

ERRATUM NOTICE
Important: There has been an erratum issued for this article. Read more …
Summary
Ces protocoles aideront les utilisateurs à sonder le métabolisme énergétique mitochondrial dans les sphéroïdes dérivés de lignées cellulaires cancéreuses 3D à l’aide de l’analyse du flux extracellulaire Seahorse.
Abstract
Les agrégats cellulaires tridimensionnels (3D), appelés sphéroïdes, sont devenus l’avant-garde de la culture cellulaire in vitro ces dernières années. Contrairement à la culture de cellules en tant que monocouches unicellulaires bidimensionnelles (culture 2D), la culture de cellules sphéroïdes favorise, régule et soutient l’architecture cellulaire physiologique et les caractéristiques qui existent in vivo, y compris l’expression des protéines de matrice extracellulaire, la signalisation cellulaire, l’expression génique, la production de protéines, la différenciation et la prolifération. L’importance de la culture 3D a été reconnue dans de nombreux domaines de recherche, notamment l’oncologie, le diabète, la biologie des cellules souches et le génie tissulaire. Au cours de la dernière décennie, des méthodes améliorées ont été développées pour produire des sphéroïdes et évaluer leur fonction métabolique et leur devenir.
Les analyseurs de flux extracellulaire (XF) ont été utilisés pour explorer la fonction mitochondriale dans des microtissus 3D tels que les sphéroïdes à l’aide d’une plaque de capture d’îlot XF24 ou d’une microplaque sphéroïde XFe96. Cependant, des protocoles distincts et l’optimisation du métabolisme énergétique mitochondrial sondant chez les sphéroïdes à l’aide de la technologie XF n’ont pas été décrits en détail. Cet article fournit des protocoles détaillés pour sonder le métabolisme énergétique mitochondrial dans des sphéroïdes 3D uniques à l’aide de microplaques sphéroïdes avec l’analyseur XFe96 XF. En utilisant différentes lignées cellulaires cancéreuses, il est démontré que la technologie XF est capable de distinguer la respiration cellulaire dans les sphéroïdes 3D de différentes tailles, mais aussi de différents volumes, numéros de cellules, contenu et type d’ADN.
Les concentrations optimales de composé effecteur mitochondrial d’oligomycine, de BAM15, de roténone et d’antimycine A sont utilisées pour sonder des paramètres spécifiques du métabolisme énergétique mitochondrial dans les sphéroïdes 3D. Cet article traite également des méthodes permettant de normaliser les données obtenues à partir de sphéroïdes et aborde de nombreuses considérations qui devraient être prises en compte lors de l’exploration du métabolisme des sphéroïdes à l’aide de la technologie XF. Ce protocole aidera à stimuler la recherche sur des modèles avancés de sphéroïdes in vitro .
Introduction
Les progrès des modèles in vitro dans la recherche biologique ont progressé rapidement au cours des 20 dernières années. Ces modèles incluent désormais des modalités d’organe sur puce, des organoïdes et des sphéroïdes de microtissus 3D, qui sont tous devenus un objectif commun pour améliorer la traduction entre les études in vitro et in vivo. L’utilisation de modèles in vitro avancés, en particulier les sphéroïdes, couvre plusieurs domaines de recherche, notamment l’ingénierie tissulaire, la recherche sur les cellules souches, le cancer et la biologie des maladies 1,2,3,4,5,6,7, et les tests de sécurité, y compris la toxicologie génétique 8,9,10, la toxicologie des nanomatériaux 11, 12,13,14, et tests d’innocuité et d’efficacité des médicaments 8,15,16,17,18,19.
La morphologie cellulaire normale est essentielle au phénotype et à l’activité biologiques. Cultiver des cellules en sphéroïdes de microtissus 3D permet aux cellules d’adopter une morphologie, une fonction phénotypique et une architecture, plus proches de celles observées in vivo mais difficiles à capturer avec les techniques classiques de culture cellulaire monocouche. Tant in vivo qu’in vitro, la fonction cellulaire est directement affectée par le microenvironnement cellulaire, qui ne se limite pas à la communication et à la programmation cellulaires (p. ex., formations de jonctions cellule-cellule, possibilités de former des niches cellulaires); l’exposition cellulaire aux hormones et aux facteurs de croissance dans les environnements immédiats (p. ex., exposition aux cytokines cellulaires dans le cadre d’une réponse inflammatoire); la composition des matrices physiques et chimiques (p. ex., si les cellules sont cultivées dans un plastique de culture tissulaire rigide ou dans un environnement tissulaire élastique); et surtout, comment le métabolisme cellulaire est affecté par la nutrition et l’accès à l’oxygène ainsi que par le traitement des déchets métaboliques tels que l’acide lactique.
L’analyse du flux métabolique est un moyen puissant d’examiner le métabolisme cellulaire dans des systèmes in vitro définis. Plus précisément, la technologie XF permet d’analyser les changements en direct et en temps réel dans la bioénergétique cellulaire des cellules et des tissus intacts. Étant donné que de nombreux événements métaboliques intracellulaires se produisent dans l’ordre de quelques secondes à quelques minutes, les approches fonctionnelles en temps réel sont primordiales pour comprendre les changements en temps réel du flux métabolique cellulaire dans les cellules et les tissus intacts in vitro.
Cet article fournit des protocoles pour la culture des lignées cellulaires dérivées du cancer A549 (adénocarcinome pulmonaire), HepG2/C3A (carcinome hépatocellulaire), MCF-7 (adénocarcinome du sein) et SK-OV-3 (adénocarcinome ovarien) en tant que modèles sphéroïdes 3D in vitro utilisant des approches d’agrégation forcée (Figure 1). Il décrit également en détail comment sonder le métabolisme énergétique mitochondrial de sphéroïdes 3D uniques à l’aide de l’analyseur Agilent XFe96 XF, (ii) met en évidence les moyens d’optimiser les tests XF à l’aide de sphéroïdes 3D uniques, et (iii) discute des considérations et des limites importantes de la sonde du métabolisme des sphéroïdes 3D à l’aide de cette approche. Plus important encore, cet article décrit comment sont collectés des ensembles de données qui permettent le calcul du taux de consommation d’oxygène (OCR) pour déterminer la phosphorylation oxydative et donc la fonction mitochondriale dans les sphéroïdes cellulaires. Bien qu’il ne soit pas analysé pour ce protocole, le taux d’acidification extracellulaire (ECAR) est un autre paramètre mesuré aux côtés des données OCR dans les expériences XF. Cependant, ECAR est souvent mal ou mal interprété à partir des jeux de données XF. Nous fournissons un commentaire sur les limites du calcul de l’ECAR en suivant les approches de base du fabricant de la technologie.
Subscription Required. Please recommend JoVE to your librarian.
Protocol

Figure 1 : Flux de travail graphique pour la génération de sphéroïdes cellulaires, l’analyse du flux extracellulaire et les essais en aval. Quatre lignées cellulaires cancéreuses ont été cultivées sélectivement sous forme de monocouches (A), détachées de flacons de culture tissulaire et ensemencées dans des microplaques ultrabasses de 96 puits pour former des sphéroïdes (B). Le carcinome pulmonaire A549, le carcinome hépatique HepG2/C3A, l’adénocarcinome ovarien SK-OV-3 et le carcinome du sein MCF-7 ont été ensemencés à 1 × 103-8 × 103 cellules/ puits et ont grandi jusqu’à 7 jours pour former des sphéroïdes uniques et optimiser la densité d’ensemencement des sphéroïdes et le temps de culture par observation continue et mesures planimétriques. Une fois formés, les sphéroïdes simples ont été lavés dans un milieu XF sans sérum et soigneusement ensemencés dans des microplaques de dosage sphéroïde, pré-enduites de poly-D-lysine (C). Les sphéroïdes ont été soumis à une analyse du flux extracellulaire à l’aide de l’analyseur XFe96 utilisant plusieurs protocoles pour traiter: (1) la taille optimale des sphéroïdes pour la réponse respiratoire mitochondriale basale; (2) titrage optimisé des inhibiteurs respiratoires mitochondriaux; (3) optimisation du placement des sphéroïdes dans les puits de microplaques. (D) Les analyses post-XF, la microscopie à contraste de phase et la quantification de l’ADN sphéroïde ont été utilisées pour la normalisation des données et d’autres essais in vitro en aval. Veuillez cliquer ici pour voir une version agrandie de cette figure.
1. Culture de lignées cellulaires cancéreuses sous forme de sphéroïdes in vitro 3D
| Lignée cellulaire | Description | Milieu de culture | Source |
| L’A549 | Lignée cellulaire de carcinome pulmonaire | RPMI 1640 | Collection européenne de cultures cellulaires authentifiées (ECACC) |
| Pyruvate de sodium (1 mM) | |||
| Pénicilline- Streptomycine - (100 U/mL – 100 mg/mL) | |||
| 10 % (v/v) FBS | |||
| HepG2/C3A | Lignée cellulaire de carcinome hépatique, un dérivé clonal de la lignée cellulaire parente HepG2 | Le | Collection américaine de culture tissulaire (ATCC) |
| Pénicilline- Streptomycine - (100 U/mL – 100 mg/mL) | |||
| 10 % (v/v) FBS | |||
| MCF7 | Lignée cellulaire d’adénocarcinome du sein | RPMI 1640 | Collection européenne de cultures cellulaires authentifiées (ECACC) |
| Pyruvate de sodium (1 mM) | |||
| Pénicilline- Streptomycine - (100 U/mL – 100 mg/mL) | |||
| 10 % (v/v) FBS | |||
| SK-OV-3 | Lignée cellulaire de l’adénocarcinome ovarien | RPMI 1640 | Collection européenne de cultures cellulaires authentifiées (ECACC) |
| Pyruvate de sodium (1 mM) | |||
| Pénicilline- Streptomycine - (100 U/mL – 100 mg/mL) | |||
| 10 % (v/v) FBS | |||
| Composant | Milieu d’essai RPMI (volume final de 50 mL) | ||
| Milieu de base | Agilent Seahorse XF RPMI, pH 7,4 | ||
| Glucose (1 M de stock stérile) | 11 mM (solution mère de 0,55 mL) | ||
| L-glutamine (stock stérile de 200 mM) | 2 mM (0,5 mL de solution mère) | ||
| Pyruvate de sodium (stock stérile de 100 mM) | 1 mM (0,5 mL de solution mère) |
Tableau 1 : Compositions des milieux de lignées cellulaires cancéreuses et des milieux XF.
- Cultivez toutes les lignées cellulaires à l’aide de la technique de culture tissulaire aseptique standard et confirmez qu’elles sont exemptes de mycoplasmes à l’aide d’un kit de dosage approprié.
- Cultiver les lignées cellulaires dans des flacons de culture tissulaire T75 ou l’équivalent, en utilisant le milieu recommandé (tableau 1). Cultivez les lignées cellulaires à 65-80% de confluence et passez-les régulièrement jusqu’à un maximum de 25 passages.
- Rincez les flacons de culture cellulaire deux fois dans la solution saline tamponnée au phosphate (DBPS) modifiée de Dulbecco.
- Détacher les cellules des flacons avec 3 mL de réactif de dissociation cellulaire (voir le tableau des matériaux) pendant 5 min à 37 °C et confirmer le détachement par microscopie.
- Aspirer doucement la suspension cellulaire détachée pour assurer une suspension unicellulaire et désactiver le réactif de dissociation cellulaire avec 7 mL de milieu de culture tissulaire complet.
- Prélever les cellules par centrifugation à 300 × g pendant 5 min, jeter le surnageant et remettre les cellules en suspension dans un milieu complet.
- Comptez les cellules à l’aide d’un hémocytomètre ou d’un compteur cellulaire automatisé et titrez à la densité cellulaire souhaitée requise pour l’ensemencement.
REMARQUE: Pour ensemencer une plaque entière de 96 puits à 100 μL / puits à 4 × 103 cellules / puits, les cellules doivent être titrées à 4 × 104 cellules / mL dans un volume recommandé de 12 mL. - Décanter la suspension cellulaire dans un réservoir stérile et distribuer 100 μL de la suspension cellulaire dans chaque puits d’une microplaque répulsive cellulaire à l’aide d’un pipettor multicanal.
