지 (최종 부피 50 mL) | ||
| 기본 매체 | 애질런트 해마 XF RPMI, pH 7.4 | ||
| 포도당 (1 M 멸균 스톡) | 11 mM (0.55 mL 원액) | ||
| L-글루타민 (200 mM 멸균 스톡) | 2 mM (0.5 mL의 원액) | ||
| 소듐 피루베이트 (100 mM 멸균 스톡) | 1 mM (0.5 mL의 원액) |
표 1: 암 세포주 배지 및 XF 배지 조성물.
- 표준 무균 조직 배양 기술을 사용하여 모든 세포주를 배양하고 적합한 분석 키트를 사용하여 마이코플라즈마가 없는지 확인합니다.
- 세포주를 T75 조직 배양 플라스크 또는 이와 동등한 권장 배지를 사용하여 배양한다(표 1). 세포주를 65-80% 컨플루언시로 배양하고 최대 25 계대까지 정기적으로 계대시킨다.
- Dulbecco의 변형된 인산염 완충 식염수(DBPS)에서 세포 배양 플라스크를 두 번 헹구십시오.
- 3 mL의 세포 해리 시약( 표 참조)으로 플라스크로부터 세포를 분리하고 37°C에서 5분 동안 현미경으로 분리를 확인하였다.
- 분리된 세포 현탁액을 부드럽게 흡인하여 단일 세포 현탁액을 보장하고, 7 mL의 완전한 조직 배양 배지로 세포 해리 시약을 비활성화시킨다.
- 세포를 300 × g 에서 5분 동안 원심분리하여 수집하고, 상층액을 버리고, 세포를 완전 배지에 재현탁시킨다.
- 혈구세포계 또는 자동화된 세포 계수기를 사용하여 세포를 계수하고 시딩에 필요한 원하는 세포 밀도로 적정한다.
참고: 전체 96웰 플레이트를 100 μL/웰에서 4 × 103 세포/웰에서 시드하려면 세포를 12mL의 권장 부피로 4 × 104 세포/mL로 적정해야 합니다. - 세포 현탁액을 멸균 저장소로 데칸트하고, 100 μL의 세포 현탁액을 다채널 피펫터를 사용하여 세포 기피제 마이크로플레이트의 각 웰에 분배한다.
참고: 마이크로플레이트의 내부 60개 웰만 시드하고 나머지는 DPBS로 채워야 합니다. 이것은 증발 장벽을 형성하고, 플레이트 전체에 걸쳐 스페로이드 균질성을 보장하며, 플레이트 에지 효과를 최소화합니다. - 스페로이드 마이크로플레이트를 300 × g 에서 15분 동안 원심분리하여 세포를 느슨한 응집체 내로 강제한다.
- 플레이트를 스페로이드 형성을 보장하기 위해 최소 3일 동안 37°C, 5%CO2 에서 인큐베이션한다.
- 표준화 된 실험실 관행을 사용하여 위상차 현미경을 수행하여 구상체의 성장을 모니터링하십시오. 세포 배양 배지를 3일마다 또는 매주 두 번 반부피 배지 교환을 수행하여 보충한다.
2. 세포외 플럭스 (XF) 기술을 사용하여 단일 구상체의 미토콘드리아 에너지 대사 조사
- 분석 준비 (하루 전)
- 4x 배율에서 위상 대비가 있는 반전된 광학 현미경을 사용하여 스페로이드 생존력을 확인하여 시료 간의 온전한 스페로이드 구조, 형태학 및 전반적인 균일성을 보장합니다.
- 센서 카트리지에 수분을 공급합니다.
- 교정제 ~20 mL를 원뿔형 튜브에 넣는다.
- 교정제를 함유하는 원뿔형 튜브를CO2 가 아닌 37°C 인큐베이터에 하룻밤 동안 놓는다.
- 분석 키트에서 내용물을 제거합니다.
- 유틸리티 플레이트에서 센서 카트리지를 분리하고 유틸리티 플레이트 옆의 작업대에 거꾸로 놓습니다.
- 200 μL의 멸균ddH2O를 다채널 P300 피펫을 사용하여 센서 카트리지 유틸리티 플레이트의 각 웰에 피펫한다.
- 센서 카트리지를 유틸리티 플레이트 위에 놓습니다.
- 각 웰의 수위가 센서 프로브를 잠수할 만큼 충분히 높은지 확인하십시오.
- 조립된 센서 카트리지를 CO2가 아닌 37°C 인큐베이터로 옮기고 밤새 그대로 둡니다.
참고: 이 단계는 분석 개시 12-72시간 전에 수행될 수 있다.
- 코트 스페로이드 분석 마이크로플레이트
- 무균 기술을 사용하여 30 μL/웰의 멸균 Poly-D-Lysine (0.1 mg/mL) 용액을 스페로이드 마이크로플레이트에 첨가하고 실온에서 30분 동안 배양한다.
- 스페로이드 마이크로플레이트의 각 웰에서 용액을 흡인하고, 플레이트를 반전시킨 다음, 티슈 페이퍼 위에 단단히 두드려 잔류 용액을 제거합니다.
- 200 μL/웰의 멸균 ddH2O로 플레이트를 2회 세척하십시오.
- 최종 세척 후, 마이크로플레이트를 뒤집어 티슈 페이퍼 위에 단단히 두드려 잔류 물을 제거한다.
- 플레이트를 사용하기 전에 30분 동안 공기 건조시키거나 향후 사용을 위해 4°C에서 보관하십시오.
참고: 스페로이드 분석 마이크로플레이트는 스페로이드가 마이크로플레이트의 바닥에 고정되도록 분자 접착제로 코팅되어야 합니다. 분자 접착제가 없으면 구상체가 빠져 나와 분석 결과를 방해 할 수 있습니다. 다른 분자 접착제는 또한 예비코팅 플레이트를 위한 Poly-D-리신에 대한 대안으로서 사용될 수 있다. 예비코팅된 플레이트는 4°C에서 저장될 수 있지만, 분석 개시 전에 실온으로 평형화되도록 방치되어야 한다.
- XF 분석 배지 준비
- 표 1에 상술된 바와 같이 XF RPMI 배지를 준비하고, 0.22 μm 시린지 필터로 멸균 여과한다.
- 분석 준비 (분석 전 1 시간)
- 보충된 XF RPMI 분석 배지를 37°C로 예열한다.
- 코팅된 스페로이드 분석 마이크로플레이트를 비-CO2 37°C 인큐베이터 또는 건식 욕조에서 예열한다.
- 센서 카트리지를 준비합니다.
- 교정제와 센서 카트리지가 들어있는 원뿔형 튜브를 공기 인큐베이터에서 꺼냅니다.
- 유틸리티 플레이트에서 센서 카트리지를 분리하고 작업 표면에 거꾸로 놓습니다.
- P300 다채널 피펫을 사용하여 유틸리티 플레이트에서 물을 흡인하고 버립니다.
- 교정 용액을 멸균 시약 저장소에 붓고 P300 다중 채널 피펫을 사용하여 미리 예열된 교정제 200μL/웰을 유틸리티 플레이트에 추가합니다.
- 센서 카트리지를 집어 들고 유틸리티 플레이트 위에 다시 올려 놓아 센서가 교정기에 잘 잠겨 있는지 확인합니다.
- 조립된 센서 카트리지를 포트 주입 용액을 로드할 준비가 될 때까지 다시 비CO2 37°C 인큐베이터 내로 옮깁니다.
- 구상체를 분석 배지로 세척한다.
- 스페로이드 배양 플레이트를 37°C, 5%CO2 인큐베이터에서 제거하고 스페로이드 전달 단계 전에 스페로이드를 현미경으로 관찰하여 이들의 완전성을 보장한다.
- 스페로이드 플레이트의 모든 웰을 배경 보정 웰을 포함하여 180μL/웰의 예열 분석 매질로 로드합니다.
- 7 cm 페트리 접시를 3 mL의 분석 배지로 부분적으로 채운다.
- 넓은 오리피스 피펫 팁이 로딩된 다채널 피펫을 사용하여, 피펫터를 10-50 μL의 흡인 부피로 설정하여 96-웰 배양 플레이트의 구상체를 7cm 페트리 접시로 옮깁니다.
- 시드 구상체를 예비코팅된 스페로이드 분석 마이크로플레이트에 넣는다.
- 해부 현미경 및 라이트박스 장치를 사용하여, 아래에 상술된 바와 같이 페트리 디쉬로부터 스페로이드 분석 마이크로플레이트로 구상체를 옮긴다.
- 넓은 오리피스 피펫 팁이 장착된 단일 채널 피펫터의 부피를 20μL로 설정하고 단일 구상체를 조심스럽게 흡인합니다. 팁을 스페로이드 분석 마이크로플레이트의 각 웰의 중앙에 직접 배치하고 중력이 단일 스페로이드를 각 웰의 중앙으로 용출시킬 수 있도록 합니다. 즉, 피펫 팁에서 어떤 배지도 배출하지 말고 모세관 작용이 피펫 팁에서 스페로이드를 인출하도록 합니다. 용출을 확인하기 위해, 피펫터의 내용물을 현미경으로 7cm 페트리 접시에 다시 피펫팅할 수 있다.
참고: 단일 스페로이드의 중력 용출은 일반적으로 스페로이드 크기/밀도에 따라 15-30초가 걸립니다. 이 시간 동안 피펫터는 제거해서는 안됩니다. 모든 배경 보정 웰에는 구상체가 없어야하며 분석 배지 만 포함되어야합니다. 현미경으로 각 구상체의 위치를 확인하십시오. 각 스페로이드는 이상적으로 각 웰의 중심 내에 위치해야합니다. - 일단 모든 구상체가 스페로이드 분석 마이크로플레이트로 옮겨지면, 플레이트를 분석 전에 최소 1시간 동안 37°C에서 비CO2 인큐베이터로 옮긴다.
- 넓은 오리피스 피펫 팁이 장착된 단일 채널 피펫터의 부피를 20μL로 설정하고 단일 구상체를 조심스럽게 흡인합니다. 팁을 스페로이드 분석 마이크로플레이트의 각 웰의 중앙에 직접 배치하고 중력이 단일 스페로이드를 각 웰의 중앙으로 용출시킬 수 있도록 합니다. 즉, 피펫 팁에서 어떤 배지도 배출하지 말고 모세관 작용이 피펫 팁에서 스페로이드를 인출하도록 합니다. 용출을 확인하기 위해, 피펫터의 내용물을 현미경으로 7cm 페트리 접시에 다시 피펫팅할 수 있다.
- 해부 현미경 및 라이트박스 장치를 사용하여, 아래에 상술된 바와 같이 페트리 디쉬로부터 스페로이드 분석 마이크로플레이트로 구상체를 옮긴다.
3. XF 분석을위한 센서 카트리지에 화합물 제조 및 로딩
| 주입 전략 | 화합물(포트) | XFe96 마이크로웰 개시 부피(μL) | 원하는 최종 우물 농도 | 포트 부피(μL) | 최종 XFe96 마이크로웰 주입 후 부피(μL) | 작업 재고 집중 |
| 1 | 올리고마이신 (A) | 180 | 3 ug/mL | 20 | 200 | 30 μg/mL |
| 로테논 (B) | 200 | 2 μM | 20 | 220 | 22 μM | |
| 항 마이신 A (B) | 200 | 2 μM | 20 | 220 | 22 μM | |
| 2 | BAM15 (A) | 180 | 5 μM | 20 | 200 | 50 μM |
| 로테논 (B) | 200 | 2 μM | 20 | 220 | 22 μM | |
| 항 마이신 A (B) | 200 | 2 μM | 20 | 220 | 22 μM |
표 2: XFe96 분석기를 사용하여 단일 3D 구상체의 미토콘드리아 에너지 대사를 프로빙하기 위한 미토콘드리아 화합물 농도.
- 완전히 보충된, 예열된 XF RPMI 분석 배지를 사용하여 표 2 에 언급된 바와 같이 각 화합물의 작업 스톡 농도를 준비한다.
- 카트리지 플레이트(유틸리티 플레이트에 연결됨)를 열 방향으로, 1-12 방향을 왼쪽에서 오른쪽으로 맞춥니다.
- 로딩 가이드를 사용하는 경우, 포트 A가 먼저 로딩되는 경우와 같이 잘 로딩된 절차에 따라 카트리지 플레이트 위에 놓으십시오(예: 포트 A가 먼저 로드되는 경우) 가이드의 왼쪽 위 모서리에 A 가 표시되는지 확인하십시오.
- 각 화합물의 작업 용액을 적절한 저장소로 옮기고 보정된 P100 다채널 피펫을 사용하여 20μL를 모든 해당 포트에 분배합니다. 각 컴파운드에 대해 나머지 포트에 대해 반복합니다.