REMARQUE: Seuls les 60 puits intérieurs d’une microplaque doivent être ensemencés et le reste rempli de DPBS. Cela formera une barrière d’évaporation, assurera l’homogénéité des sphéroïdes sur la plaque et minimisera les effets de bord de la plaque. - Centrifuger les microplaques sphéroïdes à 300 × g pendant 15 min pour forcer les cellules en agrégats lâches.
- Incuber les plaques à 37 °C, 5% de CO2 pendant au moins 3 jours pour assurer la formation de sphéroïdes.
- Effectuer une microscopie à contraste de phase en utilisant des pratiques de laboratoire normalisées pour surveiller la croissance des sphéroïdes. Reconstituer le milieu de culture cellulaire tous les 3 jours ou deux fois par semaine en effectuant un échange de milieu d’un demi-volume.
2. Sonder le métabolisme énergétique mitochondrial des sphéroïdes simples à l’aide de la technologie de flux extracellulaire (XF)
- Préparation du test (un jour avant)
- Vérifiez la viabilité des sphéroïdes à l’aide d’un microscope à lumière inversée avec contraste de phase à un grossissement de 4x pour assurer une structure sphéroïde intacte, une morphologie et une uniformité globale entre les échantillons.
- Hydratez la cartouche du capteur.
- Aliquote ~20 mL du calibrant dans un tube conique.
- Placer le tube conique contenant l’étrier dans un incubateur sans CO2 37 °C pendant la nuit.
- Retirez le contenu du kit d’analyse.
- Retirez la cartouche du capteur de la plaque utilitaire et placez-la à l’envers sur le plan de travail à côté de la plaque utilitaire.
- Pipette 200 μL de ddH2O stérile dans chaque puits de la plaque d’utilité de la cartouche du capteur à l’aide d’une pipette P300 multicanal.
- Placez la cartouche du capteur sur le dessus de la plaque utilitaire.
- Vérifiez que le niveau d’eau dans chaque puits est suffisamment élevé pour submerger les sondes du capteur.
- Transférez la cartouche de capteur assemblée dans un incubateur sans CO2 37 °C et laissez-la toute la nuit.
REMARQUE: Cette étape peut être effectuée 12 à 72 heures avant le début du test.
- Microplaque de dosage sphéroïde de couche
- À l’aide de techniques aseptiques, ajouter 30 μL/puits de poly-D-lysine stérile (0,1 mg/mL) à la microplaque sphéroïde et l’incuber pendant 30 min à température ambiante.
- Aspirer la solution de chaque puits de la microplaque sphéroïde, inverser la plaque et la tapoter fermement sur le papier de soie pour éliminer toute solution résiduelle.
- Laver la plaque deux fois avec 200 μL/puits de ddH2O stérile.
- Après le lavage final, inverser la microplaque et la tapoter fermement sur du papier de soie pour éliminer toute eau résiduelle.
- Laisser sécher la plaque à l’air libre pendant 30 min avant de l’utiliser ou de la stocker à 4 °C pour une utilisation future.
REMARQUE: La microplaque de dosage des sphéroïdes doit être recouverte d’un adhésif moléculaire pour s’assurer que les sphéroïdes sont fixés au bas de la microplaque. Sans adhésif moléculaire, les sphéroïdes peuvent se déloger et interférer avec les résultats des tests. D’autres adhésifs moléculaires peuvent également être utilisés comme alternative à la poly-D-lysine pour les plaques de prérevêtement. Les plaques prélaquées peuvent être conservées à 4 °C, mais doivent être laissées à la température ambiante avant le début du dosage.
- Préparer le milieu du test XF
- Préparer le milieu XF RPMI, comme indiqué dans le tableau 1, et le filtre stérile avec un filtre à seringue de 0,22 μm
- Préparation du test (1 h avant le test)
- Préchauffer le milieu d’essai XF RPMI supplémenté jusqu’à 37 °C.
- Préchauffer la microplaque de dosage sphéroïde revêtue dans un incubateur ou un bain sec sans CO2 à 37 °C.
- Préparez la cartouche du capteur.
- Retirez le tube conique contenant l’étrier et la cartouche du capteur de l’incubateur à air.
- Retirez la cartouche du capteur de la plaque utilitaire et placez-la à l’envers sur la surface de travail.
- À l’aide d’une pipette multicanal P300, aspirez l’eau de la plaque de service public et jetez-la.
- Versez la solution d’étriquant dans un réservoir de réactif stérile et ajoutez 200 μL/puits du calibrant préavertissé à la plaque utilitaire à l’aide d’une pipette multicanal P300.
- Prenez la cartouche du capteur et replacez-la sur la plaque d’utilité, en vous assurant que les capteurs sont bien immergés dans l’étrier.
- Transférer la cartouche de capteur assemblée dans l’incubateur sans CO2 37 °C jusqu’à ce qu’elle soit prête à charger les solutions d’injection de orifice.
- Lavez les sphéroïdes avec le milieu d’essai.
- Retirez la plaque de culture sphéroïde de l’incubateur à 37 °C, 5 % de CO2 et observez les sphéroïdes au microscope pour assurer leur intégrité avant les étapes de transfert des sphéroïdes.
- Charger tous les puits de la plaque sphéroïde avec 180 μL/puits de milieu d’essai préavertis, y compris tous les puits de correction de fond.
- Remplissez partiellement une boîte de Petri de 7 cm avec 3 mL du milieu d’essai.
- À l’aide d’une pipette multicanal chargée de larges pointes de pipette à orifice, transférez les sphéroïdes de la plaque de culture à 96 puits dans des boîtes de Petri de 7 cm en réglant le pipetteur à un volume d’aspiration de 10 à 50 μL.
- Ensemencez les sphéroïdes dans la microplaque de dosage des sphéroïdes pré-enduits.
- À l’aide d’un microscope à dissection et d’un appareil lightbox, transférez les sphéroïdes de la boîte de Petri à la microplaque de dosage des sphéroïdes, comme indiqué ci-dessous.
- Réglez le volume d’un pipetteur monocanal équipé d’une large pointe de pipette à orifice à 20 μL et aspirez soigneusement un seul sphéroïde. Placez la pointe directement au centre de chaque puits de la microplaque d’essai sphéroïde et laissez la gravité éluer un seul sphéroïde au centre de chaque puits, c’est-à-dire n’expulsez aucun milieu de la pointe de la pipette et laissez l’action capillaire retirer le sphéroïde de la pointe de la pipette. Pour confirmer l’élution, le contenu du pipetteur peut être pipeté dans la boîte de Petri de 7 cm au microscope.
REMARQUE: L’élution par gravité d’un seul sphéroïde prend généralement 15 à 30 s en fonction de la taille / densité du sphéroïde. Pendant ce temps, le pipetteur ne doit pas être retiré. Tout puits de correction de fond doit être exempt de sphéroïdes et ne contenir que du milieu d’essai. Sous le microscope, confirmez la position de chaque sphéroïde. Chaque sphéroïde devrait idéalement être positionné au centre de chaque puits. - Une fois que tous les sphéroïdes ont été transférés sur la microplaque d’essai des sphéroïdes, transférer la plaque dans un incubateur sans CO2 à 37 °C pendant au moins 1 h avant le test.
- Réglez le volume d’un pipetteur monocanal équipé d’une large pointe de pipette à orifice à 20 μL et aspirez soigneusement un seul sphéroïde. Placez la pointe directement au centre de chaque puits de la microplaque d’essai sphéroïde et laissez la gravité éluer un seul sphéroïde au centre de chaque puits, c’est-à-dire n’expulsez aucun milieu de la pointe de la pipette et laissez l’action capillaire retirer le sphéroïde de la pointe de la pipette. Pour confirmer l’élution, le contenu du pipetteur peut être pipeté dans la boîte de Petri de 7 cm au microscope.
- À l’aide d’un microscope à dissection et d’un appareil lightbox, transférez les sphéroïdes de la boîte de Petri à la microplaque de dosage des sphéroïdes, comme indiqué ci-dessous.
3. Préparation et chargement des composés dans la cartouche du capteur pour les tests XF
| Stratégie d’injection | Composé (port) | Volume de démarrage du micropuit XFe96 (μL) | Concentration finale souhaitée dans le puits | Volume du port (μL) | Volume final du micropuit XFe96 après injection (μL) | Concentration du stock de travail |
| 1 | Oligomycine (A) | 180 | 3 ug/mL | 20 | 200 | 30 μg/mL |
| Roténone (B) | 200 | 2 μM | 20 | 220 | 22 μM | |
| Antimycine A (B) | 200 | 2 μM | 20 | 220 | 22 μM | |
| 2 | BAM15 (A) | 180 | 5 μM | 20 | 200 | 50 μM |
| Roténone (B) | 200 | 2 μM | 20 | 220 | 22 μM | |
| Antimycine A (B) | 200 | 2 μM | 20 | 220 | 22 μM |
Tableau 2 : Concentrations de composés mitochondriaux pour sonder le métabolisme énergétique mitochondrial de sphéroïdes 3D uniques à l’aide de l’analyseur XFe96.
- Préparer les concentrations sur le stock de travail de chaque composé, comme indiqué dans le tableau 2 , à l’aide d’un milieu d’essai XF RPMI préavertissé entièrement complété.
- Orientez la plaque de cartouche (couplée à la plaque utilitaire) en colonne, 1-12 de gauche à droite.
- Si vous utilisez un guide de chargement, placez-le sur la plaque de la cartouche conformément à la procédure de chargement du puits, par exemple, si le port A est chargé en premier, assurez-vous que A est visible dans le coin supérieur gauche du guide.
- Transférer la solution de travail de chaque composé dans un réservoir approprié et, à l’aide d’une pipette multicanal P100 étalonnée, distribuer 20 μL dans tous les ports correspondants. Répétez l’opération pour chaque composé dans les ports restants.
REMARQUE: Si aucun port n’est utilisé sur la plaque de la cartouche du capteur, ceux-ci peuvent être laissés vides ou remplis de support de dosage. Si seule une sélection d’une lettre de port spécifique est utilisée, assurez-vous que les autres ports correspondant à cette lettre sont chargés avec un support d’essai; sinon, de l’air sera injecté dans le puits, compromettant les résultats dans ces puits. - Après le chargement du port, retirez les guides de chargement de la plaque (le cas échéant) et préparez l’analyseur pour le chargement de la cartouche du capteur.
REMARQUE: Si le test n’est pas exécuté immédiatement après le chargement des ports, replacez le couvercle sur la cartouche du capteur et remettez la plaque dans l’incubateur à air à 37 ° C jusqu’à ce qu’elle soit prête à charger dans la machine.
4. Conception des essais, stratégies d’injection et acquisition de données
- Exécution du test
- Mettez l’analyseur sous tension et connectez-le au contrôleur (ordinateur).
REMARQUE: Cela peut être vérifié par l’état de la connexion de l’instrument dans le panneau de widget du logiciel Wave Controller. - Accédez à la page des modèles dans le logiciel WAVE, recherchez le fichier de modèle de test de l’expérience et double-cliquez dessus pour l’ouvrir.
Remarque : Si le modèle de test n’apparaît pas dans la vue Modèles , importez le fichier de modèle dans le dossier de modèle à partir d’un lecteur réseau partagé ou d’un lecteur flash USB. - Pour démarrer le test, cliquez sur l’onglet Exécuter le test .
REMARQUE: Si les définitions de groupe ont été correctement attribuées dans la carte des plaques, le test sera prêt à s’exécuter comme indiqué par la coche verte sur le côté droit de la page. À ce stade, toute information supplémentaire peut être saisie sur la page de résumé de l’essai ou sur la page laissée vide; passez à l’étape suivante. En raison de la pénétration retardée des modulateurs mitochondriaux dans les sphéroïdes de microtissus 3D (Figure 2), utilisez les informations du protocole de mesure décrites dans le tableau 3.
- Mettez l’analyseur sous tension et connectez-le au contrôleur (ordinateur).
| Période de mesure | Numéro d’injection et port | Détails de la mesure | Durée de la période (h:min:s) |
| Étalonnage | Sans objet | Les analyseurs XF effectuent toujours cet étalonnage pour s’assurer que les mesures sont précises | 00:20:00 (il s’agit d’une moyenne qui peut varier d’une machine à l’autre) |
| Équillibration | Sans objet | L’équilibrage a lieu après l’étalonnage et il est recommandé. | 00:10:00 |
| Basal | Sans objet | Cycles = 5 | 00:30:00 |
| Mélange = 3:00 | |||
| Attendre = 0:00 | |||
| Mesure = 3:00 | |||
| Oligomycine / BAM15 | Injection 1 (Port A) | Cycles = 10 | 01:00:00 |
| Mélange = 3:00 | |||
| Attendre = 0:00 | |||
| Mesure = 3:00 | |||
| Roténone + antimycine A | Injection 2 (Port B) | Cycles = 10 | 01:00:00 |
| Mélange = 3:00 | |||
| Attendre = 0:00 | |||
| Mesure = 3:00 | |||
| Temps total: | 03:00:00 | ||
Tableau 3 : Configuration du protocole pour sonder le métabolisme énergétique mitochondrial de sphéroïdes 3D uniques à l’aide de l’analyseur XFe96.
- Cliquez sur Démarrer pour afficher la boîte de dialogue Emplacement d’enregistrement .
- Entrez l’emplacement d’enregistrement du fichier de résultats et placez la cartouche de capteur assemblée sur le plateau thermique qui apparaît de la porte sur le côté de l’analyseur. Attendez que le plateau thermique s’ouvre automatiquement et que l’écran affiche le message Load Calibrant Utility Plate . Avant de suivre les instructions à l’écran, assurez-vous i) que la cartouche du capteur est bien ajustée sur la plaque utilitaire, ii) que le couvercle est retiré de la cartouche du capteur et iii) que la cartouche du capteur est correctement orientée sur la plaque utilitaire.
- Suivez les commandes à l’écran pour lancer l’étalonnage de la cartouche du capteur.
REMARQUE: Le temps nécessaire pour terminer l’étalonnage est d’environ 10-20 min (pour les essais à 37 ° C). - Après l’étalonnage de la cartouche du capteur, chargez la microplaque sphéroïde dans l’analyseur en suivant les instructions à l’écran sur le contrôleur d’ondes pour lancer l’étape d’équilibrage de 12 minutes.
REMARQUE: Les cases vertes avec des coches blanches indiquent un « bon » étalonnage pour ce puits. Si des puits ne fournissent pas un « bon » étalonnage, ils seront indiqués par une boîte rouge et une croix blanche. Ces puits doivent être notés et exclus de toute analyse une fois que l’essai est terminé à l’aide de l’onglet de l’essai de modification . - Attendez que l’analyseur commence automatiquement à acquérir des mesures de base une fois que la machine a terminé l’étape d’équilibrage (comme indiqué dans le protocole de l’instrument).
- Pour terminer l’expérience, suivez les commandes à l’écran sur le contrôleur WAVE.
REMARQUE: Une fois que la microplaque sphéroïde a été retirée de l’analyseur, jetez la cartouche du capteur et mettez de côté la plaque sphéroïde pour une analyse plus approfondie si nécessaire (par exemple, quantification de l’ADN double brin (ds)). Si la microplaque n’est pas nécessaire pour une analyse plus approfondie, elle peut être jetée avec la cartouche du capteur. - Attendez que la boîte de dialogue d’analyse s’affiche et affichez les résultats ou revenez à la vue des modèles .
5. Stratégies de normalisation et d’analyse des données - normalisation post-essai et essais en aval (étapes facultatives)
- Normalisation des données
- Pour normaliser les données sphéroïdes, reportez-vous à la série de protocoles pertinents pour les stratégies de normalisation des données pour calculer la taille et le volume des sphéroïdes et quantifier l’ADNds dans les essais sphéroïdes. Ceux-ci ont été inclus en tant que fichiers supplémentaires; voir Fichier supplémentaire 1 et Fichier supplémentaire 2.
- Analyse des données
- Pour exporter des données dans l’un des générateurs d’analyse automatisés, suivez les commandes d’exportation de données sur le contrôleur WAVE et sélectionnez le générateur d’exportation qui correspond au type de test. Vous pouvez également exporter le fichier de données et le télécharger dans Seahorse Analytics.
REMARQUE: L’inconvénient des générateurs de rapports et de l’analyse Seahorse est que l’analyse des données est limitée à la façon dont le test XF est conçu et ne permet pas de prendre des moyennes à travers les cycles de mesure. L’exportation manuelle des jeux de données à partir du logiciel de l’instrument permet à l’utilisateur de préférer à cet égard. Étant donné que la stratégie d’injection pour évaluer la respiration mitochondriale des sphéroïdes 3D sera probablement différente de celle d’un test typique « MitoStress », une série de modèles de feuilles de calcul ont été développés pour aider à analyser ces ensembles de données, spécifiques aux cultures cellulaires 3D et seront fournis sur demande. Ces fichiers de modèles de données fourniront des données sur les principaux paramètres respiratoires mitochondriaux détaillés et expliqués à la figure 2. - Pour analyser les données, exportez les données sous forme de rapport de feuille de calcul à partir du logiciel de contrôleur WAVE et utilisez un modèle de feuille de calcul indépendant pour l’analyse.
- Pour exporter des données dans l’un des générateurs d’analyse automatisés, suivez les commandes d’exportation de données sur le contrôleur WAVE et sélectionnez le générateur d’exportation qui correspond au type de test. Vous pouvez également exporter le fichier de données et le télécharger dans Seahorse Analytics.

Figure 2 : Descripteurs schématiques pour les paramètres dérivés des analyses de données de flux extracellulaire. Abréviation : OCR = taux de consommation d’oxygène. Veuillez cliquer ici pour voir une version agrandie de cette figure.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Pour obtenir des sphéroïdes compacts et bien formés, chaque lignée cellulaire a été optimisée individuellement pour la densité d’ensemencement et la durée de culture (figure 3). Les lignées cellulaires A549, HepG2/C3A et SK-OV-3 ont initialement formé des agrégats lâches qui n’ont progressé vers des sphéroïdes ronds avec des périmètres clairement définis qu’après 7 jours de culture. Inversement, les cellules MCF-7 pourraient former des sphéroïdes dans les 3 jours. Il y avait une corrélation claire entre la densité initiale d’ensemencement cellulaire et le volume de sphéroïdes après la période de culture pour tous les modèles de sphéroïdes. La taille et la morphologie des sphéroïdes ont été optimisées en fonction de la densité d’ensemencement. La morphologie et la circularité ont commencé à décliner avec l’augmentation de la taille des sphéroïdes dans tous les modèles. Les stratégies d’ensemencement des lignées cellulaires ont été optimisées à 4 × 103 cellules/puits pour les cellules A549 et SK-OV-3; Les cellules HepG2/C3A ont déjà été optimisées ailleurs à 1 × 103 cellules/puits, et les cellules MCF-7 ont été utilisées à 4 × 103 cellules/puits dans tous les essais. Lors de stratégies d’ensemencement optimisées, le volume de sphéroïdes se situait entre 5,46 ×10 7 μm3 (SK-OV-3) et 1,45 × 108 μm3 (A549) (Figure 3B). Tous les types de sphéroïdes avaient une corrélation linéaire entre la densité d’ensemencement initiale et le volume de sphéroïdes où A549 et HepG2/C3A avaient des valeursR2 de 0,957 et 0,947, respectivement. Les volumes de sphéroïdes MCF-7 et SK-OV-3 présentaient tous deux une plus grande corrélation avec la densité d’ensemencement initiale, R2 = 0,977 (figure 3A).
La circularité sphéroïde a été calculée à l’aide de la planimétrie d’image dans le logiciel d’analyse FIJI en utilisant les diamètres sphéroïdes longs et courts. La symétrie sphéroïde parfaite avait une circularité = 1,0; l’écart par rapport à 1,0 indiquait une perte de circularité (figure 3C). La circularité était plus grande dans les sphéroïdes MCF-7 que dans d’autres modèles où la circularité était maintenue entre 0,83 et 0,9 à toutes les densités d’ensemencement. En comparaison, le périmètre externe des sphéroïdes SK-OV-3 n’était pas aussi clairement défini, et le volume de sphéroïdes était significativement plus petit même après 7 jours de culture, produisant des sphéroïdes avec une circularité maximale de 0,61 à une densité d’ensemencement de 4 × 103/puits. Les cellules HepG2/C3A ont également formé des sphéroïdes serrés et bien formés avec une morphologie uniforme sur toute la surface de tous les sphéroïdes, avec une circularité maintenue à 0,79 pour les cellules ensemencées à 1 × 103 cellules / puits. Les cellules A549 semblaient suivre une tendance dans laquelle la circularité et la morphologie des sphéroïdes étaient améliorées avec la densité d’ensemencement; cependant, la circularité n’était pas supérieure à 0,63 à la densité utilisée dans ces expériences.
La respiration mitochondriale basale a été calculée comme OCR mesurée à partir de sphéroïdes ensemencés à 1 × 103, 2 × 103, 4 × 103, ou 8 × 103 cellules/puits dans des microplaques de culture sphéroïde à attachement ultra-faible (Figure 3D). Pour tous les types de sphéroïdes, l’OCR augmentait avec la taille des sphéroïdes et était linéairement corrélée au volume sphéroïde avec R2 le plus élevé dans les sphéroïdes MCF-7 à 0,988 et le plus bas dans les sphéroïdes SK-OV-3 à 0,744 (Figure 3E). L’OCR mesurée était statistiquement différente entre tous les groupes expérimentaux. L’A549 avait l’OCR la plus faible, atteignant seulement 18 pmol/min/puits à la plus grande taille de sphéroïde (Figure 3D). Inversement, les sphéroïdes MCF-7 ont produit un OCR similaire à la plus petite taille de sphéroïde après seulement 3 jours de culture, atteignant une OCR de base maximale de 53 pmol / min / puits pour la plus grande taille de sphéroïde (Figure 3D). HepG2/C3A a fourni des données OCR très cohérentes avec la taille et la morphologie des sphéroïdes. Dans les sphéroïdes HepG2/C3A ensemencés à partir de 1 × 103 cellules/puits, l’OCR de base a atteint une moyenne de 15 pmol/min/puits, augmentant jusqu’à un maximum de 52 pmol/min/puits dans les plus grands sphéroïdes (Figure 3D). L’OCR dans les sphéroïdes SK-OV-3 n’était significative qu’entre les sphéroïdes cultivés à partir de 4 × 103 cellules / puits et de 8 × 103 cellules / puits, avec peu de différence observée dans les sphéroïdes cultivés entre 1 × 103, 2 × 103, ou 4 × 103 cellules. Malgré les différences de taille, les données OCR étaient très similaires entre les sphéroïdes HepG2/C3A et MCF-7 à tous les points de taille. Par rapport à la taille des sphéroïdes (μm3), l’OCR de base par les sphéroïdes MCF-7 était comparable à celle des sphéroïdes HepG2/C3A cultivés sur 7 jours à partir de 1 000 cellules par puits.