참고: 센서 카트리지 플레이트에 포트를 사용하지 않는 경우 이러한 포트는 비워 두거나 분석 매체로 채울 수 있습니다. 특정 포트 문자의 선택만이 사용되는 경우, 그 문자에 대응하는 다른 포트가 분석 매체로 로딩되도록 보장하십시오; 그렇지 않으면 공기가 우물에 주입되어 우물의 결과를 손상시킵니다. - 포트 로딩 후, 플레이트 로딩 가이드(사용되는 경우)를 제거하고 센서 카트리지를 로딩하기 위한 분석기를 준비합니다.
참고: 포트를로드 한 직후에 분석이 실행되지 않는 경우 센서 카트리지에 뚜껑을 다시 놓고 기계에 적재 할 준비가 될 때까지 플레이트를 37 °C 공기 인큐베이터에 다시 넣으십시오.
4. 분석 설계, 주입 전략 및 데이터 수집
- 분석 실행
- 분석기의 전원을 켜고 컨트롤러(컴퓨터)에 연결합니다.
참고: 이는 웨이브 컨트롤러 소프트웨어의 위젯 패널에 있는 계측기 연결 상태를 통해 확인할 수 있습니다. - WAVE 소프트웨어에서 템플릿 페이지로 이동하여 실험에 대한 분석 템플릿 파일을 찾은 다음 두 번 클릭하여 엽니다.
참고: 분석 템플릿이 템플릿 보기에 나타나지 않으면 템플릿 파일을 공유 네트워크 드라이브 또는 USB 플래시 드라이브에서 템플릿 폴더로 가져옵니다. - 분석을 시작하려면 분석 실행 탭을 클릭합니다.
참고: 그룹 정의가 플레이트 맵 내에 올바르게 할당되면 페이지 오른쪽의 녹색 눈금으로 표시된 대로 분석을 실행할 준비가 됩니다. 이 단계에서, 임의의 추가 정보는 분석 요약 페이지 또는 비워 둔 페이지에 입력될 수 있다; 다음 단계로 진행합니다. 3D 미세조직 구상체에서 미토콘드리아 조절제의 지연된 침투로 인해(그림 2), 표 3에 설명된 측정 프로토콜 정보를 사용하십시오.
- 분석기의 전원을 켜고 컨트롤러(컴퓨터)에 연결합니다.
| 측정 기간 | 주입 수 및 포트 | 측정 세부 정보 | 기간 기간(h:min:s) |
| 교정 | 해당 사항 없음 | XF 분석기는 측정이 정확한지 확인하기 위해 항상 이 보정을 수행합니다. | 00:20:00 (평균이며 기계마다 다를 수 있음) |
| 평형 | 해당 사항 없음 | 평형화는 교정 후에 발생하며 권장됩니다. | 00:10:00 |
| 기저 | 해당 사항 없음 | 사이클 = 5 | 00:30:00 |
| 믹스 = 3:00 | |||
| 대기 = 0:00 | |||
| 측정 = 3:00 | |||
| 올리고마이신/BAM15 | 주입 1(포트 A) | 사이클 = 10 | 01:00:00 |
| 믹스 = 3:00 | |||
| 대기 = 0:00 | |||
| 측정 = 3:00 | |||
| 로테논 + 안티마이신 A | 주입 2(포트 B) | 사이클 = 10 | 01:00:00 |
| 믹스 = 3:00 | |||
| 대기 = 0:00 | |||
| 측정 = 3:00 | |||
| 총 시간: | 03:00:00 | ||
표 3: XFe96 분석기를 사용하여 단일 3D 구상체의 미토콘드리아 에너지 대사를 프로빙하기 위한 프로토콜 설정.
- 실행 시작을 클릭하여 저장 위치 대화 상자를 표시합니다.
- 결과 파일의 저장 위치를 입력하고 조립된 센서 카트리지를 분석기 측면의 도어에서 나타나는 열 트레이에 놓습니다. 열 트레이가 자동으로 열리고 화면에 교정 유틸리티 플레이트 로드 메시지가 표시될 때까지 기다립니다. 화면의 지시를 따르기 전에 i) 유틸리티 플레이트에 센서 카트리지가 제대로 맞는지 확인하고, ii) 센서 카트리지에서 뚜껑을 분리하고, iii) 유틸리티 플레이트에서 센서 카트리지의 올바른 방향을 확인하십시오.
- 화면의 명령에 따라 센서 카트리지 교정을 시작합니다.
참고: 교정을 완료하는 데 걸리는 시간은 약 10-20분입니다(37°C에서의 분석의 경우). - 센서 카트리지 교정 후 웨이브 컨트롤러의 화면 지침에 따라 스페로이드 마이크로플레이트를 분석기에 로드하여 12분 평형 단계를 시작합니다.
참고 : 흰색 눈금이있는 녹색 상자는 그 우물에 대한 '양호한'교정을 나타냅니다. 어떤 우물이 '좋은'교정을 제공하지 못하면 빨간색 상자와 흰색 십자가로 표시됩니다. 이러한 웰은 수정 분석 탭을 사용하여 분석이 완료된 후 임의의 분석으로부터 주목되고 배제되어야 한다 . - 기계가 평형 단계를 완료한 후(계측기 프로토콜에 설명된 대로) 분석기가 기준 측정값을 자동으로 획득하기 시작할 때까지 기다립니다.
- 실험을 완료하려면 WAVE 컨트롤러의 화면에 표시되는 명령을 따릅니다.
참고: 스페로이드 마이크로플레이트가 분석기에서 제거되면 센서 카트리지를 폐기하고 필요한 경우 추가 분석을 위해 스페로이드 플레이트를 따로 두십시오(예: 이중 가닥(ds) DNA 정량화). 마이크로플레이트가 추가 분석을 위해 필요하지 않은 경우, 센서 카트리지와 함께 폐기될 수 있다. - 분석 대화 상자가 나타날 때까지 기다렸다가 결과를 보거나 템플릿 보기로 돌아갑니다.
5. 데이터 정규화 및 분석 전략 - 사후 분석 정규화 및 다운스트림 분석 (선택적 단계)
- 데이터 정규화
- 스페로이드 데이터를 정규화하려면 스페로이드 크기 및 부피를 계산하고 스페로이드 분석에서 dsDNA를 정량화하기 위한 데이터 정규화 전략과 관련된 일련의 프로토콜을 참조하십시오. 이들은 보충 파일로 포함되었습니다. 보충 파일 1 및 보충 파일 2를 참조하십시오.
- 데이터 분석
- 자동화된 분석 생성기 중 하나로 데이터를 내보내려면 WAVE 컨트롤러의 데이터 내보내기 명령에 따라 분석 유형과 일치하는 내보내기 생성기를 선택합니다. 또는 데이터 파일을 내보내고 Seahorse 분석에 업로드합니다.
참고: 보고서 생성기 및 Seahorse 분석의 단점은 데이터 분석이 XF 분석이 설계된 방식으로 제한되며 측정 주기 전반에 걸쳐 평균을 취할 수 없다는 것입니다. 계측기 소프트웨어에서 데이터 세트를 수동으로 내보내면 이와 관련하여 사용자가 선호할 수 있습니다. 3D 구상체의 미토콘드리아 호흡을 평가하기위한 주사 전략이 일반적인 'MitoStress'테스트의 주입 전략과 다를 수 있다는 점을 감안할 때, 일련의 스프레드 시트 템플릿이 개발되어 3D 세포 배양에 특화된 이러한 데이터 세트를 분석하는 데 도움이되며 요청시 제공 될 것입니다. 이들 데이터 템플릿 파일들은 도 2에서 상세하고 설명된 주요 미토콘드리아 호흡 파라미터들에 대한 데이터를 제공할 것이다. - 데이터를 분석하려면 데이터를 WAVE 컨트롤러 소프트웨어에서 스프레드시트 보고서로 내보내고 분석을 위해 독립적인 스프레드시트 템플릿을 사용합니다.
- 자동화된 분석 생성기 중 하나로 데이터를 내보내려면 WAVE 컨트롤러의 데이터 내보내기 명령에 따라 분석 유형과 일치하는 내보내기 생성기를 선택합니다. 또는 데이터 파일을 내보내고 Seahorse 분석에 업로드합니다.

그림 2: 세포외 플럭스 데이터 분석에서 파생된 파라미터에 대한 개략적인 설명자. 약어 : OCR = 산소 소비율. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
잘 형성되고 컴팩트한 구상체를 얻기 위해, 각 세포주는 시딩 밀도 및 배양 기간 동안 개별적으로 최적화되었다(도 3). A549, HepG2/C3A 및 SK-OV-3 세포주는 초기에 느슨한 응집체를 형성하였고, 이는 배양 7일 후까지 명확하게 정의된 둘레를 갖는 둥근 구상체로 진행되지 않았다. 반대로, MCF-7 세포는 3일 이내에 구상체를 형성할 수 있었다. 모든 스페로이드 모델에 대한 배양 기간 후의 초기 세포 시딩 밀도와 스페로이드 부피 사이에는 명확한 상관관계가 있었다. 스페로이드 크기 및 형태학은 시딩 밀도에 최적화되었다. 형태학과 원형성은 모든 모델에서 스페로이드 크기가 증가함에 따라 감소하기 시작했습니다. 세포주에 대한 시딩 전략은 A549 및 SK-OV-3 세포에 대해 4 × 103 세포/웰에서 최적화되었다; HepG2 / C3A 세포는 이전에 다른 곳에서 1 × 10 3 세포 / 웰로 최적화되었으며 MCF-7 세포는 모든 분석에서 4 × 103 세포 / 웰에서 사용되었습니다. 최적화된 시딩 전략에서, 스페로이드 부피는 5.46 × 107 μm3 (SK-OV-3) 및 1.45 × 108 μm3 (A549) 사이였다 (도 3B). 모든 스페로이드 유형은 초기 시딩 밀도와 스페로이드 부피 사이의 선형 상관 관계를 가졌으며, A549 및 HepG2 / C3A는 각각 0.957 및 0.947의R2 값을 가졌습니다. MCF-7 및 SK-OV-3 스페로이드 부피는 모두 초기 시딩 밀도,R2=0.977과 더 큰 상관관계를 갖는 것으로 밝혀졌다(도 3A).
스페로이드 원형도는 길고 짧은 스페로이드 직경을 사용하여 FIJI 분석 소프트웨어 내에서 이미지 평탄도를 사용하여 계산되었습니다. 완벽한 스페로이드 대칭은 원형 = 1.0; 1.0으로부터의 편차는 원형성의 손실을 나타냈다(도 3C). 원형성은 MCF-7 구상체에서 모든 시딩 밀도에서 0.83과 0.9 사이에서 원형이 유지되는 다른 모델보다 컸다. 이에 비해 SK-OV-3 구상체의 외주부는 명확하게 정의되지 않았으며, 배양 7일 후에도 스페로이드 부피가 현저히 작아 4 × 103/웰의 시딩 밀도에서 최대 원형이 0.61인 구상체를 산출하였다. HepG2 / C3A 세포는 또한 모든 구상체의 표면적에 걸쳐 균일 한 형태를 가진 단단하고 잘 형성된 구상체를 형성하는 것으로 밝혀졌으며, 1 × 103 세포 / 웰에 시딩 된 세포에 대해 원형도가 0.79로 유지되었습니다. A549 세포는 스페로이드 원형도 및 형태학이 시딩 밀도로 강화되는 경향을 따르는 것으로 나타났다; 그러나, 원형도는 이들 실험에 사용된 밀도에서 0.63보다 크지 않았다.
기저 미토콘드리아 호흡은 초저 부착 스페로이드 배양 마이크로플레이트에서 1 × 10 3, 2 × 10 3, 또는 8 × 103 세포/웰에 시딩된 구×상체로부터 측정된 OCR로 계산되었다(도 3D). 모든 스페로이드 유형에 대해 OCR은 스페로이드 크기에 따라 증가했으며 스페로이드 부피와 선형적으로 상관 관계가 있었으며 0.988에서 MCF-7 구상체에서R2가 가장 높고 SK-OV-3 구상체가 0.744에서 가장 낮았습니다 (그림 3E). 측정된 OCR은 모든 실험군 간에 통계적으로 상이하였다. A549는 OCR이 가장 낮았고, 가장 큰 스페로이드 크기에서 단지 18pmol/min/well만을 달성했다(그림 3D). 반대로, MCF-7 구상체는 배양 3일 만에 가장 작은 스페로이드 크기에서 유사한 OCR을 산출하여 가장 큰 스페로이드 크기에 대해 53pmol/min/well의 최대 기준선 OCR에 도달했습니다(그림 3D). HepG2 / C3A는 스페로이드 크기 및 형태와 매우 일치하는 OCR 데이터를 산출했습니다. 1 × 10 3 세포/웰에서 시딩된 HepG2/C3A 구상체에서 기준선 OCR은 평균 15 pmol/min/웰에 도달하여 가장 큰 구상체에서 최대 52 pmol/min/웰로 증가했습니다(그림 3D). SK-OV-3 구상체의 OCR은 4 × 10 3 세포/웰 및 8 × 10 3 세포/웰에서 성장한 구상체 사이에서만 유의하였으며, 1 내지 103, 2 × 10 3 또는 4 × 103 세포 사이에서 성장한 구상체에서 거의 차이가 ×지 않았다. 크기 차이에도 불구하고 OCR 데이터는 모든 크기 지점에서 HepG2 / C3A와 MCF-7 구상체 사이에서 매우 유사했습니다. 스페로이드 크기 (μm3)에 비해, MCF-7 구상체에 의한 기준선 OCR은 웰 당 1,000 세포로부터 7 일 동안 성장한 HepG2 / C3A 구상체의 그것과 비교되었다.

그림 3: 스페로이드 성장 파라미터는 기준선 미토콘드리아 호흡을 결정합니다. (A) A549, HepG2/C3A, MCF-7 및 SK-OV-3 구상체는 시딩 밀도에 최적화되었으며 1 × 10 3, 2 × 10 3, 4 × 10 3 및 8 × 10 3 세포 / 웰에서 각각 왼쪽 위에서 오른쪽 하단까지 각 현미경 사진에서 성장을 모니터링했습니다. 스케일 바 = 500 μm. (b) 스페로이드 크기는 수집된 현미경 사진으로부터 평면 데이터를 사용하여 계산되고 피어슨의 상관 통계량을 사용하여 비교되었다; 점선은 95% 신뢰 구간의 분포를 나타냅니다. (c) 스페로이드 형태학은 원형성의 계산에 의해 비교되었다. (d) OCR을 5x로 측정하였고, 그 후 애질런트 시호스 XFe96 분석기를 사용하여 비미토콘드리아 호흡률을 설명하기 위해 로테논-안티마이신 A를 첨가하였다. OCR기저측정은 OCR - OCRr/a로 측정하여 시딩 밀도(D)와 스페로이드 부피(E)를 비교하였다. 데이터는 스페로이드 유형 및 세포 시딩 밀도 당 5-8 웰 반복실험에서 SEM에 ±한 평균이다. 약어: OCR = 산소 소비율; OCR 기저 =기저 미토콘드리아 호흡; OCRr/a = 로테논-안티마이신 A를 첨가한 후 OCR. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
XF 분석에서 호흡 조절제에 노출되기 위한 농도 및 시간 과정은 분석 최적화에서 중요한 단계입니다. 호흡 조절제 화합물-올리고마이신, BAM15, 로테논-안티마이신 A- 또는 DMSO 비히클 대조군의 혼합물을 MCF-7 구상체를 함유하는 마이크로플레이트 웰 내로 센서 카트리지 주입 포트를 통해 순차적으로 주입하였다(도 4A). 모든 샘플 그룹에 대해 30-40 pmol/min/well의 평균 기저 OCR을 결정하기 위해 네 번의 측정 사이클이 완료되었다. 분석 기간의 나머지 기간 동안, 호흡 조절제를 주사 1 후 0.5 μM (0.5 μg/mL 올리고마이신)의 최종 웰 농도를 달성하기 위해 5 측정 사이클마다 순차적으로 첨가하였다; 주사 후 2.0 μM (2 μg/mL 올리고마이신) 2; 주사 후 5 μM (5 μg/mL 올리고마이신) 3; 마지막으로, 네 번째 순차적 첨가 후 11 μM (11 μg/mL 올리고마이신)의 최대 웰 농도. MCF-7 구상체는 실험 전반에 걸쳐 비히클 제어에 반응하지 않았다(도 4A). 기저 OCR은 0.5 μM 또는 0.5 μg/mL 올리고마이신의 최저 농도에서 각 각각의 화합물을 처음 주입한 직후에 변화하였다(도 4B). MCF-7 구상체의 OCR은 0.5 μg/mL의 첫 번째 주사 후 5회 측정 사이클 후에 올리고마이신으로 41 pmol/min/well에서 23 pmol/min/well로 낮아졌다(그림 4B).
0.5 μm BAM15에 반응하여, OCR은 두 번째 주입 전에 33에서 41 pmol/min/웰로 증가하였다(도 4C). 비교적으로, 로테논과 안티마이신 A의 조합은 두 번째 주사 전에 OCR을 37에서 13 pmol/min/well로 낮췄다(그림 4D). 운동 흔적은 OCR에서 꾸준한 선형 감소 (올리고마이신 및 로테논-안티마이신 A) 또는 증가 (BAM15)를 추가로 밝혀냈다. 모든 화합물 투약 요법에 대해, 정상 상태 OCR은 2 μM BAM15, 2 μM 로테논, 2 μM 안티마이신 A 및 2 μg/mL 올리고마이신의 총 웰 농도에서 10-12 완전한 측정 사이클(60-72 분) 내에 달성되었다(도 4A). 산소 소모율은 ~19 pmol/min/well (올리고마이신), 52 pmol/min/well (BAM15) 및 10 pmol/min/well (로테논-안티마이신 A)에서 정상 상태 고원에 도달하였다(그림 4A). 올리고마이신, BAM15, 또는 로테논 플러스 안티마이신 A의 화합물 농도를 증가시키는 것은 OCR에 명백한 영향을 미치지 않았으며, 이는 분석의 나머지 부분에 걸쳐 일정하게 유지되었다. 이 데이터는 3D 구상체를 사용할 때 화합물 농도 및 호흡 조절 화합물에 대한 노출 시간 과정 모두 분석 최적화를 위해 고려되어야한다는 것을 보여줍니다.

도 4: 세포외 플럭스 분석을 최적화하기 위한 중요한 단계로서 호흡 조절제 화합물의 적정. (A) MCF-7 구상체를 4× 10,3 세포/웰에서 시딩하고 XF RPMI를 포함하는 스페로이드 분석 마이크로플레이트의 웰에 배치하기 전에3일 동안 배양하고 XFe96 분석기를 사용하여 OCR ± 미토콘드리아 조절제에 대해 프로브하였다. OCR은 5x를 측정하였고, 그 후 비히클 대조군, 올리고마이신 (B), BAM15 (C), 또는 로테논-안티마이신 A(D)를 첨가하여 미토콘드리아 ATP 합성 효소를 억제하고, 최대 호흡 능력을 결정하거나, 비미토콘드리아 호흡률을 각각 확립하였다. 각 미토콘드리아 조절제의 농도는 최적의 화합물 농도에 대한 반응으로 최대 정상 상태 OCR을 결정하기 위해 4개의 개별 적정 주입 전략(0.5 μM, 1.5 μM, 3 μM 및 6 μM; 올리고마이신의 단위는 μg/mL임)에 걸쳐 증가하였다. OCR은 각 주사 사이의 5 측정 사이클 동안 측정되었다. 데이터는 5-8개의 개별 웰 반복실험으로부터의 SEM± 평균이다. 약어 : OCR = 산소 소비율. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
XF 기술의 주요 이점 중 하나는 손상되지 않은 세포와 조직에서 미토콘드리아 기능을 조사 할 수 있다는 것입니다. 세포 및 조직에서 미토콘드리아 기능의 특정 측면을 조사하기 위해, 미토콘드리아 조절제는 센서 카트리지의 4개의 사용 가능한 주입 포트를 통해 샘플 마이크로플레이트의 웰에 순차적으로 첨가된다. XF 분석에서 미토콘드리아 파라미터를 프로브하는 데 사용되는 조절제의 전형적인 서열은 올리고마이신, 프로토노포어 (예를 들어, FCCP 또는 BAM15) 및 로테논 플러스 안티마이신 A의 조합이며, 이들은 미토콘드리아 ATP 신타제를 억제하고, 최대 호흡 용량을 결정하고, 각각 비미토콘드리아 호흡률에 대해 정확하기 위해 순차적으로 첨가된다. 이러한 조절제 추가 시퀀스는 분석 기술 제조업체에 의해 MitoStress 테스트라고 불립니다. 올리고마이신이 일부 세포 단층(20)에서 비커플러 자극된 호흡을 억제할 수 있다는 것을 감안할 때, 우리는 결합되지 않은 자극된 OCR(OCRmax)을 측정함으로써 암 유래 3D 구상체로 이것을 조사하였다(단일) 및 후(순차적) 올리고마이신 주입(도 5A-D). OCRmax는 HEPG2/C3A 또는 SK-OV-3으로부터 형성된 구상체에 올리고마이신의 첨가에 의해 유의하게 제한되지 않았다(도 5E 및 도 5G). 그러나,OCRmax는 BAM15의 단일 주사로부터 달성된OCRmax에 비해 올리고마이신의 순차적 주사 후 BAM15의 순차적 주사에 이어 A549 및 MCF-7 구상체에서 유의하게 저하되었다(도 5F 및 도 5H). 달리 알려지지 않는 한, 따라서 3D 구상체의 미토콘드리아 에너지 대사를 탐구 할 때 로테논과 항 마이신 A를 최종 첨가하여 올리고 마이신과 언 커플러로 치료하기 위해 별도의 웰을 사용하는 것이 좋습니다. 이 접근법은 여전히 화합물이 순차적으로 첨가되는 전형적인 MitoStress 테스트와 마찬가지로 모든 미토콘드리아 매개 변수의 계산을 허용합니다.

도 5: 미토콘드리아 호흡기 화합물의 단일 또는 순차적 주사. MCF-7, HEPG2/C3A, SK-OV-3 및 A549의 암 세포 유래 구상체를 XF RPMI의 XFe96 스페로이드 마이크로플레이트의 웰에 넣고 애질런트 시호스 XFe96 분석기를 사용하여 OCR에 대해 프로브하였다. OCR은 5x로 측정한 후, 2 μg/mL 올리고마이신 (주입 포트 A: 녹색 트레이스) 또는 5 μM BAM15 (주입 포트 A: 청색 트레이스 또는 주점 포트 B: 녹색 트레이스)를 측정하여 미토콘드리아 ATP 신타제를 억제하고 최대 호흡 용량을 각각 결정하였다. 동역학적 OCR 데이터는 % 기저(A-D)로서 표현된다. 최대 호흡 용량(OCRmax)은 식에 의해 기저 OCR의 인자로서 계산되었다:OCRmax = OCRBAM15/OCR기저. OCRmax는 BAM15 주사 후 8-10 (녹색 막대) 및 (청색 막대) 올리고마이신이 없는 (파란색 막대) 측정 사이클에 걸친 OCR 평균으로부터 얻어졌다. 데이터는 스페로이드 분석 마이크로플레이트± 걸쳐 3-8개의 개별 웰 반복실험으로부터의 SEM에 대한 평균이다. 약어 : OCR = 산소 소비율. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
이러한 최적화 실험에서 결정된 최적의 세포 시딩 밀도, 화합물 농도, 주입 전략 및 측정 주기 주기(표 3)를 사용하여 기저 미토콘드리아 호흡을 정확하게 조사하기 위한 상세한 프로토콜을 개발했습니다: OCR기저(그림 6A), ADP 인산화 호흡: OCRADP(그림 6B), 누출 호흡: OCR오미(그림 6C), 결합 효율(그림 6D ), 최대 호흡 용량 : OCR최대 (그림 6E) 및 예비 호흡 용량 : 암 유래 3D 구상체를 사용하는 OCR예비 (그림 6F).