Figure 3 : Les paramètres de croissance des sphéroïdes déterminent la respiration mitochondriale de base. (A) Les sphéroïdes A549, HepG2/C3A, MCF-7 et SK-OV-3 ont été optimisés pour la densité d’ensemencement et leur croissance surveillée à 1 ×10 3, 2 × 10 3, 4 × 103 et 8 × 103 cellules/puits dans chacune des photomicrographies, du haut à gauche vers le bas à droite, respectivement; barres d’échelle = 500 μm. (B) La taille des sphéroïdes a été calculée à l’aide de données planimétriques provenant de photomicrographies recueillies et comparée à l’aide de la statistique de corrélation de Pearson; les lignes pointillées représentent la distribution de l’intervalle de confiance à 95 %. (C) La morphologie des sphéroïdes a été comparée par calcul de circularité. (D) L’OCR a été mesurée 5x, après quoi la roténone-antimycine A a été ajoutée pour tenir compte de la fréquence respiratoire non mitochondriale à l’aide de l’analyseur Agilent Seahorse XFe96. OCRbasal mesuré comme OCR - OCRr / a a été comparé entre la densité d’ensemencement (D) et le volume sphéroïde (E). Les données sont des moyennes ± SEM de 5 à 8 répliqués de puits par type de sphéroïde et densité d’ensemencement cellulaire. Abréviations: OCR = taux de consommation d’oxygène; OCRbasal = Respiration mitochondriale basale; OCRr/a = OCR après ajout de roténone-antimycine A. Veuillez cliquer ici pour voir une version plus grande de cette figure.
La concentration et l’évolution temporelle de l’exposition aux modulateurs respiratoires dans l’analyse XF sont une étape critique dans l’optimisation des essais. Les composés modulateurs respiratoires - oligomycine, BAM15, un mélange de roténone-antimycine A - ou un véhicule de contrôle DMSO ont été injectés séquentiellement à travers les orifices d’injection de la cartouche du capteur dans des puits de microplaques contenant les sphéroïdes MCF-7 (Figure 4A). Quatre cycles de mesure ont été effectués pour déterminer l’OCR basale moyenne de 30 à 40 pmol/min/puits pour tous les groupes d’échantillons. Pour le reste de la durée de l’essai, des modulateurs respiratoires ont été ajoutés séquentiellement tous les 5 cycles de mesure pour atteindre des concentrations finales de puits de 0,5 μM (0,5 μg/mL d’oligomycine) après l’injection 1; 2,0 μM (2 μg/mL d’oligomycine) après injection 2; 5 μM (5 μg/mL d’oligomycine) après l’injection 3; et enfin, une concentration maximale dans le puits de 11 μM (11 μg/mL d’oligomycine) après la quatrième addition séquentielle. Les sphéroïdes MCF-7 n’ont pas répondu au contrôle du véhicule tout au long de l’expérience (Figure 4A). L’OCR basale a immédiatement changé après la première injection de chaque composé respectif à la concentration la plus faible de 0,5 μM ou 0,5 μg/mL d’oligomycine (figure 4B). L’OCR dans les sphéroïdes MCF-7 a été abaissée avec l’oligomycine de 41 pmol/min/puits à 23 pmol/min/puits après 5 cycles de mesure suivant la première injection de 0,5 μg/mL (Figure 4B).
En réponse à 0,5 μm de BAM15, l’OCR a été augmentée de 33 à 41 pmol/min/puits avant la deuxième injection (Figure 4C). Comparativement, la combinaison de roténone et d’antimycine A a abaissé l’OCR de 37 à 13 pmol/min/puits avant la deuxième injection (Figure 4D). Les traces cinétiques ont en outre révélé une diminution linéaire constante (oligomycine et roténone-antimycine A) ou une augmentation (BAM15) de l’OCR. Pour tous les schémas posologiques composés, une OCR à l’état d’équilibre a été obtenue en 10 à 12 cycles de mesure complets (60 à 72 min) à une concentration totale de puits de 2 μM BAM15, 2 μM de roténone, 2 μM d’antimycine A et 2 μg/mL d’oligomycine (figure 4A). Le taux de consommation d’oxygène a atteint un plateau à l’état d’équilibre à ~19 pmol/min/puits (oligomycine), 52 pmol/min/puits (BAM15) et 10 pmol/min/puits (roténone-antimycine A) (Figure 4A). L’augmentation de la concentration composée d’oligomycine, de BAM15 ou de roténone plus antimycine A n’a pas eu d’effet évident sur l’OCR, qui est resté constant pendant le reste du test. Ces données démontrent que la concentration en composés et l’évolution temporelle de l’exposition aux composés modulateurs respiratoires doivent être prises en compte pour l’optimisation des essais lors de l’utilisation de sphéroïdes 3D.

Figure 4 : Titrage des composés modulateurs respiratoires en tant qu’étape importante pour optimiser l’analyse du flux extracellulaire. (A) Les sphéroïdes MCF-7 ont été ensemencés à 4 × 103 cellules/puits et cultivés pendant 3 jours avant d’être placés dans des puits d’une microplaque de dosage sphéroïde contenant XF RPMI et sondés pour l’OCR ± modulateurs mitochondriaux à l’aide de l’analyseur XFe96. L’OCR a été mesurée 5x, après quoi des titrages de l’un ou l’autre véhicule témoin, de l’oligomycine (B), du BAM15 (C) ou de la roténone-antimycine A (D) ont été ajoutés pour inhiber l’ATP synthase mitochondriale, déterminer la capacité respiratoire maximale ou établir la fréquence respiratoire non mitochondriale, respectivement. La concentration de chaque modulateur mitochondrial a été augmentée au cours de 4 stratégies d’injection de titration individuelles (0,5 μM, 1,5 μM, 3 μM et 6 μM; les unités pour l’oligomycine sont μg / mL) pour déterminer l’OCR maximale à l’état d’équilibre en réponse à la concentration optimale de composés. L’OCR a été mesurée pendant 5 cycles de mesure entre chaque injection. Les données sont des moyennes ± SEM de 5 à 8 répliqués de puits individuels. Abréviation : OCR = taux de consommation d’oxygène. Veuillez cliquer ici pour voir une version agrandie de cette figure.
L’un des principaux avantages de la technologie XF est la capacité de sonder la fonction mitochondriale dans les cellules et les tissus intacts. Pour examiner des aspects spécifiques de la fonction mitochondriale dans les cellules et les tissus, des modulateurs mitochondriaux sont ajoutés séquentiellement aux puits de la microplaque de l’échantillon via les 4 ports d’injection disponibles sur la cartouche du capteur. La séquence typique de modulateurs utilisés pour sonder les paramètres mitochondriaux dans les tests XF est l’oligomycine, un protonophore (par exemple, FCCP ou BAM15) et une combinaison de roténone et d’antimycine A, qui sont ajoutés séquentiellement pour inhiber l’ATP synthase mitochondriale, déterminer la capacité respiratoire maximale et corriger la fréquence respiratoire non mitochondriale, respectivement. Cette séquence typique d’ajouts de modulateurs est appelée le test MitoStress par le fabricant de la technologie d’essai. Étant donné que l’oligomycine peut inhiber la respiration stimulée par un découpleur dans certaines monocouches cellulaires20, nous l’avons examiné avec des sphéroïdes 3D dérivés du cancer en mesurant l’OCR non couplée -stimulée (OCRmax) avant (simple) et après (séquentielle) injection d’oligomycine (Figure 5A-D). L’OCRmax n’a pas été significativement limitée par l’ajout d’oligomycine dans les sphéroïdes formés à partir de HEPG2/C3A ou SK-OV-3 (Figure 5E et Figure 5G). Cependant, l’OCRmax a été significativement abaissé dans les sphéroïdes A549 et MCF-7 après une injection séquentielle de BAM15 après oligomycine par rapport à l’OCRmax obtenue à partir d’une seule injection de BAM15 (Figure 5F et Figure 5H). Sauf indication contraire, il est donc recommandé d’utiliser des puits séparés pour traiter avec de l’oligomycine et un découpleur, avec un ajout final de roténone et d’antimycine A lors de l’exploration du métabolisme énergétique mitochondrial des sphéroïdes 3D. Cette approche permet toujours le calcul de tous les paramètres mitochondriaux comme avec un test MitoStress typique où les composés sont ajoutés séquentiellement.

Figure 5 : Injection simple ou séquentielle de composés respiratoires mitochondriaux. Des sphéroïdes dérivés de cellules cancéreuses de MCF-7, HEPG2/C3A, SK-OV-3 et A549 ont été placés dans des puits d’une microplaque sphéroïde XFe96 dans XF RPMI et sondés pour OCR à l’aide de l’analyseur Agilent Seahorse XFe96. L’OCR a été mesurée 5x, après quoi 2 μg/mL d’oligomycine (port d’injection A : trace verte) ou 5 μM de BAM15 (port d’injection A : trace bleue ou port d’injection B : trace verte) pour inhiber l’ATP synthase mitochondriale et déterminer la capacité respiratoire maximale, respectivement. Les données OCR cinétiques sont exprimées en % basal (A-D). La capacité respiratoire maximale (OCRmax) a été calculée comme un facteur d’OCR basal par l’équation : OCRmax = OCRBAM15 / OCRbasal. L’OCRmax a été obtenue à partir des moyennes DE l’OCR sur les cycles de mesure 8-10 après l’injection de BAM15 avec (barres vertes) et sans (barres bleues) oligomycine. Les données sont des moyennes ± SEM de 3 à 8 répliqués de puits individuels à travers la microplaque de test sphéroïde. Abréviations : OCR = taux de consommation d’oxygène. Veuillez cliquer ici pour voir une version agrandie de cette figure.
En utilisant les densités d’ensemencement cellulaire optimales, les concentrations de composés, la stratégie d’injection et la période de cycle de mesure déterminées dans ces expériences d’optimisation (Tableau 3), nous avons développé un protocole détaillé pour sonder avec précision la respiration mitochondriale basale : OCRbasal (Figure 6A), la respiration de phosphorylation ADP : OCRADP (Figure 6B), la respiration des fuites : OCRomy (Figure 6C), l’efficacité du couplage (Figure 6D ), capacité respiratoire maximale : OCRmax (Figure 6E) et capacité respiratoire de réserve : OCRde rechange (Figure 6F) à l’aide de sphéroïdes 3D dérivés du cancer.

Figure 6 : Sondage de l’OCR avec la technologie XF pour établir le métabolisme énergétique mitochondrial des sphéroïdes dérivés du cancer. Les sphéroïdes dérivés de cellules cancéreuses de MCF-7, HEPG2/C3A, SK-OV-3 et A549 ont été placés dans des puits d’une microplaque de dosage sphéroïde dans XF RPMI et sondés pour OCR à l’aide de l’analyseur Agilent Seahorse XFe96. L’OCR a été mesurée 5x, après quoi 2 μg / mL d’oligomycine, ou 5 μM BAM15, et la PR a été ajoutée pour inhiber l’ATP synthase mitochondriale, déterminer la capacité respiratoire maximale et calculer la fréquence respiratoire non mitochondriale, respectivement. (A) La respiration mitochondriale basale (OCRbasale) a été calculée comme la moyenne de l’OCR à partir des 3 cycles de mesure avant l’injection du port A. (B) L’efficacité de couplage de la phosphorylation oxydative a été approchée en exprimantl’OCR ADP (OCR basal-OCR leak) en pourcentage de l’OCRbasal. (C) La respiration par phosphorylation de l’ADP (OCRADP) a été mesurée en OCR sensible à l’oligomycine, calculée à partir de l’OCR moyenne sur les cycles de mesure 11 à 13 précédant l’injection de BAM15. (D) La respirationpar fuite (fuite OCR) a été mesurée comme insensible à l’OCR à l’oligomycine, calculée à partir de la moyenne moyenne de l’OCR sur les cycles de mesure 11 à 13. (E) La capacité respiratoire maximale (OCRmax) a été mesurée comme l’OCR maximale moyenne mesurée après l’injection de BAM15. (F) La capacité respiratoire disponible a été calculée en exprimant laréserve OCR (OCRmax - OCRbasal) en pourcentage de l’OCRbasal. L’OCR après injection de roténone-antimycine A (OCRr/a) a été soustraite de tous les taux pour corriger l’OCR nonmitochondriale. Les données sont des moyennes ± SEM de 3 à 8 répliqués de puits individuels sur la plaque sphéroïde XFe96. Abréviations: OCR = taux de consommation d’oxygène; RA = 2 μM roténone-2 μM antimycine A. Veuillez cliquer ici pour voir une version plus grande de cette figure.
Les sphéroïdes MCF-7 cultivés à partir de 4 × 103 cellules / puits sur 3 jours ont été utilisés comme modèle pour déterminer le transfert, le placement et l’analyse optimaux dans les microplaques de dosage des sphéroïdes. En utilisant les dimensions fournies pour la microplaque sphéroïde par le fabricant, la surface du puits a été divisée en trois zones pour un placement optimal des sphéroïdes (Figure 7A), où la zone 1 a été mise en évidence comme la zone optimale au centre du puits. Avec un pipetage minutieux à l’aide d’embouts de pipette à orifice large, les sphéroïdes ont été transférés dans les plaques sphéroïdes et répartis de manière aléatoire sur les surfaces du puits par élution par gravité (figure 7B). Lorsque les sphéroïdes étaient soigneusement transférés à l’aide de l’élution par gravité, la plupart des sphéroïdes pouvaient généralement être trouvés dans les zones 1 à 2 de la microplaque, en utilisant les techniques de transfert recommandées par le fabricant. Lorsque les sphéroïdes étaient forcés de sortir de la pointe de la pipette par aspiration, les sphéroïdes étaient souvent placés au-delà de ces zones et ne pouvaient pas être vus à l’aide de la microscopie.
Pour comparer les positions de placement des sphéroïdes, les sphéroïdes MCF-7 ont été transférés dans les microplaques d’essai des sphéroïdes dans les zones désignées 1 à 3 ou hors zone (figure 7A). Ces 4 puits ont été suivis par une expérience cinétique OCR au départ et après l’ajout d’oligomycine, de BAM15 ou de roténone-antimycine A (Figure 7C). L’OCR a été calculée à partir de la moyenne de trois cycles avant chaque injection (figure 7B). L’OCR a été mesurée cinétiquement pendant 200 minutes dans les 4 puits sélectionnés (figure 7C) et corrigée de la ligne de base (figure 7D). Lorsque les sphéroïdes étaient placés dans la zone 3 ou hors zone, les OCR de base étaient significativement plus faibles que les sphéroïdes placés dans les zones 1 et 2 (figure 7C). Les effets des composés respiratoires oligomycine, BAM15 et roténone-antimycine A différaient également considérablement entre les sphéroïdes placés dans les zones 1 et 2 par rapport aux régions de zone 3 et hors zone. Une augmentation de l’OCR a été observée avec l’oligomycine chez les sphéroïdes placés dans la zone 3 ou hors zone (figure 7E). De plus, les sphéroïdes placés dans la zone 3 ou hors zone ont présenté une réponse excessivement élevée au BAM15 avec une OCR supérieure à la ligne de base après l’injection de roténone-antimycine A (figure 7E). Malgré une augmentation presque deux fois de l’OCR basale (Figure 7C) avec des sphéroïdes placés dans la zone 2 par rapport à la zone 1, les changements de plis en réponse à tous les composés respiratoires étaient très similaires (Figure 7E), ce qui suggère que les différences d’OCR basal entre les sphéroïdes placés dans les zones 1 ou 2 sont peu susceptibles d’être le résultat d’un placement dans le puits.

Figure 7 : Le placement des sphéroïdes dans la microplaque du test sphéroïde dicte les effets de l’OCR basal et du modulateur mitochondrial à l’aide de la technologie XF. Les sphéroïdes MCF-7 ont été ensemencés à 4 × 103 cellules / puits et cultivés pendant 3 jours avant d’être placés dans les puits de la microplaque sphéroïde contenant XF RPMI et sondés pour l’OCR ± modulateurs mitochondriaux à l’aide de l’analyseur Agilent Seahorse XFe96. A) Photomicrographies des positions de la zone sphéroïde dans les microplaques d’essai sphéroïdes après la durée de l’essai; barre d’échelle = 500 μm et OCR capturés à partir des puits correspondants au fil du temps, exprimés en pmol/min-1/puits-1 (B) ou % basal (C). D) Effets modulateurs mitochondriaux des sphéroïdes MCF-7 placés dans différentes zones de la microplaque d’essai des sphéroïdes; données exprimées sous forme de changement de pli par rapport à la base. (E) Exemple de trace cinétique mettant en évidence quelles mesures de données OCR (cercles rouges) sont utilisées pour calculer la réponse de chaque modulateur mitochondrial pour les données présentées dans E. Les données présentées proviennent des réponses individuelles aux puits. Abréviation : OCR = taux de consommation d’oxygène. Veuillez cliquer ici pour voir une version agrandie de cette figure.
Les critères de sélection de l’arrière-plan sont d’une grande importance; l’utilisation de puits les plus extérieurs pour la correction de fond n’est pas représentative de tous les puits de microplaques, ce qui peut conduire à des hypothèses de données incorrectes et à des conclusions erronées en raison d’effets de bord sur la microplaque sphéroïde. Pour évaluer cette observation, des sphéroïdes MCF-7 ont été utilisés pour comparer les procédures de correction du test afin d’obtenir des valeurs OCR en réponse à l’ajout d’un témoin de véhicule, l’oligomycine, le BAM15 ou la roténone-antimycine A (figure 8). Tous les composés respiratoires ont produit les profils OCR cinétiques attendus pour les composés sélectionnés, révélant un taux de respiration basale stable moyen de 20 à 30 pmol/min/puits (figure 8A). Toutefois, lorsque les données d’essai ont été analysées à l’aide des puits les plus extérieurs pour la correction de la température de fond (A1, A12, H1 et H12), les valeurs révélées pour l’OCR après l’ajout de composés respiratoires étaient particulièrement faibles; L’OCR a donné des valeurs négatives pour la roténone-antimycine A. En réponse à ces observations, une autre analyse a été effectuée à l’aide d’une série de puits vides, répartis au hasard sur la microplaque sphéroïde, comme puits de correction de la température de fond (figure 8B). Lorsque d’autres corrections de fond ont été appliquées, tous les effets composés relatifs sur l’OCR étaient les mêmes dans les deux ensembles d’analyse; toutefois, les valeurs absolues d’OCR ont augmenté d’environ 10 pmol/min/puits (figure 8). Ces données mettent en évidence la puissance et l’importance de la correction de la température de fond sur les microplaques de dosage des sphéroïdes et soulignent l’importance de l’optimisation de l’utilisateur pour l’analyse XF.

Figure 8 : Sélection aléatoire des puits pour la correction de fond afin d’améliorer le contrôle des gradients de température à travers la microplaque d’essai sphéroïde. Données OCR extrapolées à partir de la figure 2A à l’aide de puits recommandés pour la correction de fond (A) par rapport à des puits assignés au hasard pour la correction de fond (B). Abréviation : OCR = taux de consommation d’oxygène. Veuillez cliquer ici pour voir une version agrandie de cette figure.
Contrairement aux monocouches cellulaires, les sphéroïdes représentent une agrégation hétérogène de cellules dans un espace 3D et nécessitent donc une réflexion approfondie en ce qui concerne l’analyse, en particulier lors de la normalisation de ces données. Cet article présente trois approches pour normaliser les données XF acquises à partir des sphéroïdes MCF-7 (Figure 9). Lorsqu’elle n’est pas normalisée, l’OCR est positivement corrélée (R2 = 0,98) avec la taille des sphéroïdes (déterminée par la densité initiale d’ensemencement des cellules) de manière significative par rapport au coefficient de corrélation de Pearson, P = 0,0057 (figure 9A). Cette relation linéaire est réduite lorsque l’OCR est normalisée à la densité initiale d’ensemencement cellulaire (R2 = 0,78) et n’est plus significativement corrélée à la taille des sphéroïdes (P = 0,117, figure 9B). C’est également le cas lorsqu’il est normalisé au volume sphéroïde (R2 = 0,77; Coefficient de corrélation de Pearson P = 0,120, figure 9C) et teneur en ADNds nucléaire (R2 = 0,58; Coefficient de corrélation de Pearson P= 0,233, figure 9D). Ces données soulignent l’importance de normaliser les données XF lors de l’étude du métabolisme mitochondrial des sphéroïdes, surtout s’ils sont de tailles différentes.

Figure 9 : Normalisation des données de flux extracellulaire acquises à partir de sphéroïdes cellulaires. (A) Les données OCR brutes ont été obtenues à partir de MCF-7 cultivés sur 3 jours et tracées à l’aide du modèle de Pearson pour obtenir un coefficient de corrélation entre la densité d’ensemencement des sphéroïdes et l’OCR; Valeur P fixée à 0,05. B) Les données OCR brutes ont été normalisées par rapport à la densité initiale d’ensemencement des sphéroïdes; C) volume de sphéroïde MCF-7 obtenu par planimétrie microscopique; et (D) la teneur en ADN nucléaire ds comparée à l’aide du coefficient de corrélation de Pearson. Abréviations: OCR = taux de consommation d’oxygène; ADN ds = ADN double brin. Veuillez cliquer ici pour voir une version agrandie de cette figure.
| Lignée cellulaire | Densité d’ensemencement (puits) | Croissance sphéroïde (jours) | Volume final des sphéroïdes (μM3) | OCR basal (pmolO2/min/puits) | Sensibilité à l’OCR basale satisfaite (OUI/NON) |
| SKOV | 1000 | 5 | 9.52E+06 | 28 ± 3,5 | Oui |
| SKOV | 2000 | 5 | 2.38E+07 | 26 ± 1,4 | Oui |
| SKOV | 4000 | 5 | 4,92E+07 | 36 ± 3.1 | Oui |
| SKOV | 8000 | 5 | 1.11E+08 | 50 ± 7,9 | Oui |
| HepG2 | 1000 | 5 | 1.11E+07 | 15 ± 0,7 | Non |
| HepG2 | 2000 | 5 | 2,88E+07 | 23 ± 1,8 | Oui |
| HepG2 | 4000 | 5 | 5,46E+07 | 31 ± 1,7 | Oui |
| HepG2 | 8000 | 5 | 1.21F+08 | 52 ± 2,8 | Oui |
| L’A549 | 1000 | 5 | 2.11E+07 | 30 ± 2,5 | Oui |
| L’A549 | 2000 | 5 | 3,57E+07 | 41 ± 1,6 | Oui |
| L’A549 | 4000 | 5 | 6,93E+07 | 53 ± 7,2 | Oui |
| L’A549 | 8000 | 5 | 1,44E+08 | 65 ± 8,4 | Oui |
| MCF-7 | 1000 | 3 | 1,60E+07 | 29 ± 0,8 | Oui |
| MCF-7 | 2000 | 3 | 2.52E+07 | 37 ± 1,7 | Oui |
| MCF-7 | 4000 | 3 | 6.00E+07 | 46 ± 1,7 | Oui |
| MCF-7 | 8000 | 3 | 1.06E+08 | 66 ± 2,9 | Oui |
Tableau 4 : Paramètres optimisés pour la détermination des mesures OCR basales dans des sphéroïdes 3D uniques. Abréviation : OCR = taux de consommation d’oxygène.
Fichier supplémentaire 1 : Analyse de la taille et du volume des sphéroïdes. Veuillez cliquer ici pour télécharger ce fichier.
Fichier supplémentaire 2 : Quantification de l’ADN double brin à partir de sphéroïdes dans la microplaque sphéroïde. Veuillez cliquer ici pour télécharger ce fichier.