그림 6: 암 유래 구상체의 미토콘드리아 에너지 대사를 확립하기 위한 XF 기술로 OCR을 조사함. MCF-7, HEPG2/C3A, SK-OV-3 및 A549의 암 세포 유래 구상체를 XF RPMI의 스페로이드 분석 마이크로플레이트의 웰에 배치하고 애질런트 시호스 XFe96 분석기를 사용하여 OCR에 대해 프로브하였다. OCR은 5x로 측정한 후, 2 μg/mL 올리고마이신 또는 5 μM BAM15를 첨가하고, RA를 첨가하여 미토콘드리아 ATP 합성 효소를 억제하고, 최대 호흡 능력을 결정하고, 비미토콘드리아 호흡률을 각각 계산하였다. (a) 기저 미토콘드리아 호흡(OCR기저)은 포트 A 주입 전의 3 측정 사이클로부터의 OCR의 평균으로서 계산되었다. (b) 산화적 인산화의 커플링 효율은 OCR 기저의 백분율로서 OCRADP (OCR기저-OCR누출)를 발현시킴으로써 근사화되었다. (c) ADP 인산화 호흡 (OCRADP)은 BAM15 주사 전에 측정 사이클 11-13에 걸쳐 평균화된 OCR로부터 계산된 올리고마이신 민감성 OCR로서 측정되었다. (d) 오미노미 리크스(OCRleak)는 올리고마이신에 민감하지 않은 OCR로서 측정되었고, 측정 사이클 11-13에 걸친 평균 평균 OCR로부터 계산되었다. (e) 최대 호흡 용량(OCRmax)은 BAM15 주사 후 측정된 평균 최대 OCR로서 측정되었다. (f) 예비 호흡 용량은 OCR 기저의 백분율로서 OCR스페어(OCRmax-OCR 기저)를 발현시킴으로써 계산하였다. 로테논-안티마이신 A 주사 후 OCR (OCRr/a)은 비미토콘드리아 OCR에 대해 교정하기 위해 모든 비율에서 뺀다. 데이터는 XFe96 스페로이드 플레이트에 걸쳐 3-8개의 개별 웰 반복실험으로부터의 SEM에 ±한 평균이다. 약어: OCR = 산소 소비율; RA = 2 μM 로테논-2 μM 안티마이신 A. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
3일 동안 4 × 10개의3 개 세포/웰에서 성장한 MCF-7 구상체를 스페로이드 분석 마이크로플레이트 내에서 최적의 전달, 배치 및 분석을 결정하기 위한 모델로 사용하였다. 제조업체로부터 스페로이드 마이크로플레이트에 제공된 치수를 사용하여, 웰 표면을 최적의 스페로이드 배치를 위해 세 개의 구역-영역으로 분할하였고(그림 7A), 여기서 구역 1은 웰의 중심에서 최적 구역으로 강조되었다. 넓은 오리피스 피펫 팁을 사용하여 조심스럽게 피펫팅을 한 후, 구상체를 스페로이드 플레이트로 옮기고 중력 용출에 의해 우물 표면에 무작위로 분포시켰다(그림 7B). 중력 용출을 사용하여 구상체를 조심스럽게 옮긴 경우, 대부분의 구상체는 일반적으로 제조업체의 권장 전달 기술을 사용하여 마이크로 플레이트의 구역 1-2에서 발견 될 수 있습니다. 구상체가 열망에 의해 피펫 끝에서 쫓겨난 곳에서, 구상체는 종종이 영역 너머에 배치되었으며 현미경을 사용하여 볼 수 없었습니다.
스페로이드 배치 위치를 비교하기 위해, MCF-7 구상체를 지정된 구역 1-3 또는 구역 밖의 스페로이드 분석 마이크로플레이트로 옮겼다(도 7A). 이들 4개의 웰은 올리고마이신, BAM15, 또는 로테논-안티마이신 A를 첨가한 후 기준선에서 OCR의 동역학적 실험을 통해 추적되었다(도 7C). OCR은 각 주입 전의 세 사이클 판독값의 평균으로부터 계산되었다(도 7B). OCR은 선택된 4개의 웰(도 7C) 및 기준선 보정(도 7D)에서 200분에 걸쳐 운동적으로 측정되었다. 구상체가 구역 3 또는 구역 밖에 배치된 경우, 기준선 OCR은 구역 1 및 구역 2에 배치된 구상체보다 현저히 낮았다(그림 7C). 호흡기 화합물 올리고마이신, BAM15 및 로테논-안티마이신 A의 효과는 또한 구역 3 및 구역 밖 영역과 비교하여 구역 1과 구역 2에 배치된 구상체 사이에서 극적으로 달랐다. OCR의 증가는 구역 3 또는 구역 밖에 놓인 구상체에서 올리고마이신과 함께 관찰되었다 (도 7E). 더욱이, 구역 3 또는 구역 밖에 배치된 구상체는 로테논-안티마이신 A 주사 후 기준선보다 높은 OCR을 갖는 BAM15에 대해 과도하게 높은 반응을 경험하였다(도 7E). 구역 2 대 구역 1에 구상체가 배치된 기저 OCR(그림 7C)의 거의 2배 증가에도 불구하고, 모든 호흡기 화합물에 대한 반응의 폴드 변화는 매우 유사했으며(그림 7E), 구역 1 또는 2에 배치된 구상체 사이의 기저 OCR의 차이가 우물 내에 배치된 결과가 될 가능성은 거의 없음을 시사한다.

그림 7: 스페로이드 분석 마이크로플레이트 내에 구상체를 배치하면 XF 기술을 사용한 기저 OCR 및 미토콘드리아 조절제 효과가 결정됩니다. MCF-7 구상체를 4 × 10개의 3개 세포/웰에 시딩하고 XF RPMI를 함유하는 스페로이드 마이크로플레이트의 웰에 넣기 전에3일에 걸쳐 배양하고, 애질런트 시호스 XFe96 분석기를 사용하여 OCR ± 미토콘드리아 조절제에 대해 프로빙하였다. (a) 분석 지속 시간 후 스페로이드 분석 마이크로플레이트에서의 스페로이드 구역 위치의 현미경 사진; 스케일 바 = 500 μm 및 OCR은 pmol/min-1/well-1(B) 또는 % 기저(C)로 표현되는 시간에 따라 상응하는 웰로부터 포획된다. (d) 스페로이드 분석 마이크로플레이트 내의 상이한 구역에 배치된 MCF-7 구상체의 미토콘드리아 조절제 효과; 기저부에서 접이식 변화로 표현 된 데이터. (e) E에 제시된 데이터에 대한 각 미토콘드리아 조절제의 반응을 계산하기 위해 OCR 데이터 측정치(적색 원)가 사용되는 것을 강조하는 운동 추적 예. 표시된 데이터는 개별 우물 응답에서 나온 것입니다. 약어 : OCR = 산소 소비율. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
배경에 대한 선택 기준은 매우 중요합니다. 배경 보정을 위해 최외곽 웰을 사용하는 것은 모든 마이크로플레이트 웰을 대표하지 않으며, 이로 인해 스페로이드 마이크로플레이트 전체의 에지 효과로 인해 잘못된 데이터 가정이 도출되고 잘못된 데이터 결론이 발생할 수 있습니다. 이러한 관찰을 평가하기 위해, MCF-7 구상체를 사용하여 비히클 대조군, 올리고마이신, BAM15, 또는 로테논-안티마이신 A의 첨가에 반응하여 OCR 값을 도출하기 위한 검정 보정 절차를 비교하였다(도 8). 모든 호흡기 화합물은 선택된 화합물에 대해 예상되는 운동 OCR 프로파일을 산출하여 20-30 pmol/min/well의 평균 꾸준한 기저 호흡률을 나타냈다(도 8A). 그러나, 분석 데이터가 배경 온도 보정을 위해 최외곽 웰(A1, A12, H1 및 H12)을 사용하여 분석된 경우, 호흡기 화합물 첨가 후 OCR에 대해 밝혀진 값은 특히 낮았다; OCR은 로테논-안티마이신 A에 대해 음성 값을 산출하였다. 이러한 관찰에 응답하여, 대체 분석은 배경 온도 보정 웰로서 스페로이드 마이크로플레이트 전체에 랜덤하게 분포된 일련의 빈 웰을 사용하여 수행되었다(도 8B). 대안적인 배경 보정이 적용된 경우, OCR에 대한 모든 상대적 화합물 효과는 두 분석 세트 모두에서 동일하였다; 그러나 절대 OCR 값은 약 10pmol/min/well만큼 증가했습니다(그림 8). 이 데이터는 스페로이드 분석 마이크로플레이트에서 배경 온도 보정의 힘과 중요성을 강조하고 XF 분석을 위한 사용자 최적화의 중요성을 강조합니다.

그림 8: 스페로이드 분석 마이크로플레이트에 걸친 온도 구배에 대한 제어를 개선하기 위한 배경 보정을 위한 웰의 무작위 선택. OCR 데이터는 배경 보정(A)을 위한 권장 웰과 배경 보정을 위해 무작위로 할당된 웰(B)을 사용하여 그림 2A에서 외삽되었습니다. 약어 : OCR = 산소 소비율. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
세포 단일층과는 달리, 구상체는 3D 공간에서 세포의 이질적인 응집을 나타내므로 특히 이러한 데이터를 정규화 할 때 분석과 관련하여 철저한 고려가 필요합니다. 이 백서에서는 MCF-7 구상체에서 획득한 XF 데이터를 정규화하는 세 가지 방법을 제시합니다(그림 9). 정규화되지 않은 경우, OCR은 Pearson 상관 계수, P = 0.0057 (그림 9A)과 통계적으로 비교할 때 스페로이드 크기 (초기 세포 시딩 밀도에 의해 결정됨)와 유의하게 양의 상관 관계 (R2 = 0.98)를 나타냅니다. 이러한 선형 관계는 OCR이 초기 셀 시딩 밀도(R2=0.78)로 정규화되고 더 이상 스페로이드 크기와 유의한 상관관계가 없을 때 낮아진다(P=0.117, 도 9B). 이는 또한 스페로이드 부피로 정규화되는 경우이다(R2=0.77; 피어슨 상관 계수 P = 0.120, 도 9C) 및 핵 dsDNA 함량 (R2 = 0.58; 피어슨 상관 계수 P = 0.233, 그림 9D). 이 데이터는 구상체의 미토콘드리아 대사를 조사 할 때, 특히 크기가 다른 경우 XF 데이터를 정상화하는 것이 중요하다는 것을 강조합니다.

도 9: 세포 구상체로부터 획득한 세포외 플럭스 데이터의 정규화. (A) 원시 OCR 데이터는 3일에 걸쳐 배양된 MCF-7로부터 수득되었고, 스페로이드 시딩 밀도와 OCR 사이의 상관 계수를 얻기 위해 피어슨의 모델을 사용하여 플롯팅되었다; P 값은 0.05로 설정됩니다. (b) 원시 OCR 데이터는 초기 스페로이드 시딩 밀도에 대해 정규화되었고; (c) 현미경 평탄도로부터 수득된 MCF-7 스페로이드 부피; 및 (D) 피어슨의 상관계수를 사용하여 비교한 핵 ds DNA 함량. 약어: OCR = 산소 소비율; ds DNA = 이중 가닥 DNA. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
| 세포주 | 시드 밀도 (우물) | 스페로이드 성장 (일) | 최종 스페로이드 부피(μM3) | 기저 OCR (pmolO2/분/웰) | 기저 OCR 충족에 대한 민감도 (예 / 아니오) |
| 증권 시세 표시기 | 1000 | 5 | 9.52E+06 | 28 ± 3.5 | 예 |
| 증권 시세 표시기 | 2000 | 5 | 2.38E+07 | 26 ± 1.4 | 예 |
| 증권 시세 표시기 | 4000 | 5 | 4.92E+07 | 36 ± 3.1 | 예 |
| 증권 시세 표시기 | 8000 | 5 | 1.11E+08 | 50 ± 7.9 | 예 |
| HepG2 | 1000 | 5 | 1.11E+07 | 15 ± 0.7 | 아니요 |
| HepG2 | 2000 | 5 | 2.88E+07 | 23 ± 1.8 | 예 |
| HepG2 | 4000 | 5 | 5.46E+07 | 31 ± 1.7 | 예 |
| HepG2 | 8000 | 5 | 1.21E+08 | 52 ± 2.8 | 예 |
| A549 | 1000 | 5 | 2.11E + 07 | 30 ± 2.5 | 예 |
| A549 | 2000 | 5 | 3.57E+07 | 41 ± 1.6 | 예 |
| A549 | 4000 | 5 | 6.93E+07 | 53 ± 7.2 | 예 |
| A549 | 8000 | 5 | 1.44E+08 | 65 ± 8.4 | 예 |
| MCF-7 | 1000 | 3 | 1.60E+07 | 29 ± 0.8 | 예 |
| MCF-7 | 2000 | 3 | 2.52E+07 | 37 ± 1.7 | 예 |
| MCF-7 | 4000 | 3 | 6.00E + 07 | 46 ± 1.7 | 예 |
| MCF-7 | 8000 | 3 | 1.06E+08 | 66 ± 2.9 | 예 |
표 4: 단일 3D 구상체에서 기초 OCR 측정을 결정하기 위한 최적화된 파라미터. 약어 : OCR = 산소 소비율.
보충 파일 1 : 스페로이드 크기 및 부피 분석. 이 파일을 다운로드하려면 여기를 클릭하십시오.
보충 파일 2 : 스페로이드 마이크로 플레이트의 구상체로부터 이중 가닥 DNA의 정량화. 이 파일을 다운로드하려면 여기를 클릭하십시오.
보충 파일 3: 신뢰할 수 있는 XF 분석 데이터 세트를 얻는 데 필요한 반복실험 횟수에 대한 권장 사항입니다. 이 파일을 다운로드하려면 여기를 클릭하십시오.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
주요 결과 및 결과
이 논문은 XFe96 XF 분석기와 함께 일련의 암 유래 세포주를 사용하여 단일 3D 구상체의 미토콘드리아 에너지 대사를 조사하는 상세한 프로토콜을 제공한다. 강제 응집을 위한 세포 기피제 기술을 사용하여 A549, HepG2/C3A, MCF7 및 SK-OV-3 세포 구상체의 신속한 배양을 위한 방법이 개발되고 기술된다. 이 프로토콜은 (1) 스페로이드 배양 프로토콜의 최적화 및 기술 제조업체의 특정 스페로이드 분석 마이크로 플레이트로의 스페로이드 처리 및 전달을 포함하여 XF 기술로 스페로이드 대사를 조사하는 많은 고려 사항을 해결합니다. (2) 사용되는 호흡기 화합물의 농도 및 시간 의존성 화합물 침투; (3) 사용되는 주사 전략; (4) 실험 그룹 간의 데이터를 정규화하는 방법. 이러한 모든 고려 사항은 현재 논문에서 검토되었으며 아래에서 더 자세히 논의됩니다. 이러한 방법은 XFe96 Flux 분석기를 사용하여 단일 3D 구상체를 사용하여 일관된 대사 산소 플럭스 데이터를 생성하는 단순화된 접근법으로 제시됩니다. 이 실험적 접근법은 기본 실험실 설정 내에서 쉽게 구현 될 수있는 다른 스페로이드 모델에 사용하기위한 출발점 및 루브릭으로 사용할 수 있습니다.
고려 사항
XF 기술의 스페로이드 성장, 크기 및 감도
XF 기술로 재현 가능한 데이터를 설정하려면 특정 모델에 대한 분석을 특성화하고 최적화하는 것이 필수적입니다. 이 접근법은 세포의 기본 단층에서 비교적 간단합니다. 그러나 이것은 세포를 3D 구상체로 재배 할 때 추가적인 도전을 제시합니다. 여기에 제시된 실험 동안, 제조자로부터의 RPMI 배지는 사용 시에 보충되었다. 일부 세포주, 즉 HepG2/C3A가 DMEM 성장 배지에서 배양되었다는 것이 주목되는 반면, 이러한 비교적 짧은 분석(~3-5시간) 동안, RPMI-DMEM 제형으로의 치환은 XF 분석에 제한적인 영향을 미쳤다. 두 배지의 제형은 매우 유사하며, 사용자는 보충, 예를 들어, 글루코스 증가, 탄수화물 공급원의 추가 첨가를 통해 그들의 세포 배양 배지의 매트릭스와 일치하도록 Seahorse RPMI 배지를 '조정'할 수 있다. 모든 XF 완충액 및 배지의 최종 제형화에 중요한 것은 XF 프로브 카트리지 플레이트 내의 형광 프로브를 방해할 가능성이 있는 페놀-레드 및 중탄산나트륨의 부재이며, 이는 세포 배양 인큐베이터에 존재하는CO2 완충의 부족으로 인해 알칼리도를 야기할 것이다. 다른 매체 및 버퍼는 사내에서 구매 및/또는 제작될 수 있다. 예를 들어, 크렙스 링거 HEPES 버퍼는 스페로이드 모델을 포함한 많은 다른 세포에서 호흡을 평가하는 데 사용할 수있는 간단한 버퍼입니다. 그러나 XF 분석 사용자는 배지 / 버퍼 및 보충의 변화가 전체 완충 능력을 변경할 수 있음을 유의해야합니다. 이것은 사용자가 ECAR 측정에 관심이있을 때 특히 우려되며, ECAR 변환을 양성자 유출률 (PER)으로 변환 할 수 있도록 매체의 버퍼 계수를 평가해야합니다.
XF 기술로 측정된 세포 OCR은 웰 내의 세포 수가 시스템의 민감도 내에 있을 때 세포 밀도에 비례하므로, 단일 3D 구상체를 사용하여 이러한 관계를 조사하는 것이 중요하였다. 웰당 1,000개, 2,000개, 4,000개 또는 8,000개 세포의 밀도로 시딩된 4개의 서로 다른 암 세포주에서 배양된 단일 3D 구상체의 OCR을 조사함으로써, XFe96 분석기가 서로 다른 세포 시딩 밀도에서 성장한 3D 구상체 사이의 미토콘드리아 호흡 속도의 변화를 포착할 수 있을 만큼 민감하다는 것을 보여줍니다(그림 3). 우리는 OCR을 프로빙하기 위한 3D 구상체 형성을 위한 세포 시딩 밀도, 따라서 스페로이드 부피의 최적 범위가 세포 유형에 따라 다르다는 것을 보여준다. 이는 OCR과 시딩 밀도 또는 스페로이드 부피 사이의 선형 관계로 표시됩니다(그림 3). A549 및 HepG2/C3A 셀의 경우, OCR 감도를 위한 최적의 시딩 밀도는 1,000 ~ 8,000 셀/웰 사이였습니다. MCF-7의 경우 2,000-8,000 세포/웰, SK-OV-3 세포의 경우 4,000-8,000 세포/웰이었습니다. 이러한 데이터는 XF 기술을 사용하여 OCR을 평가할 때 스페로이드 크기의 최적화가 특히 중요하다는 것을 보여줍니다.
최소 및 최대 스페로이드 부피 및 기저 OCR에 대한 고려 사항
일반적으로, 이러한 실험을 위해 제조에 의해 추천되는 측정가능한 OCR 파라미터에 대한 최소 및 최대 임계값이 항상 존재할 것이다. XFe96 분석기의 경우, 20pmol O 2/min/웰과 200pmol O2/min/well 사이의 기저 OCR이 각각 하한과 상한입니다. 이것은 단층 세포 및 구상체를 사용하는 경우이며, 실험 모델이 이러한 동적 OCR 범위 내에 위치하는 경우는 이용 가능한 생물학적 물질의 양, 예를 들어, 단층으로서의 세포의 수 또는 구상체의 크기에 의존할 것이다. OCR 임계값이 여기에 사용된 스페로이드 모델에 의해 어떻게 달성되었는지에 대한 예는 표 4를 참조한다. 이러한 데이터가 또한 레벨 데이터로서 이들 측정으로부터 이용가능한 우물 내의 산소 수준을 확인하는 것이 현명할 수 있다. 이것은 품질 관리 목적으로 각 실험에서 일상적으로 볼 수 있어야합니다. 우물에 산소 고갈이 있으면 데이터 내에서 분명히 알 수 있습니다. 이 경우 실험 내에서 측정 주기를 조정해야 할 수도 있습니다. 예를 들어, 웰 내의 산소 레벨이 측정 사이클 내의 다음 측정 기간 전에 회복되도록 혼합 단계를 증가시킨다. 가능하지만, 우리는 이것이 설명 된 세포주를 사용하는 단일 스페로이드 실험에 대해 매우 드물다는 것을 발견했습니다.
세포외 플럭스 분석을 위한 미토콘드리아 언커플러 선택
카르보닐 시아나이드 4-(트리플루오로메톡시)페닐히드라존(FCCP)21, 카르보닐 시아나이드 m-클로로페닐히드라존(CCCP)22 또는 BAM15 23과 같은 양성자 이오노포어는 미토콘드리아 막에 걸친 전기화학적 양성자 구배를 방해하고, ATP의 생성을 억제하고, 궁극적으로 미토콘드리아 호흡을 분리할 수 있는 강력한 소분자 화학물질이다(24) . 새로운 소분자는 이러한 목적을 위해, 특히 대사성 질환의 치료에서 계속 개발되고 있다25,26,27; 두 가지 우수한 리뷰28,29를 참조하십시오. 반대로, 산화적 호흡의 비결합은 바람직하지 않은 오프타겟 독성(30)과 연결되었다. 그러나, 시험관내 세포 분석에서, FCCP 분자는 미토콘드리아 막 전위를 탈분극시키고, 원형질막 탈분극, NA+ 이온 플럭스(31)를 교란시키는 것과 같은 오프 타겟 효과를 발휘한다; 세포 단백질 처리(32)를 방해하고, 심지어 세포 노화(33)를 유도한다. BAM15는 원래 2013년에 원형질막(23)에 대한 영향을 최소화한 미토콘드리아 언커플러로 도입되었으며, 전체 세포에서 마이크로몰 범위의 프로토노포릭 활성과 분리된 미토콘드리아에서 나노몰 범위23,34의 프로토노포릭 활성이 있다.
원형질막 탈분극에 대한 FCCP의 효능을 감안할 때, BAM15는 세포외 플럭스 분석에서 무손상 전체 세포에서 호흡을 풀기 위한 보다 신뢰할 수 있는 프로토노포어이다. FCCP와 그 대응물인 CCCP는 최대 호흡 능력을 분석하기 위해 50년 이상 사용되어 왔으며 XF 연구에서 계속 널리 사용되고 있지만, 이러한 소분자의 사용은 종종 미토콘드리아 및 세포 대사 능력을 과소평가합니다. 이것은 부분적으로 XF 기술을 사용하는 많은 출판물이 FCCP를 사용할 때 부정적인 예비 호흡 용량을보고하거나 진정한 미토콘드리아 호흡 용량을 과소 평가하는 함정에 빠지는 이유와 관련이 있습니다. 무손상 세포 및 조직에서 FCCP의 추가 효능은 종종 미토콘드리아 기능 저하를 초래하며, 세포는 매우 낮은 농도에서도 추가 후 여러 측정 사이클에 걸쳐 최대 호흡 용량을 유지하기 위해 적절하게 작동하기 위해 고군분투 할 수 있습니다35. 따라서, FCCP에 대한 세포의 반응은 초기 측정 주기 주기 이후에 감소하는 많은 연구에서 발견될 수 있다. FCCP가 XF 분석에 일상적으로 사용되었지만, BAM15는 10 μM3만큼 높은 농도에서 완전히 탈분극된 미토콘드리아에서 최대 호흡 능력을 유지할 수 있다는 점을 감안할 때 전체 세포 또는 스페로이드 모델과 관련된 경우에 우선적으로 사용됩니다. 더욱이, BAM15는FCCP3보다 더 큰 정도로HCO3- 및 H+를 형성하기 위해CO2의 수화를 통한 영양 산화의 그것과 일치하는 세포외 산성화에 대한 효과를 유도한다. 그럼에도 불구하고, 단리된 미토콘드리아 및 투과화된 세포의 경우, 이들 비커플러 중 임의의 것이 정확한 농도로 적정된다면 미토콘드리아 비결합에 대한 BAM15 뿐만 아니라 수행되어야 한다.
화합물 침투 및 분석 사이클링의 동역학
XF 분석기를 사용하여 3D 구상체로 일반적인 MitoStress 테스트를 수행하는 데 사용되는 화학 화합물의 농도, 침투 및 운동 프로파일은 해결하기가 더 복잡합니다. 구상체가 3D 구조를 가지고 있다는 것을 감안할 때, 스페로이드의 직경을 가로 지르는 분자의 침투는 세포 단일층을 가로 지르는 것보다 무한히 복잡한 과정입니다. 예를 들어, 화학요법적 소라페닙에 대한 동역학적 침투 및 따라서, 민감성은 스페로이드 연령 및 따라서, HepG2 스페로이드 모델(36)에서의 크기에 의해 결정되었다. 생물학적 표적에 도달하는 소분자 화학물질(예를 들어, 약물, 나노입자)의 능력은37,38을 통해 동적으로 침투 및 확산되는 시스템의 복잡성을 포함하는 몇 가지 근본적인 인자에 의존한다. 이것은 종양 조직(39)을 표적화하는 약물에 대해 특히 그러하다. 3D 구상체의 맥락에서 종양 표적화와 유사하게, 크기, 치밀성, 및 약물 수송체 단백질의 발현과 같은 다른 표현형 반응은 생물학적 반응을 이끌어내는 데 필요한 화합물의 침투 시간 및 농도를 지배할 수 있다.