Fichier supplémentaire 3 : Recommandations concernant le nombre de réplications requises pour obtenir des ensembles de données de dosage XF fiables. Veuillez cliquer ici pour télécharger ce fichier.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Principales constatations et extrants
Cet article fournit un protocole détaillé pour sonder le métabolisme énergétique mitochondrial de sphéroïdes 3D uniques à l’aide d’une série de lignées cellulaires dérivées du cancer avec l’analyseur XFe96 XF. Une méthode est développée et décrite pour la culture rapide des sphéroïdes cellulaires A549, HepG2/C3A, MCF7 et SK-OV-3 en utilisant des technologies d’hydrofuge cellulaire pour l’agrégation forcée. Ce protocole aborde de nombreuses considérations relatives à la sonde du métabolisme des sphéroïdes avec la technologie XF, y compris (1) l’optimisation des protocoles de culture des sphéroïdes et la manipulation et le transfert des sphéroïdes dans des microplaques de dosage sphéroïdes spécifiques du fabricant de la technologie à partir de leurs récipients de culture d’origine; 2° la concentration des composés respiratoires à utiliser et la dépendance temporelle de la pénétration des composés; 3° les stratégies d’injection à utiliser; et (4) les moyens de normaliser les données entre les groupes expérimentaux. Toutes ces considérations ont été examinées dans le présent document et sont examinées plus en détail ci-dessous. Ces méthodes sont présentées comme des approches simplifiées pour générer des données de flux d’oxygène métabolique cohérentes à l’aide de sphéroïdes 3D uniques avec l’analyseur de flux XFe96. Cette approche expérimentale peut être utilisée comme point de départ et rubrique pour une utilisation dans d’autres modèles sphéroïdes qui sont facilement mis en œuvre dans un environnement de laboratoire de base.
Considérations
Croissance, taille et sensibilité des sphéroïdes de la technologie XF
Pour établir des données reproductibles avec la technologie XF, il est essentiel de caractériser et d’optimiser le test pour le modèle spécifique. Cette approche est relativement simple dans une monocouche de base de cellules; cependant, cela présente des défis supplémentaires lors de la culture de cellules en tant que sphéroïdes 3D. Au cours des expériences présentées ici, le milieu RPMI du fabricant a été complété lors de l’utilisation. Bien qu’il soit noté que certaines lignées cellulaires, à savoir HepG2/ C3A, ont été cultivées dans un milieu de croissance DMEM, au cours de ces essais relativement courts (~ 3-5 h), la substitution par des formulations RPMI-DMEM a eu un impact limité sur l’analyse XF. La formulation des deux milieux est très similaire, et les utilisateurs pourraient « ajuster » les milieux RPMI seahorse pour qu’ils correspondent à la matrice de leurs milieux de culture cellulaire grâce à une supplémentation, par exemple une augmentation du glucose, un ajout supplémentaire de sources de glucides. L’absence de phénol-rouge, qui est susceptible d’interférer avec les sondes fluorescentes dans la plaque de la cartouche de sonde XF, et de bicarbonate de sodium, qui conduira à l’alcalinité en raison de l’absence de tampon CO2 présent dans les incubateurs de culture cellulaire, est essentielle à la formulation finale de tous les tampons XF. D’autres supports et tampons peuvent être achetés et/ou fabriqués en interne. Par exemple, le tampon HEPES de Krebs Ringer est un tampon simple qui peut être utilisé pour évaluer la respiration dans de nombreuses cellules différentes, y compris les modèles sphéroïdes. Cependant, les utilisateurs de tests XF doivent noter qu’un changement de milieu/tampon et sa supplémentation peuvent modifier sa capacité tampon globale. Ceci est particulièrement préoccupant lorsque les utilisateurs peuvent être intéressés par la mesure de l’ECAR, dans lequel le facteur tampon du milieu doit être évalué pour permettre la transformation de l’ECAR en taux d’efflux de protons (PER).
Comme l’OCR cellulaire mesurée par la technologie XF est proportionnelle à la densité cellulaire lorsque le nombre de cellules dans le puits est dans la sensibilité du système, il était important d’étudier cette relation à l’aide de sphéroïdes 3D uniques. En sondant l’OCR de sphéroïdes 3D uniques cultivés à partir de 4 lignées cellulaires cancéreuses différentes ensemencées à des densités de 1 000, 2 000, 4 000 ou 8 000 cellules par puits, nous montrons que l’analyseur XFe96 est suffisamment sensible pour détecter les changements dans le taux de respiration mitochondriale entre les sphéroïdes 3D cultivés à partir de différentes densités d’ensemencement cellulaire (Figure 3). Nous montrons que la plage optimale de densité d’ensemencement cellulaire, et donc le volume sphéroïde pour former des sphéroïdes 3D pour sonder l’OCR, diffèrent selon le type de cellule. Ceci est illustré par la relation linéaire entre l’OCR et la densité d’ensemencement ou le volume sphéroïde (Figure 3). Pour les cellules A549 et HepG2/C3A, la densité d’ensemencement optimale pour la sensibilité à l’OCR se situait entre 1 000 et 8 000 cellules/puits; il était de 2 000 à 8 000 cellules/puits pour le MCF-7 et de 4 000 à 8 000 cellules/puits pour les cellules SK-OV-3. Ces données démontrent que l’optimisation de la taille des sphéroïdes est particulièrement importante lors de l’évaluation de l’OCR à l’aide de la technologie XF.
Considérations sur les volumes sphéroïdes minimaux et maximaux et l’OCR basal
En général, il y aura toujours des seuils minimaux et maximaux pour les paramètres OCR mesurables recommandés par le fabricant pour ces expériences. Pour l’analyseur XFe96, l’OCR basal entre 20 pmol O2/min/puits et 200 pmol O2/min/puits sont les limites inférieure et supérieure, respectivement. C’est le cas des cellules monocouches et des sphéroïdes, et l’emplacement du modèle expérimental dans cette plage d’OCR dynamique dépendra de la quantité de matériel biologique disponible, par exemple, du nombre de cellules en monocouche ou de la taille des sphéroïdes. Voir le tableau 4 pour un exemple de la façon dont les seuils d’OCR ont été atteints par les modèles sphéroïdes utilisés ici. Il peut être prudent de vérifier le niveau d’oxygène dans le puits pour lequel ces données sont également disponibles à partir de ces mesures en tant que données de niveau. Cela devrait être vu régulièrement à partir de chaque expérience à des fins de contrôle de la qualité. S’il y a un épuisement de l’oxygène dans le puits, cela sera mis en évidence dans les données. Si tel est le cas, il peut être nécessaire d’ajuster les cycles de mesure au sein de l’expérience; par exemple, augmenter l’étape de mélange de manière à ce que le niveau d’oxygène dans le puits soit récupéré avant la prochaine période de mesure du cycle de mesure. Bien que possible, nous avons constaté que cela était très peu probable pour les expériences à sphéroïde unique utilisant les lignées cellulaires décrites.
Choix de découpleurs mitochondriaux pour les tests de flux extracellulaire
Les ionophores de protons, tels que le cyanure de carbonyle 4-(trifluorométhoxy) phénylhydrazone (FCCP)21, le cyanure de carbonyle m-chlorophénylhydrazone (CCCP)22 ou BAM1523, sont de puissants produits chimiques à petites molécules capables de perturber le gradient électrochimique des protons à travers les membranes mitochondriales, d’inhiber la production d’ATP et, en fin de compte, de découpler la respiration mitochondriale24 . De nouvelles petites molécules continuent d’être développées à ces fins, en particulier dans le traitement des maladiesmétaboliques 25,26,27; se référer à deux excellentes critiques 28,29. Inversement, le découplage de la respiration oxydative a été associé à une toxicité indésirable hors cible30. Cependant, dans les essais cellulaires in vitro, la molécule FCCP dépolarise le potentiel de la membrane mitochondriale et exerce des effets hors cible tels que la dépolarisation de la membrane plasmique, perturbant le flux d’ions NA+ 31; interférence avec le traitement des protéines cellulaires32, et même induisant la sénescence cellulaire33. BAM15 a été introduit à l’origine en 2013 en tant que découpleur mitochondrial avec une influence minimale sur les membranes plasmiques23, avec une activité protonophorique de l’ordre micromolaire dans les cellules entières et de la gamme nanomolaire dans les mitochondries isolées 23,34.
Compte tenu de la puissance du FCCP sur la dépolarisation de la membrane plasmique, BAM15 est un protonophore plus fiable pour découpler la respiration dans des cellules entières intactes dans des tests de flux extracellulaires. Bien que le FCCP et son homologue, le CCCP, soient utilisés depuis plus de 50 ans pour tester les capacités respiratoires maximales et continuent d’être largement utilisés dans les études XF, l’utilisation de ces petites molécules sous-estime souvent la capacité métabolique mitochondriale et cellulaire. Cela est en partie lié à la raison pour laquelle tant de publications utilisant la technologie XF tombent dans le piège de signaler des capacités respiratoires inutilisées négatives ou de sous-estimer les capacités respiratoires mitochondriales réelles lorsque le FCCP est utilisé. La puissance supplémentaire du FCCP dans les cellules et les tissus intacts conduit souvent à une fonction mitochondriale compromise, et les cellules peuvent avoir du mal à fonctionner de manière appropriée pour maintenir une capacité respiratoire maximale sur plusieurs cycles de mesure après leur ajout, même à de très faibles concentrations35. Par conséquent, la réponse des cellules au FCCP peut être trouvée dans de nombreuses études pour diminuer après la période du cycle de mesure initial. Alors que le FCCP a été couramment utilisé pour l’analyse XF, BAM15 est utilisé de préférence dans les cas impliquant des cellules entières ou des modèles sphéroïdes, étant donné qu’il peut maintenir une capacité respiratoire maximale dans les mitochondries entièrement dépolarisées à des concentrations aussi élevées que 10 μM 3. De plus, BAM15 induit des effets sur l’acidification extracellulaire, qui coïncide avec celle de l’oxydation des nutriments par l’hydratation du CO2 pour former HCO3- et H+ dans une plus grande mesure que le FCCP3. Néanmoins, dans le cas de mitochondries isolées et de cellules perméabilisées, l’un de ces découpleurs devrait fonctionner aussi bien que BAM15 pour le découplage mitochondrial s’il est titré à la concentration correcte.
Cinétique de la pénétration des composés et du cycle d’essai
Les concentrations, la pénétration et les profils cinétiques des composés chimiques utilisés pour effectuer un test MitoStress typique avec des sphéroïdes 3D à l’aide de l’analyseur XF sont plus complexes à traiter. Étant donné que les sphéroïdes présentent une structure 3D, la pénétration des molécules sur le diamètre du sphéroïde est un processus infiniment plus complexe que sur les monocouches cellulaires. Par exemple, la pénétration cinétique et, par conséquent, la sensibilité au sorafénib chimiothérapeutique ont été déterminées par l’âge sphéroïde et, par conséquent, la taille dans un modèle de sphéroïde HepG236. La capacité des produits chimiques à petites molécules (p. ex., médicaments, nanoparticules) à atteindre une cible biologique dépend de plusieurs facteurs sous-jacents, y compris la complexité du système à pénétrer et à diffuser dynamiquement à travers37,38. Cela est particulièrement vrai pour les médicaments ciblant le tissutumoral 39. Semblable au ciblage tumoral dans le contexte d’un sphéroïde 3D, la taille, la compacité et d’autres réponses phénotypiques telles que l’expression de protéines transporteuses de médicaments peuvent régir le temps de pénétration et la concentration d’un composé nécessaire pour provoquer une réponse biologique.
Dans ce protocole, nous avons abordé la question du temps de pénétration et de la concentration de petites molécules en réponse à l’inhibiteur de l’ATP synthase oligomycine, au protonophore et au découpleur mitochondrial BAM15, et à la combinaison des inhibiteurs du complexe I et du complexe III roténone et antimycine A. En sondant l’OCR de sphéroïdes MCF-7 uniques exposés à de multiples titrages de ces composés respiratoires courants, nous démontrons que la concentration optimale de chaque composé nécessaire pour induire une fréquence respiratoire à l’état d’équilibre se situe dans une plage similaire à celle des cellules monocouches (Figure 4). Fait important, et contrairement à leurs homologues monocouches, il est démontré que l’augmentation du nombre de cycles de mesure entre les injections est essentielle pour obtenir un OCR à l’état d’équilibre dans les sphéroïdes 3D uniques. Ces données soulignent l’importance de la pénétration des composés et de leurs profils cinétiques respectifs lors de l’exploration des paramètres respiratoires mitochondriaux des sphéroïdes 3D à l’aide de ces approches. En utilisant les propriétés d’optimisation des sphéroïdes, les concentrations de composés et les temps de cycle de mesure éclairés par les données présentées à la figure 3, à la figure 4 et à la figure 5, un test MitoStress validé a été établi pour sonder des paramètres spécifiques du métabolisme oxydatif mitochondrial dans une gamme de sphéroïdes 3D dérivés du cancer (figure 6). Fait important, et comme certaines lignées cellulaires cancéreuses monocouches40, la capacité respiratoire maximale (taux de respiration non couplée-stimulée) de certains sphéroïdes 3D dérivés du cancer a été inhibée par l’oligomycine (figure 5). Plus précisément, les sphéroïdes 3D cultivés à partir de cellules A549 ou MCF-7 ont montré un taux de respiration maximal significativement plus faible lorsqu’ils étaient découplés avec BAM15 après injection d’oligomycine par rapport à ceux qui étaient découplés par BAM15 sans oligomycine (Figure 5F et Figure 5H). Étant donné que cet effet peut être présent dans d’autres cultures de sphéroïdes 3D, nous suggérons qu’à moins qu’un protocole préalablement validé ne soit utilisé, la capacité respiratoire maximale dans les sphéroïdes 3D devrait être estimée sans oligomycine.