이 프로토콜에서, 우리는 ATP 신타제 억제제 올리고마이신, 프로토노포어 및 미토콘드리아 언커플러 BAM15, 및 콤플렉스 I 및 콤플렉스 III 억제제인 로테논과 안티마이신 A의 조합에 반응하여 침투 시간 및 소분자 농도에 관한 문제를 해결했다. 이러한 일반적인 호흡기 화합물의 다중 적정에 노출된 단일 MCF-7 구상체의 OCR을 조사함으로써, 우리는 정상 상태 호흡률을 유도하는 데 필요한 각 화합물의 최적 농도가 단층 세포의 최적 범위 내에 있음을 입증한다(그림 4). 중요하게도, 그들의 단층 대응물과 다르게, 주사들 사이의 측정 사이클 수를 증가시키는 것이 단일 3D 구상체에서 정상 상태 OCR을 달성하는 열쇠임을 보여준다. 이 데이터는 이러한 접근법을 사용하여 3D 구상체의 미토콘드리아 호흡 매개 변수를 탐색 할 때 화합물 침투와 각각의 운동 프로파일의 중요성을 강조합니다. 도 3, 도 4 및 도 5에 제시된 데이터에 의해 정보된 스페로이드 최적화 특성, 화합물의 농도 및 측정 주기 시간을 사용하여, 암 유래 3D 구상체의 범위에서 미토콘드리아 산화 대사의 특정 파라미터를 조사하기 위한 검증된 MitoStress 테스트가 확립되었다(도 6). 중요도, 일부 단층 암 세포주(40)와 마찬가지로, 특정 암 유래 3D 구상체의 최대 호흡 용량(결합되지 않은 자극된 호흡의 속도)은 올리고마이신에 의해 억제되었다(도 5). 구체적으로, A549 또는 MCF-7 세포로부터 성장한 3D 구상체는 올리고마이신 주입 후 BAM15와 결합하지 않을 때 올리고마이신이 없는 BAM15에 의해 결합되지 않은 것에 비해 현저히 낮은 최대 호흡률을 보였다(도 5F 및 도 5H). 이 효과가 다른 3D 스페로이드 배양 내에 존재할 수 있다는 것을 감안할 때, 이전에 검증 된 프로토콜이 사용되지 않는 한, 올리고 마이신 없이 3D 구상체의 최대 호흡 용량을 추정해야한다고 제안합니다.
세포 구상체에서 글리콜 플럭스의 척도로서 ECAR 데이터의 동시 수집
기술 제조업체의 문헌 또는 정보에서 일반적으로 볼 수 있듯이 ECAR로 측정 된 구상체의 글리콜 레이트는 OCR과 함께 캡처 할 수있는 보조 매개 변수입니다. ECAR 단독으로는CO2 에서HCO3- 및 H+로의 수화로부터 발생하는 XF 분석 완충액 또는 미토콘드리아 산성화의 첨가의 완충 능력에 대해 보정되지 않기 때문에 어떠한 XF 실험에서도 유용하거나 의미있는 파라미터가 아니다. ECAR은 이러한 데이터 보정이 적용된 후에만 통찰력이 있으며, 그 후에 글리콜 플럭스에 대한보다 정확한 결론을 제공 할 수있게됩니다. 버퍼링 용량을 수정하여 보다 의미있는 PER 데이터를 생성하려면 스페로이드 마이크로플레이트에 대한 마이크로챔버의 부피를 알아야 한다. 제조업체는 스페로이드 마이크로 플레이트로 이에 대한 실제 부피를 제공 할 수 없었기 때문에 PER 데이터를 쉽게 결정할 수 없습니다. 사실, 이러한 측정이 경험적으로 달성 될 수 있지만, 이것은이 원고의 범위를 벗어났습니다. 그러나, 적절한 보정 및 주어진 스페로이드 크기에 대한 마이크로챔버의 부피를 알면(예를 들어, 스페로이드 밀도의 척도를 획득하는) 웰 내에 존재함으로써, ECAR 데이터는 의미가 있을 것이고, 글리콜성 PER의 계산이 이루어질 수 있을 것이다. 따라서 XF 데이터는 구상체에서 글리콜 및 산화 대사를 조사하는 데 더 많은 정보를 제공 할 수 있지만 이러한 매개 변수가 깊이있게 고려 된 경우에만 가능합니다.
스페로이드 형성, 취급, 전송 및 이동
일부 세포주는 다른 세포주보다 구상체 형성에 더 적합하고, 구상체를 전혀 형성하지 않을 수 있으며, 예를 들어, MCF-7 난소암 세포41,42는 다른 세포주에 비해 고도로 원형 구상체를 형성한다(도 3). 또 다른 예로서, Capan-1 췌장암 세포는 Panc-1 또는 BxPC343보다 더 나은 구상체를 형성하는 것으로 나타났다. 유사하게, 간암종 세포주는 HepG2 대 HepG2/C3A 9,45,46 또는 HepaRG 구상체17,47,48의 경우와 같이 향상된 약물 대사 또는 알부민 생산과 같은 표현형의 변화가 관찰된 바와 함께 콤팩트한 구상 체 5,44를 형성하는 가변적인 능력을 갖는 것으로 알려져 있다. . 따라서 사용자는 그에 따라 스페로이드 배양 기술을 최적화하고 적정 실험을 수행하여 최적의 시딩 밀도 및 재배 시간 과정을 결정해야합니다. 또한, 분석 매질의 제형 및 조성은 메틸셀룰로스의 첨가를 포함하는 스페로이드 제형에 영향을 미치는 것으로 나타났으며, 종종 매트릭스 점도 43,49,50을 증가시키기 위해 매질에 첨가된다. 따라서, 최적의 세포 배지 조성은 사용되는 모든 세포주에 대해 경험적으로 결정되어야 한다.
스페로이드 배양 전반에 걸친 배지 교환의 수는 사용된 세포주에 의해 결정된다. 그러나 일반적으로 2-3 일마다 절반 부피의 중간 교환이 대부분의 경우 영양소를 보충하기 위해 적용됩니다. 우리는 XF 분석 연구에서 스페로이드 모델의 신속한 개발 및 배치를 위해 상업적으로 이용 가능한 소스의 세포 기피성 마이크로 플레이트를 사용하여 3D 구상체를 생성하기 위해 강제 응집 접근법을 사용했습니다. 그러나, 대안적인 플랫폼들은 다른 세포 유형들, 예를 들어, 교수형-드롭 또는 매트릭스-임베디드 접근법들로부터 구상체들을 생성하는데 더 적합할 수 있다. 자원-제한된 실험실에서, 사용자는 초기 스페로이드 방법 개발 단계의 경제적 비용을 상당히 감소시키기 위해 세포-구충성 마이크로플레이트 표면(51,52)의 형성을 위한 아가로스-액체 오버레이 기술을 바라볼 수 있다. 배양 용기 사이의 구상체의 이동은 XF 분석 및 기타 하류 분석을 수행하는 데 필요합니다. 전달의 용이성은 일반적으로 스페로이드 크기와 전체 밀도에 의해 결정됩니다. 스페로이드 무결성을 유지하기 위해 P200 또는 P1000 와이드 오리피스 피펫 팁을 사용하는 것이 좋습니다. 더 작은 보어 피펫 팁은 구상체의 기계적 파괴를 위험에 빠뜨릴 수 있으며, 이는 상업적으로 구입하거나 오리피스를 늘리기 위해 피펫 팁의 끝을 간단히 트리밍하여 조심스럽게 만들 수 있습니다. 그러나 이러한 접근 방식은 팁 끝 주위의 플라스틱에 퍼링을 도입하기 쉬울 수 있으며, 이는 취급 중에 기계적 장애를 일으킬 수 있습니다. 백라이트 또는 라이트박스의 사용은 스페로이드 분석 마이크로플레이트 내로의 스페로이드의 성공적인 전달을 보장하기 위한 필수 단계로서 해부 현미경 하에서의 스페로이드 취급 및 관찰에도 유용하다. 더욱이, 스페로이드 분석 마이크로플레이트의 웰 내의 스페로이드 위치는 특히 중요하며, 전형적인 MitoStress 테스트 동안 OCR 및 화합물 효과에 직접적으로 영향을 미치며(도 7), 스페로이드 위치와 센서 프로브 형광단의 관계로 인한 것일 가능성이 가장 높다.
배경 보정 및 온도 제어 웰
마이크로플레이트 기반 분석의 사용은 여러 연구 분야에서 널리 사용되는 접근법입니다. 그러나 그들의 사용은 몇 가지 실질적인 도전을 제시합니다. 다른 실험 접근법, 특히 96 (또는 그 이상) 배열 형식을 사용하는 접근법에서 사실과 마찬가지로, 마이크로 플레이트 지오메트리 및 포지셔닝은 시간이 지남에 따라 플레이트 전체의 온도 및 가스 교환 구배에 영향을 줄 수 있으며 종종 '에지 효과'53,54라고합니다. 우리는 스페로이드 분석 마이크로플레이트의 경우에도 마찬가지라는 것을 발견했다. 제조업체의 지침과 프로토콜에 따라 최외곽 모서리 우물(A1, A12, H1 및 H12)은 항상 XFe96 분석기의 배경 보정 및 온도 제어 웰로 지정됩니다. 반대로, 24웰 어레이 형식에서는 A1 및 D6이 제어 웰로 지정되며, B4 및 C3에서 플레이트 중앙에 고르게 분산된 두 개의 다른 웰이 함께 지정됩니다. XF 스페로이드 분석을 수행 할 때 제조업체의 지침을 사용하여 처음 수집 된 데이터에서 상당한 편차가 있음을 발견했습니다. 이것은 데이터의 획득을 시작하기 전에 온도 및CO2 함량에 대한 분석이 사전 평형을 보장하기 위해 필요한 단계의 포함에도 불구하고, 종종 특정 호흡기 억제제의 주입 후 OCR에 대한 음의 값을 산출하였다 (도 8).
우리는 이러한 관찰이 스페로이드 분석 마이크로플레이트 전반의 에지 효과 때문일 가능성이 있음을 발견했습니다. 도 8에서, 배경 제어 웰을 마이크로플레이트 전체에 걸쳐 재분배하는 것을 발견하였고, XF 데이터는 대략 2배 조정되었다. 가장 가능성이 높은 두 가지 이유는 (1) 엣지 웰에서의 증발 효과로 인해 XFe96 프로브가 샘플링할 수 있는 총 부피 챔버가 작아지고 (2) 배경 보정을 위해 지정된 웰과 샘플 웰 사이의 부적절한 온도 평형으로 인해 OCR을 마스킹하거나 과도하게 팽창시키는 데이터 세트가 발생하기 때문입니다. 따라서 이러한 결과를 피하기 위해, 특히 스페로이드 분석의 맥락 내에서, 사용자는 스페로이드 분석 마이크로플레이트 전체에 걸쳐 배경 교정을 위해 지정된 웰을 재분배하고 XF 데이터를 획득하기 전에 분석을 사전 평형화하는 데 필요한 조치를 취하는 것이 좋습니다.
데이터 정규화
XF 기술로 단일 3D 구상체의 미토콘드리아 에너지 대사를 조사하기 위한 상세한 프로토콜을 제공하는 것 외에도, 이 논문은 3D 구상체로 얻은 미토콘드리아 호흡률 데이터를 정상화하는 가능한 방법도 제시한다. 상이한 세포 시딩 밀도에서 배양된 MCF-7 구상체로 얻은 호흡률 데이터를 사용하여(도 3), 초기 세포 시딩 밀도, 스페로이드 부피 및 dsDNA 함량으로 정규화될 때 증가된 크기 및 직경의 MCF-7 구상체로부터의 기저 미토콘드리아 호흡률을 제시한다(도 9). XF 데이터 세트의 정확한 해석을 위해서는 적절한 정규화 방법이 가장 중요하며, 특히 시험관내 3D 스페로이드 모델과 다른 세포 유형을 비교할 때 더욱 그렇습니다. 정규화가 잘못되면 데이터 세트 간에 비교할 수 없는 잘못된 결과가 발생할 수 있습니다. 단백질 함량은 스페로이드 XF 데이터의 정상화에 바람직하지 않은데, 전처리는 호흡률에 유의한 영향 없이 단백질 합성 속도에 영향을 미칠 수 있기 때문이다. 더욱이, 중요하고 일관성 없는 양의 단백질은 세포 용해시 스페로이드 마이크로플레이트에 결합할 수 있으며, 웰 사이에 단백질 함량의 변화를 가져올 수 있다. 이것은 단백질을 포함 할 수있는 생체 분자 접착제가 필요한 구상체 또는 비 부착 세포를 사용하는 XF 분석에서 더욱 복잡 할 수 있습니다.
세포내 단백질 함량과는 달리, 핵 DNA 함량은 세포 유형에 무관하며 세포 수에 비례한다(도 9D)-세포 수 정량화를 위한 구상체의 분해보다 더 정확하고 시간이 덜 소요되는 접근법이다. 반대로, Yepéz et al.55는 섬유아세포 세포의 단층에서 XF 분석을 수행하여, XF 데이터를 세포 수로 정규화하면 정규화 이전보다 데이터의 분산이 더 크다는 것을 발견했다. 핵 DNA 함량은 분화된 상태 또는 표현형과 무관하며, 따라서 단백질 함량보다 XF 분석에서 스페로이드 데이터의 정규화에 대해 더 정확하다. DNA 함량은 또한 다른 대사-연결된 데이터세트(56)의 분석을 위한 입증된 전략이었다. 그러나, 핵 DNA 함량은 구상체 내에 존재하는 모든 세포로부터 정량화된다는 점에 유의하는 것이 중요하다; 따라서 DNA 함량에 대한 정상화는 구상체가 세포 생존력의 상당한 손실을 초래할 수있는 치료를받는 XF 데이터 세트에는 권장되지 않습니다. 이러한 데이터 세트의 경우, 실현 가능한 경우, 세포 생존력에 대한 정규화가 선호되거나, 데이터는 기초 호흡으로 기준선으로 보정될 수 있다.
예비 호흡 용량을 데이터 정규화의 중요성에 대한 모범으로 사용
예비 호흡 용량은 최대 미토콘드리아 호흡 능력에서 기저 미토콘드리아 호흡률을 뺀 비율의 척도입니다(그림 6). 그러나 이러한 유형의 데이터를 속도, 즉 특정 실험 내에서 pmolO2 / min / well으로보고하는 문제는 데이터가 정규화가 불가능하다는 것입니다. 스페로이드 데이터를 세포 밀도 / DNA 함량으로 정규화하더라도, 이것은 종종 세포 내의 미토콘드리아 밀도에 대해 정규화되어야하는 핵심 매개 변수를 배제합니다. 미토콘드리아 밀도의 변화가 기저 및 최대 호흡의 비례적 변화로 이어질 것이라는 점을 감안할 때, 예비 용량 또한 증가 할 것입니다. 예를 들어, 스페로이드 OCR기저 부가 200이고 OCR최대 값이 400인 경우 예비 용량은 200으로 보고됩니다. OCR기초 가 100이고 OCR이최대 인 경우 예비 용량도 100입니다. 그러나 백분율로, 그들은 모두 최대치의 50 % (또는 기초의 100 %)입니다. 따라서, 예비 용량은 pmols O2/min/well로 계산될 때 200과 100의 속도의 차이에도 불구하고, 이들두 예들 사이에서 변경되지 않는다. 내부적으로 정규화 된 값은보다 안정적이고 통찰력이있어 XF 데이터를 연구 및 프로젝트 전반에 걸쳐 비교할 수 있습니다. 여분의 호흡 능력을 위해이 작업을 수행하기 위해, 우리는 이것을 절대 비율 대신 최대 호흡의 백분율로 제시하기로 결정했습니다. 이것은 또한 기저 호흡의 백분율로 제시 될 수 있습니다. 이것은 세포 또는 구상체로 작업하는 경우에 해당됩니다. 그러나, 마이크로웰 플레이트 내의 스페로이드의 위치가 절대 OCR을 변화시킬 수 있지만 억제제 또는 언커플러와의 상대적 변화는 변화시키지 않는다는 점을 감안할 때, 구상체에서 내부적으로 정규화된 반응을 폴드 변화 또는 백분율로 보는 것이 더 중요하다.
여기서 생성된 스페로이드 모델은 고전적인 2D 모델에서는 캡처할 수 없는 다양한 세포 유형과 아키텍처를 제시합니다. 이들은 세 차원에서의 세포의 이질적, 공간적 배열, 강화된 세포-세포 접촉(예를 들어, 갭 접합 및 세포외 매트릭스의 형성), 및 스페로이드 직경을 가로지르는 생화학적 구배(예를 들어, pH 구배, 영양소에 대한 산소 확산 접근)를 포함한다. 시험관내 스페로이드 생물학을 연구하기 위해 세포외 플럭스를 사용하면 대사 섭동 관찰을 통해 약물 요법에 대한 최적의 표적을 확인할 수 있습니다. 이들은 시험관 내 구상체로부터 생체내 종양으로 외삽될 수 있고, 스페로이드-종양 대사, 예를 들어, 스페로이드 성장 동안 탄수화물 이용을 표적으로 할 수 있는 경로를 확인할 수 있다. 치료 양식은 초기 성장 단계에서 구상체를 표적화하는 데 효과적 일 수 있지만 대사 네트워크 복잡성이 성숙함에 따라 스페로이드 성장의 후기 단계에서는 덜 효과적이라는 것이 입증되었습니다. 결론적으로, 생물학적 연구에서 3D 세포 배양 모델 및 정교한 분석 기술의 개발은 타의 추종을 불허하는 잠재력을 지닌 역동적이고 빠르게 변화하는 분야가 될 것입니다. 시험관 내 세포 배양 구상체의 세포 외 플럭스 분석은 인간 관련 생물학을 더 잘 이해하고, 연구에서 동물 모델의 사용을 줄이며, 환자 중심 연구를 향상시키기 위해 외삽 될 수있는 연구 결과를 발전시키기위한 최첨단 연구 방법으로 사용될 수 있습니다.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
저자는 선언 할 이해 상충이 없습니다.
Acknowledgments
N.J.C는 Sygnature Discovery Ltd (BB/M01116X/1, 1940003)와 함께 BBSRC MIBTP CASE Award의 지원을 받았습니다.
Materials
| Name | Company | Catalog Number | Comments |
| A549 | ECACC | #86012804 | Lung carcinoma cell line |
| Agilent Seahorse XF RPMI Medium, pH 7.4 | Agilent Technologies Inc. | 103576-100 | XF assay medium with 1 mM HEPES, without phenol red, sodium bicarbonate, glucose, L-glutamine, and sodium pyruvate |
| Agilent Seahorse XFe96 Extracellular Flux Analyzer | Agilent Technologies Inc. | - | Instrument for measuring rates of spheroid oxygen uptake in single spheroids |
| Antimycin A | Merck Life Science | A8674 | Mitochondrial respiratory complex III inhibitor |
| BAM15 | TOCRIS bio-techne | 5737 | Mitochondrial protnophore uncoupler |
| Black-walled microplate | Greiner Bio-One | 655076 | For fluorescence-based assays |
| CELLSTAR cell-repellent surface 96 U well microplates | Greiner Bio-One | 650970 | Microplates for generating spheroids |
| CellTiter-Glo 3D Cell Viability Assay | Promega | G9681 | Assay for the determination of cell viability in 3D microtissue spheroids |
| Cultrex Poly-D-Lysine | R&D Systems a biotechne brand | 3439-100-01 | Molecular cell adhesive for coating XFe96 spheroid microplates to facillitate attachment of spheroids |
| D-(+)-Glucose | Merck Life Sciences | G8270 | Supplement for cell culture growth and XF assay medium |
| Dulbecco’s Modified Eagle Medium (DMEM) | Gibco | 11885084 | Culture medium for HepG2/C3A spheroids |
| EVOS XL Core Imaging System | Thermo Fisher Scientific | AMEX1000 | Phase-contrast imaging microscope |
| EZ-PCR Mycoplasma test kit | Biological Industries | 20-700-20 | Mycoplasma screening in cell cultures |
| FIJI Is Just Image J | Analysis of collated images | ||
| Foetal bovine serum | Merck Life Science | F7524 | Supplement for cell culture medium |
| HepG2/C3A | ATCC | #CRL-10741 | Hepatic carcinoma cell line, a clonal derivative of the parent HepG2 cell line |
| Lactate-Glo | Promega | J5021 | Assay for measurement of lactate within spheorid culture medium |
| L-glutamine (200 mM solution) | Merk Life Sciences | G7513 | Supplement for cell culture growth and XF assay medium |
| M50 Stereo microscope | Leica Microsytems | LEICAM50 | Stereo dissection micrscope; used for spheorid handling |
| MCF-7 | ECACC | #86012803 | Breast adenocarcinoma cell line |
| Oligomycin from Streptomyces diastatochromogenes | Merck Life Science | O4876 | ATP Synthase Inhibitor |
| Penicilin-Streptomycin | Gibco | 15140122 | Antibiotics added to cell culture medium |
| Quant-iT PicoGreen dsDNA Assay Kit | Initrogen | P7589 | Analysis of dsDNA in spehroids |
| Rotenone | Merck Life Science | R8875 | Mitochondrial Respiratory Complex I Inhibitor |
| RPMI 1640 | Gibco | 21875091 | Culture medium for A549, MCF7, and SK-OV-3 spheroids |
| Seahorse Analytics | Agilent Technologies Inc. | Build 421 | https://seahorseanalytics.agilent.com |
| Seahorse XFe96 Spheroid FluxPak | Agilent Technologies Inc. | 102905-100 | Each Seahorse XFe96 Spheroid FluxPak contains: 6 Seahorse XFe96 Spheroid Microplates (102978-100), 6 XFe96 sensor cartridges, and 1 bottle of Seahorse XF Calibrant Solution 500 mL (100840-000) |
| Serological pipette: 5, 10, and 25 mL | Greiner Bio-One | 606107; 607107; 760107 | Consumables for cell culture |
| SK-OV-3 | ECACC | #HTB-77 | Ovarian adenocarcinoma cell line |
| Sodium pyruvate (100 mM solution) | Merck Life Science | S8636 | Supplement for cell culture growth and XF assay medium |
| T75 cm2 cell culture flask | Greiner Bio-One | 658175 | Tissue culture treated flasks for maintaining cell cultures |
| TrypLExpress | Gibco | 12604-021 | Cell dissociation reagent |
| Wave controller software | Agilent Technologies Inc. | - | |
| Wide orifice tip | STARLAB International GmbH | E1011-8400 | Pipette tips with wide opening for spheroid handling |
References
- Correa de Sampaio, P., et al. A heterogeneous in vitro three dimensional model of tumour-stroma interactions regulating sprouting angiogenesis. PLoS One. 7 (2), 30753 (2012).
- Amann, A., et al. Development of an innovative 3D cell culture system to study tumour-stroma interactions in non-small cell lung cancer cells. PLoS One. 9 (3), 92511 (2014).
- Russell, S., Wojtkowiak, J., Neilson, A., Gillies, R. J. Metabolic profiling of healthy and cancerous tissues in 2D and 3D. Scientific Reports. 7 (1), 15285 (2017).
- Zanoni, M., et al. 3D tumor spheroid models for in vitro therapeutic screening: a systematic approach to enhance the biological relevance of data obtained. Scientific Reports. 6, 19103 (2016).
- Song, Y., et al. Patient-derived multicellular tumor spheroids towards optimized treatment for patients with hepatocellular carcinoma. Journal of Experimental and Clinica Cancer Research. 37 (1), 109 (2018).
- Courau, T., et al. Cocultures of human colorectal tumor spheroids with immune cells reveal the therapeutic potential of MICA/B and NKG2A targeting for cancer treatment. Journal for ImmunoTherapy of Cancer. 7 (1), 74 (2019).
- Ivanova, E., et al. Use of ex vivo patient-derived tumor organotypic spheroids to identify combination therapies for HER2 mutant non-small cell lung cancer. Clinical Cancer Research. 26 (10), 2393-2403 (2020).
- Mandon, M., Huet, S., Dubreil, E., Fessard, V., Le Hegarat, L. Three-dimensional HepaRG spheroids as a liver model to study human genotoxicity in vitro with the single cell gel electrophoresis assay. Scientific Reports. 9 (1), 10548 (2019).
- Stampar, M., et al. Hepatocellular carcinoma (HepG2/C3A) cell-based 3D model for genotoxicity testing of chemicals. Science of the Total Environment. 755, 143255 (2020).
- Coltman, N. J., et al. Application of HepG2/C3A liver spheroids as a model system for genotoxicity studies. Toxicology Letters. 345, 34-45 (2021).
- Tchoryk, A., et al. Penetration and uptake of nanoparticles in 3D tumor spheroids. Bioconjugate Chemistry. 30 (5), 1371-1384 (2019).
- Leite, P. E. C., et al. Suitability of 3D human brain spheroid models to distinguish toxic effects of gold and poly-lactic acid nanoparticles to assess biocompatibility for brain drug delivery. Partical Fibre Toxicology. 16 (1), 22 (2019).
- Elje, E., et al. Hepato(Geno)toxicity assessment of nanoparticles in a HepG2 liver spheroid model. Nanomaterials. 10 (3), 545 (2020).
- Conway, G. E., et al. Adaptation of the in vitro micronucleus assay for genotoxicity testing using 3D liver models supporting longer-term exposure durations. Mutagenesis. 35 (4), 319-330 (2020).
- Wang, Z., et al. HepaRG culture in tethered spheroids as an in vitro three-dimensional model for drug safety screening. Journal of Applied Toxicology. 35 (8), 909-917 (2015).
- Proctor, W. R., et al. Utility of spherical human liver microtissues for prediction of clinical drug-induced liver injury. Archives of Toxicology. 91 (8), 2849-2863 (2017).
- Basharat, A., Rollison, H. E., Williams, D. P., Ivanov, D. P. HepG2 (C3A) spheroids show higher sensitivity compared to HepaRG spheroids for drug-induced liver injury (DILI). Toxicology and Applied Pharmacology. 408, 115279 (2020).