Collecte simultanée de données ECAR comme mesure du flux glycolytique dans les sphéroïdes cellulaires
Comme on le voit généralement dans la littérature ou les informations du fabricant de la technologie, le taux glycolytique des sphéroïdes, mesuré en ECAR, est un paramètre secondaire qui peut être capturé avec l’OCR. Le calcul de l’ECAR seul n’est pas un paramètre utile ou significatif dans toute expérience XF car il n’est pas corrigé pour la capacité tampon du tampon de dosage XF ou l’ajout d’acidification mitochondriale, qui résulte de l’hydratation du CO2 en HCO3- et H +. L’ECAR n’est perspicace qu’une fois ces corrections de données appliquées, après quoi il devient possible de fournir des conclusions plus précises sur le flux glycolytique. Pour corriger la capacité de mise en mémoire tampon afin de générer des données PER plus significatives, il faut connaître le volume de la microchambre pour la microplaque sphéroïde. Le fabricant n’a pas été en mesure de fournir un volume réel pour cela avec la microplaque sphéroïde, et par conséquent, les données PER ne peuvent pas être déterminées facilement. En effet, bien que ces mesures aient pu être réalisées empiriquement, cela dépassait le cadre de ce manuscrit. Cependant, avec les corrections appropriées et la connaissance du volume de la microchambre pour une taille de sphéroïde donnée présente (par exemple, l’obtention d’une mesure de la densité sphéroïde) dans le puits, les données ECAR deviendraient significatives et des calculs de PER glycolytique pourraient être effectués. Par conséquent, les données XF pourraient alors être plus informatives pour étudier le métabolisme glycolytique et oxydatif dans les sphéroïdes, mais seulement si ces paramètres étaient examinés en profondeur.
Formation, manipulation, transfert et mouvement de sphéroïdes
Certaines lignées cellulaires sont mieux adaptées à la formation de sphéroïdes que d’autres et peuvent ne pas former de sphéroïdes du tout, par exemple, les cellules cancéreuses de l’ovaire MCF-741,42 forment des sphéroïdes hautement circulaires par rapport à d’autres lignées cellulaires (Figure 3). Comme autre exemple, il a été démontré que les cellules cancéreuses du pancréas Capan-1 forment de meilleurs sphéroïdes que Panc-1 ou BxPC343. De même, les lignées cellulaires de carcinome hépatique sont connues pour avoir des capacités variables à formerdes sphéroïdes compacts 5,44, avec un changement observé dans le phénotype tel qu’un métabolisme accru des médicaments ou la production d’albumine, comme c’est le cas pour HepG2 versus HepG2/C3A 9,45,46 ou HepaRG sphéroïdes 17,47,48 . Par conséquent, les utilisateurs doivent optimiser les techniques de culture des sphéroïdes en conséquence et effectuer des expériences de titrage pour déterminer la densité d’ensemencement optimale et le déroulement de la culture. En outre, il a été démontré que la formulation et la composition des milieux d’essai ont un impact sur la formulation des sphéroïdes, y compris l’ajout de méthylcellulose, souvent ajoutée aux milieux pour augmenter la viscosité de la matrice 43,49,50. Par conséquent, la composition optimale du milieu cellulaire doit être déterminée empiriquement pour toutes les lignées cellulaires utilisées.
Le nombre d’échanges de milieu tout au long de la culture sphéroïde est déterminé par la lignée cellulaire utilisée. Cependant, généralement, des échanges de milieu à demi-volume tous les 2-3 jours sont applicables dans la plupart des cas pour reconstituer les nutriments. Nous avons utilisé l’approche de l’agrégat forcé pour générer des sphéroïdes 3D à l’aide de microplaques répulsives à partir de sources disponibles dans le commerce pour le développement et le déploiement rapides de modèles de sphéroïdes dans les études d’analyses XF. Cependant, d’autres plates-formes peuvent être mieux adaptées pour générer des sphéroïdes à partir d’autres types de cellules, par exemple, des approches suspendues-tombantes ou intégrées à la matrice. Dans les laboratoires aux ressources limitées, les utilisateurs peuvent envisager la technique de superposition agarose-liquide pour la formation de surfaces de microplaques répulsivescellulaires 51,52 afin de réduire considérablement les coûts économiques des étapes initiales de développement de la méthode sphéroïde. Le mouvement des sphéroïdes entre les récipients de culture est nécessaire pour effectuer une analyse XF et d’autres essais en aval. La facilité de transfert est généralement dictée par la taille des sphéroïdes et la densité globale. Nous vous recommandons d’utiliser une pointe de pipette à orifice large P200 ou P1000 pour maintenir l’intégrité des sphéroïdes; les embouts de pipette de plus petit calibre risquent de perturber mécaniquement le sphéroïde, qui peut être acheté dans le commerce ou, avec soin, fabriqué en coupant simplement l’extrémité de l’extrémité de la pipette pour augmenter l’orifice. Cependant, cette approche peut être susceptible d’introduire de la fourrure dans le plastique autour de l’extrémité de la pointe, ce qui pourrait provoquer des perturbations mécaniques lors de la manipulation. L’utilisation d’un rétroéclairage ou d’une boîte à lumière est également utile pour la manipulation et l’observation des sphéroïdes sous un microscope à dissection comme étape essentielle pour assurer le transfert réussi des sphéroïdes dans la microplaque de dosage des sphéroïdes. De plus, la position du sphéroïde dans le puits d’une microplaque d’essai sphéroïde est particulièrement importante et a un impact direct sur l’OCR et l’effet composé lors d’un test MitoStress typique (Figure 7), probablement en raison de la relation entre la position du sphéroïde et les fluorophores de la sonde du capteur.
Puits de correction de fond et de contrôle de la température
L’utilisation de tests à base de microplaques est une approche largement utilisée dans plusieurs domaines de recherche; cependant, leur utilisation présente plusieurs défis pratiques. Comme c’est le cas dans d’autres approches expérimentales, en particulier celles qui utilisent le format de matrice 96 (ou plus), la géométrie et le positionnement des microplaques peuvent influencer les gradients de température et d’échange de gaz à travers la plaque au fil du temps, souvent appelés « effets de bord»53,54. Nous avons constaté qu’il en allait de même pour la microplaque de dosage des sphéroïdes. Selon les directives et les protocoles du fabricant, les puits d’angle les plus extérieurs: A1, A12, H1 et H12 sont toujours désignés comme puits de correction de fond et de contrôle de la température pour l’analyseur XFe96. Inversement, avec le format matriciel de 24 puits, A1 et D6 sont désignés comme puits de contrôle, aux côtés de deux autres puits répartis uniformément au milieu de la plaque à B4 et C3. Lors de l’analyse des sphéroïdes XF, nous avons constaté un écart important dans les données initialement collectées à l’aide des directives du fabricant. Ceci malgré l’inclusion des étapes nécessaires pour assurer la prééquilibration du test à la température et à la teneur en CO2 avant de commencer l’acquisition des données, donnant souvent des valeurs négatives pour l’OCR après l’injection de certains inhibiteurs respiratoires (Figure 8).
Nous avons constaté que ces observations étaient probablement dues à des effets de bord sur la microplaque du test sphéroïde. Dans la figure 8, nous avons constaté que la redistribution des puits de contrôle de fond à travers la microplaque, les données XF ont été ajustées environ 2 fois. Les deux raisons les plus probables sont (1) dues aux effets d’évaporation au niveau des puits de bord, ce qui entraîne une chambre de volume total plus petite pour la sonde XFe96 à échantillonner, et (2) à des équilibres de température inadéquats entre les puits désignés pour la correction de fond et les puits d’échantillonnage, ce qui donne des ensembles de données qui masquent ou gonflent excessivement l’OCR. Pour éviter de tels résultats, il est donc recommandé, en particulier dans le contexte de l’analyse des sphéroïdes, que les utilisateurs redistribuent les puits destinés à la correction de fond sur l’ensemble de la microplaque du test sphéroïde et prennent les mesures nécessaires pour pré-équilibrer leur essai avant d’acquérir des données XF.
Normalisation des données
En plus de fournir un protocole détaillé pour sonder le métabolisme énergétique mitochondrial des sphéroïdes 3D uniques avec la technologie XF, cet article présente également des moyens possibles de normaliser les données de fréquence respiratoire mitochondriale obtenues avec des sphéroïdes 3D. À l’aide des données de fréquence respiratoire obtenues avec des sphéroïdes MCF-7 cultivés à différentes densités d’ensemencement cellulaire (figure 3), nous présentons les fréquences respiratoires mitochondriales basales des sphéroïdes MCF-7 de taille et de diamètre croissants lorsqu’elles sont normalisées à la densité d’ensemencement cellulaire initiale, au volume sphéroïde et à la teneur en ADNds (figure 9). La méthode de normalisation appropriée est primordiale pour l’interprétation précise des ensembles de données XF, en particulier lors de la comparaison de modèles de sphéroïdes 3D in vitro et de différents types de cellules. Une mauvaise normalisation peut conduire à des résultats erronés qui ne peuvent tout simplement pas être comparés entre les ensembles de données. La teneur en protéines n’est pas préférée pour la normalisation des données XF sphéroïdes, car les prétraitements peuvent avoir un impact sur les taux de synthèse des protéines sans effet significatif sur la fréquence respiratoire. De plus, des quantités significatives et incohérentes de protéines peuvent se lier à des microplaques sphéroïdes lors de la lyse cellulaire, introduisant une variation de la teneur en protéines entre les puits. Cela peut être encore compliqué dans les analyses XF utilisant des sphéroïdes ou des cellules non adhérentes qui nécessitent des colles biomoléculaires pour se lier, qui peuvent contenir des protéines.
Contrairement à la teneur en protéines intracellulaires, la teneur en ADN nucléaire est indépendante du type cellulaire et est proportionnelle au nombre de cellules (Figure 9D) - une approche plus précise et moins longue que la désagrégation des sphéroïdes pour la quantification du nombre de cellules. Inversement, Yepéz et al.55, en effectuant des analyses XF dans des monocouches de cellules fibroblastiques, ont constaté que la normalisation des données XF en nombre de cellules introduisait une plus grande dispersion des données qu’avant la normalisation. La teneur en ADN nucléaire est indépendante de l’état différencié ou du phénotype et est donc plus précise pour la normalisation des données sphéroïdes dans les tests XF que la teneur en protéines. Le contenu de l’ADN a également été une stratégie éprouvée pour l’analyse d’autres ensembles de données liés au métabolisme56. Cependant, il est important de noter que la teneur en ADN nucléaire est quantifiée à partir de toutes les cellules présentes dans le sphéroïde; par conséquent, la normalisation de la teneur en ADN n’est pas recommandée pour les ensembles de données XF dans lesquels les sphéroïdes subissent des traitements pouvant entraîner une perte importante de viabilité cellulaire. Pour de tels ensembles de données, si possible, la normalisation de la viabilité cellulaire est préférable, ou les données peuvent être corrigées de base à la respiration basale.