- Benning, L., Peintner, A., Finkenzeller, G., Peintner, L. Automated spheroid generation, drug application and efficacy screening using a deep learning classification: a feasibility study. Scientific Reports. 10 (1), 11071 (2020).
- Mittler, F., et al. High-content monitoring of drug effects in a 3D spheroid model. Frontiers in Oncology. 7, 293 (2017).
- Brand, M. D., Nicholls, D. G.
- Benz, R., McLaughlin, S. The molecular mechanism of action of the proton ionophore FCCP (carbonylcyanide p-trifluoromethoxyphenylhydrazone). Biophysical Journal. 41 (3), 381-398 (1983).
- Kasianowicz, J., Benz, R., McLaughlin, S. The kinetic mechanism by which CCCP (carbonyl cyanide m-chlorophenylhydrazone) transports protons across membranes. The Journal of Membrane Biology. 82 (2), 179-190 (1984).
- Kenwood, B. M., et al. Identification of a novel mitochondrial uncoupler that does not depolarize the plasma membrane. Molecular Metabolism. 3 (2), 114-123 (2013).
- Mitchell, P. Coupling of phosphorylation to electron and hydrogen transfer by a chemi-osmotic type of mechanism. Nature. 191, 144-148 (1961).
- Alexopoulos, S. J., et al. Mitochondrial uncoupler BAM15 reverses diet-induced obesity and insulin resistance in mice. Nature Communications. 11 (1), 2397 (2020).
- Chen, S. -Y., et al. Mitochondrial uncoupler SHC517 reverses obesity in mice without affecting food intake. Metabolism - Clinical and Experimental. 117, 154724 (2021).
- Goedeke, L., Shulman, G. I. Therapeutic potential of mitochondrial uncouplers for the treatment of metabolic associated fatty liver disease and NASH. Molecular Metabolism. 46, 101178 (2021).
- Hill, B. G., et al. Integration of cellular bioenergetics with mitochondrial quality control and autophagy. Biological chemistry. 393 (12), 1485-1512 (2012).
- Demine, S., Renard, P., Arnould, T. Mitochondrial uncoupling: a key controller of biological processes in physiology and diseases. Cells. 8 (8), 795 (2019).
- Wang, J., et al. Uncoupling effect of F16 is responsible for its mitochondrial toxicity and anticancer activity. Toxicological Sciences. 161 (2), 431-442 (2018).
- Tretter, L., Chinopoulos, C., Adam-Vizi, V. Plasma membrane depolarization and disturbed Na+ homeostasis induced by the protonophore carbonyl cyanide-p-trifluoromethoxyphenyl-hydrazon in isolated nerve terminals. Molecular Pharmacology. 53 (4), 734-741 (1998).
- Connop, B. P., Thies, R. L., Beyreuther, K., Ida, N., Reiner, P. B. Novel effects of FCCP [carbonyl cyanide p-(trifluoromethoxy) phenylhydrazone] on amyloid precursor protein processing. Journal of neurochemistry. 72 (4), 1457-1465 (1999).
- Stöckl, P., et al. Partial uncoupling of oxidative phosphorylation induces premature senescence in human fibroblasts and yeast mother cells. Free Radical Biology and Medicine. 43 (6), 947-958 (2007).
- Firsov, A. M., et al. Protonophoric action of BAM15 on planar bilayers, liposomes, mitochondria, bacteria and neurons. Bioelectrochemistry. 137, 107673 (2021).
- Dranka, B. P., Hill, B. G., Darley-Usmar, V. M. Mitochondrial reserve capacity in endothelial cells: The impact of nitric oxide and reactive oxygen species. Free Radical Biology and Medicine. 48 (7), 905-914 (2010).
- Eilenberger, C., Rothbauer, M., Ehmoser, E. K., Ertl, P., Kupcu, S. Effect of spheroidal age on sorafenib diffusivity and toxicity in a 3D HepG2 spheroid model. Scientific Reports. 9 (1), 4863 (2019).
- vanden Brand, D., Veelken, C., Massuger, L., Brock, R. Penetration in 3D tumor spheroids and explants: Adding a further dimension to the structure-activity relationship of cell-penetrating peptides. Biochimica et Biophysica Acta (BBA) - Biomembranes. 1860 (6), 1342-1349 (2018).
- Niora, M., et al. Head-to-head comparison of the penetration efficiency of lipid-based nanoparticles into tumor spheroids. ACS Omega. 5 (33), 21162-21171 (2020).
- Millard, M., et al. Drug delivery to solid tumors: the predictive value of the multicellular tumor spheroid model for nanomedicine screening. International Journal of Nanomedicine. 12, 7993-8007 (2017).
- Ruas, J. S., et al. Underestimation of the maximal capacity of the mitochondrial electron transport system in oligomycin-treated cells. PLoS One. 11 (3), 0150967 (2016).
- Benton, G., DeGray, G., Kleinman, H. K., George, J., Arnaoutova, I. In vitro microtumors provide a physiologically predictive tool for breast cancer therapeutic screening. PLoS One. 10 (4), 0123312 (2015).
- Hirpara, J., et al. Metabolic reprogramming of oncogene-addicted cancer cells to OXPHOS as a mechanism of drug resistance. Redox Biology. 25, 101076 (2019).
- Ware, M. J., et al. Generation of homogenous three-dimensional pancreatic cancer cell spheroids using an improved hanging drop technique. Tissue Engineering. Part C, Methods. 22 (4), 312-321 (2016).
- Song, Y., et al. TGF-β-independent CTGF induction regulates cell adhesion mediated drug resistance by increasing collagen I in HCC. Oncotarget. 8 (13), 21650-21662 (2017).
- Wrzesinski, K., et al. HepG2/C3A 3D spheroids exhibit stable physiological functionality for at least 24 days after recovering from trypsinisation. Toxicology Research. 2 (3), 163-172 (2013).
- Gaskell, H., et al. Characterization of a functional C3A liver spheroid model. Toxicology Research. 5 (4), 1053-1065 (2016).
- Takahashi, Y., et al. 3D spheroid cultures improve the metabolic gene expression profiles of HepaRG cells. Bioscience Reports. 35 (3), 00208 (2015).
- Hendriks, D. F. G., Puigvert, L. F., Messner, S., Mortiz, W., Ingelman-Sundberg, M. Hepatic 3D spheroid models for the detection and study of compounds with cholestatic liability. Scientific Reports. 6, 35434 (2016).
- Leung, B. M., Lesher-Perez, S. C., Matsuoka, T., Moraes, C., Takayama, S. Media additives to promote spheroid circularity and compactness in hanging drop platform. Biomaterials Science. 3 (2), 336-344 (2015).
- Cavo, M., et al. A synergic approach to enhance long-term culture and manipulation of MiaPaCa-2 pancreatic cancer spheroids. Scientific Reports. 10 (1), 10192 (2020).
- Carlsson, J., Yuhas, J. M.
- Costa, E. C., Gaspar, V. M., Coutinho, P., Correia, I. J. Optimization of liquid overlay technique to formulate heterogenic 3D co-cultures models. Biotechnology and Bioengineering. 111 (8), 1672-1685 (2014).
- Lundholt, B. K., Scudder, K. M., Pagliaro, L. A simple technique for reducing edge effect in cell-based assays. Journal of Biomolecular Screening. 8 (5), 566-570 (2003).
- Zhang, X. D., et al. The use of strictly standardized mean difference for hit selection in primary RNA interference high-throughput screening experiments. Journal of Biomolecular Screening. 12 (4), 497-509 (2007).
- Yepez, V. A., et al. OCR-Stats: Robust estimation and statistical testing of mitochondrial respiration activities using Seahorse XF Analyzer. PLoS One. 13 (7), 0199938 (2018).
- Silva, L. P., et al. Measurement of DNA concentration as a normalization strategy for metabolomic data from adherent cell lines. Analytical Chemistry. 85 (20), 9536-9542 (2013).
Tags
암 연구 문제 180Erratum
Formal Correction: Erratum: Exploring Mitochondrial Energy Metabolism of Single 3D Microtissue Spheroids using Extracellular Flux Analysis
Posted by JoVE Editors on 03/11/2022.
Citeable Link.
An erratum was issued for: Exploring Mitochondrial Energy Metabolism of Single 3D Microtissue Spheroids using Extracellular Flux Analysis. The Representative Results section was updated.
Figure 5 was updated from:
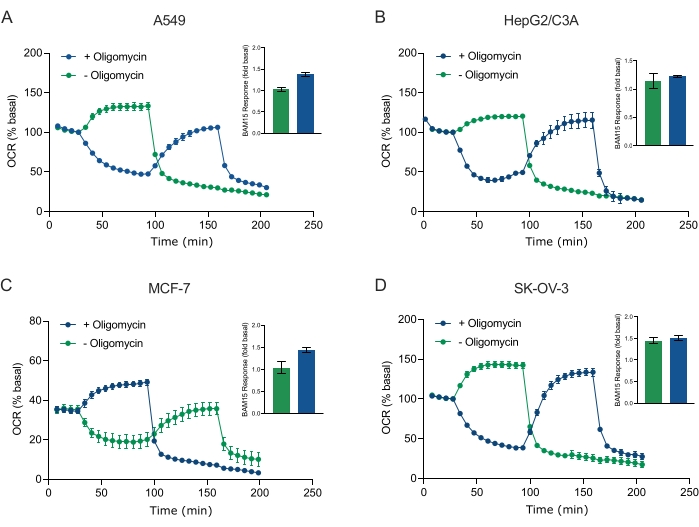
Figure 5: Single or sequential injection of mitochondrial respiratory compounds. Cancer-cell-derived spheroids of MCF-7, HEPG2/C3A, SK-OV-3, and A549 were placed into wells of an XFe96 spheroid microplate in XF RPMI and probed for OCR using the Agilent Seahorse XFe96 analyzer. OCR was measured 5x, after which 2 µg/mL oligomycin (injection Port A: green trace) or 5 µM BAM15 (injection Port A: blue trace or injection port B: green trace) to inhibit the mitochondrial ATP synthase and determine maximal respiratory capacity, respectively. Kinetic OCR data are expressed as % basal (A-D). Maximal respiratory capacity (OCRmax) was calculated as a factor of basal OCR by the equation: OCRmax = OCRBAM15 / OCRbasal. OCRmax was obtained from OCR averages across measurement cycles 8-10 post BAM15 injection with (green bars) and without (blue bars) oligomycin. Data are averages ± SEM from 3-8 individual well replicates across the spheroid assay microplate. Abbreviations: OCR = oxygen consumption rate. Please click here to view a larger version of this figure.
to:
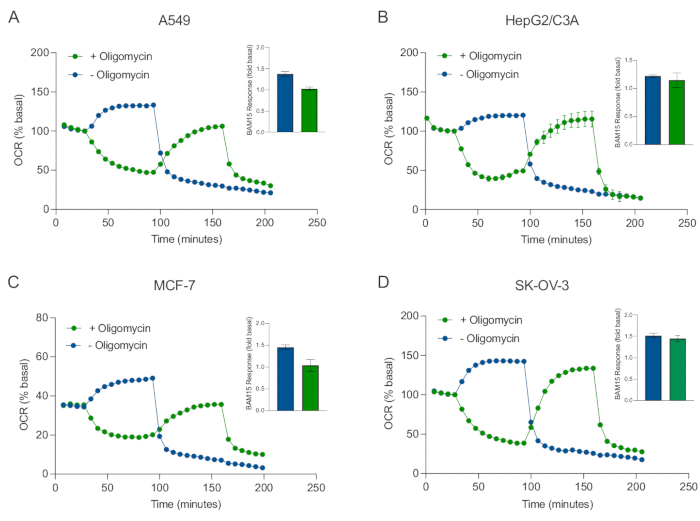
Figure 5: Single or sequential injection of mitochondrial respiratory compounds. Cancer-cell-derived spheroids of MCF-7, HEPG2/C3A, SK-OV-3, and A549 were placed into wells of an XFe96 spheroid microplate in XF RPMI and probed for OCR using the Agilent Seahorse XFe96 analyzer. OCR was measured 5x, after which 2 µg/mL oligomycin (injection Port A: green trace) or 5 µM BAM15 (injection Port A: blue trace or injection port B: green trace) to inhibit the mitochondrial ATP synthase and determine maximal respiratory capacity, respectively. Kinetic OCR data are expressed as % basal (A-D). Maximal respiratory capacity (OCRmax) was calculated as a factor of basal OCR by the equation: OCRmax = OCRBAM15 / OCRbasal. OCRmax was obtained from OCR averages across measurement cycles 8-10 post BAM15 injection with (green bars) and without (blue bars) oligomycin. Data are averages ± SEM from 3-8 individual well replicates across the spheroid assay microplate. Abbreviations: OCR = oxygen consumption rate. Please click here to view a larger version of this figure.