Utiliser la capacité respiratoire inutilisée comme exemple de l’importance de la normalisation des données
La capacité respiratoire de réserve est une mesure du taux de capacité respiratoire mitochondriale maximale moins la fréquence respiratoire mitochondriale basale (Figure 6). Cependant, le problème avec la déclaration de données de ce type sous forme de taux, c’est-à-dire de pmolO2 / min / puits dans certaines expériences, est que les données sont dépourvues de normalisation. Même si l’on normalise les données sphéroïdes en densité cellulaire / contenu en ADN, cela exclut souvent le paramètre clé qui doit être normalisé pour la densité mitochondriale dans les cellules. Étant donné qu’un changement de densité mitochondriale entraînera un changement proportionnel de la respiration basale et maximale, la capacité inutilisée augmentera également. Par exemple, si l’OCR sphéroïdebasal est de 200 et l’OCRmax est de 400, la capacité inutilisée est de 200; si l’OCRbasal est de 100 et l’OCRmax, la capacité de réserve est également de 100; cependant, en pourcentage, ils sont tous les deux de 50% du maximum (ou 100% de la base). Par conséquent, la capacité inutilisée n’est pas modifiée entre ces deux exemples, malgré des différences de taux de 200 et 100 lorsqu’elles sont calculées en pmols O2/min/puits. Les valeurs normalisées en interne sont plus fiables et plus perspicaces pour rendre les données XF plus comparables entre les études et les projets. Pour ce faire, pour la capacité respiratoire inutilisée, nous avons choisi de la présenter en pourcentage de la respiration maximale au lieu d’un taux absolu. Cela pourrait également être présenté comme un pourcentage de la respiration basale. Ce serait le cas si vous travaillez avec des cellules ou des sphéroïdes. Cependant, étant donné que l’emplacement du sphéroïde dans la plaque de micropuit peut modifier l’OCR absolue, mais pas les changements relatifs avec les inhibiteurs ou les découpleurs, il est plus important d’examiner les réponses normalisées en interne chez les sphéroïdes en tant que changement de pli ou pourcentages.
Les modèles sphéroïdes générés ici présentent une gamme de types de cellules et d’architecture qui ne peuvent pas être capturés dans les modèles 2D classiques. Il s’agit notamment de l’arrangement spatial hétérogène des cellules en trois dimensions, de l’amélioration des contacts cellule-cellule (p. ex., formation de jonctions lacunaires et de matrices extracellulaires) et des gradients biochimiques à travers le diamètre du sphéroïde (p. ex., gradients de pH, accès de la diffusion de l’oxygène aux nutriments). L’utilisation du flux extracellulaire pour étudier la biologie des sphéroïdes in vitro pourrait permettre d’identifier des cibles optimales pour les thérapies médicamenteuses grâce à des observations de perturbation métabolique. Ceux-ci pourraient être extrapolés des sphéroïdes in vitro aux tumeurs in vivo et identifier les voies qui peuvent cibler le métabolisme sphéroïde-tumoral, par exemple, l’utilisation des glucides pendant la croissance des sphéroïdes. Les modalités thérapeutiques peuvent être efficaces pour cibler les sphéroïdes dans les phases de croissance précoces, mais s’avèrent moins efficaces dans les phases ultérieures de la croissance des sphéroïdes à mesure que la complexité du réseau métabolique mûrit. Pour conclure, le développement de modèles de culture cellulaire 3D et de technologies d’analyse sophistiquées dans la recherche biologique continuera d’être un domaine dynamique et en évolution rapide avec un potentiel inégalé. L’analyse du flux extracellulaire des sphéroïdes de culture cellulaire in vitro pourrait être utilisée comme méthode de recherche de pointe pour faire progresser les résultats de la recherche qui pourraient être extrapolés pour mieux comprendre la biologie pertinente pour l’homme, réduire l’utilisation de modèles animaux dans la recherche et améliorer la recherche centrée sur le patient.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Les auteurs n’ont aucun conflit d’intérêts à déclarer.
Acknowledgments
N.J.C a reçu un BBSRC MIBTP CASE Award avec Sygnature Discovery Ltd (BB/M01116X/1, 1940003)
Materials
| Name | Company | Catalog Number | Comments |
| A549 | ECACC | #86012804 | Lung carcinoma cell line |
| Agilent Seahorse XF RPMI Medium, pH 7.4 | Agilent Technologies Inc. | 103576-100 | XF assay medium with 1 mM HEPES, without phenol red, sodium bicarbonate, glucose, L-glutamine, and sodium pyruvate |
| Agilent Seahorse XFe96 Extracellular Flux Analyzer | Agilent Technologies Inc. | - | Instrument for measuring rates of spheroid oxygen uptake in single spheroids |
| Antimycin A | Merck Life Science | A8674 | Mitochondrial respiratory complex III inhibitor |
| BAM15 | TOCRIS bio-techne | 5737 | Mitochondrial protnophore uncoupler |
| Black-walled microplate | Greiner Bio-One | 655076 | For fluorescence-based assays |
| CELLSTAR cell-repellent surface 96 U well microplates | Greiner Bio-One | 650970 | Microplates for generating spheroids |
| CellTiter-Glo 3D Cell Viability Assay | Promega | G9681 | Assay for the determination of cell viability in 3D microtissue spheroids |
| Cultrex Poly-D-Lysine | R&D Systems a biotechne brand | 3439-100-01 | Molecular cell adhesive for coating XFe96 spheroid microplates to facillitate attachment of spheroids |
| D-(+)-Glucose | Merck Life Sciences | G8270 | Supplement for cell culture growth and XF assay medium |
| Dulbecco’s Modified Eagle Medium (DMEM) | Gibco | 11885084 | Culture medium for HepG2/C3A spheroids |
| EVOS XL Core Imaging System | Thermo Fisher Scientific | AMEX1000 | Phase-contrast imaging microscope |
| EZ-PCR Mycoplasma test kit | Biological Industries | 20-700-20 | Mycoplasma screening in cell cultures |
| FIJI Is Just Image J | Analysis of collated images | ||
| Foetal bovine serum | Merck Life Science | F7524 | Supplement for cell culture medium |
| HepG2/C3A | ATCC | #CRL-10741 | Hepatic carcinoma cell line, a clonal derivative of the parent HepG2 cell line |
| Lactate-Glo | Promega | J5021 | Assay for measurement of lactate within spheorid culture medium |
| L-glutamine (200 mM solution) | Merk Life Sciences | G7513 | Supplement for cell culture growth and XF assay medium |
| M50 Stereo microscope | Leica Microsytems | LEICAM50 | Stereo dissection micrscope; used for spheorid handling |
| MCF-7 | ECACC | #86012803 | Breast adenocarcinoma cell line |
| Oligomycin from Streptomyces diastatochromogenes | Merck Life Science | O4876 | ATP Synthase Inhibitor |
| Penicilin-Streptomycin | Gibco | 15140122 | Antibiotics added to cell culture medium |
| Quant-iT PicoGreen dsDNA Assay Kit | Initrogen | P7589 | Analysis of dsDNA in spehroids |
| Rotenone | Merck Life Science | R8875 | Mitochondrial Respiratory Complex I Inhibitor |
| RPMI 1640 | Gibco | 21875091 | Culture medium for A549, MCF7, and SK-OV-3 spheroids |
| Seahorse Analytics | Agilent Technologies Inc. | Build 421 | https://seahorseanalytics.agilent.com |
| Seahorse XFe96 Spheroid FluxPak | Agilent Technologies Inc. | 102905-100 | Each Seahorse XFe96 Spheroid FluxPak contains: 6 Seahorse XFe96 Spheroid Microplates (102978-100), 6 XFe96 sensor cartridges, and 1 bottle of Seahorse XF Calibrant Solution 500 mL (100840-000) |
| Serological pipette: 5, 10, and 25 mL | Greiner Bio-One | 606107; 607107; 760107 | Consumables for cell culture |
| SK-OV-3 | ECACC | #HTB-77 | Ovarian adenocarcinoma cell line |
| Sodium pyruvate (100 mM solution) | Merck Life Science | S8636 | Supplement for cell culture growth and XF assay medium |
| T75 cm2 cell culture flask | Greiner Bio-One | 658175 | Tissue culture treated flasks for maintaining cell cultures |
| TrypLExpress | Gibco | 12604-021 | Cell dissociation reagent |
| Wave controller software | Agilent Technologies Inc. | - | |
| Wide orifice tip | STARLAB International GmbH | E1011-8400 | Pipette tips with wide opening for spheroid handling |
References
- Correa de Sampaio, P., et al. A heterogeneous in vitro three dimensional model of tumour-stroma interactions regulating sprouting angiogenesis. PLoS One. 7 (2), 30753 (2012).
- Amann, A., et al. Development of an innovative 3D cell culture system to study tumour-stroma interactions in non-small cell lung cancer cells. PLoS One. 9 (3), 92511 (2014).
- Russell, S., Wojtkowiak, J., Neilson, A., Gillies, R. J. Metabolic profiling of healthy and cancerous tissues in 2D and 3D. Scientific Reports. 7 (1), 15285 (2017).
- Zanoni, M., et al. 3D tumor spheroid models for in vitro therapeutic screening: a systematic approach to enhance the biological relevance of data obtained. Scientific Reports. 6, 19103 (2016).
- Song, Y., et al. Patient-derived multicellular tumor spheroids towards optimized treatment for patients with hepatocellular carcinoma. Journal of Experimental and Clinica Cancer Research. 37 (1), 109 (2018).
- Courau, T., et al. Cocultures of human colorectal tumor spheroids with immune cells reveal the therapeutic potential of MICA/B and NKG2A targeting for cancer treatment. Journal for ImmunoTherapy of Cancer. 7 (1), 74 (2019).
- Ivanova, E., et al. Use of ex vivo patient-derived tumor organotypic spheroids to identify combination therapies for HER2 mutant non-small cell lung cancer. Clinical Cancer Research. 26 (10), 2393-2403 (2020).
- Mandon, M., Huet, S., Dubreil, E., Fessard, V., Le Hegarat, L. Three-dimensional HepaRG spheroids as a liver model to study human genotoxicity in vitro with the single cell gel electrophoresis assay. Scientific Reports. 9 (1), 10548 (2019).
- Stampar, M., et al. Hepatocellular carcinoma (HepG2/C3A) cell-based 3D model for genotoxicity testing of chemicals. Science of the Total Environment. 755, 143255 (2020).
- Coltman, N. J., et al. Application of HepG2/C3A liver spheroids as a model system for genotoxicity studies. Toxicology Letters. 345, 34-45 (2021).
- Tchoryk, A., et al. Penetration and uptake of nanoparticles in 3D tumor spheroids. Bioconjugate Chemistry. 30 (5), 1371-1384 (2019).
- Leite, P. E. C., et al. Suitability of 3D human brain spheroid models to distinguish toxic effects of gold and poly-lactic acid nanoparticles to assess biocompatibility for brain drug delivery. Partical Fibre Toxicology. 16 (1), 22 (2019).
- Elje, E., et al. Hepato(Geno)toxicity assessment of nanoparticles in a HepG2 liver spheroid model. Nanomaterials. 10 (3), 545 (2020).
- Conway, G. E., et al. Adaptation of the in vitro micronucleus assay for genotoxicity testing using 3D liver models supporting longer-term exposure durations. Mutagenesis. 35 (4), 319-330 (2020).
- Wang, Z., et al. HepaRG culture in tethered spheroids as an in vitro three-dimensional model for drug safety screening. Journal of Applied Toxicology. 35 (8), 909-917 (2015).
- Proctor, W. R., et al. Utility of spherical human liver microtissues for prediction of clinical drug-induced liver injury. Archives of Toxicology. 91 (8), 2849-2863 (2017).
- Basharat, A., Rollison, H. E., Williams, D. P., Ivanov, D. P. HepG2 (C3A) spheroids show higher sensitivity compared to HepaRG spheroids for drug-induced liver injury (DILI). Toxicology and Applied Pharmacology. 408, 115279 (2020).
- Benning, L., Peintner, A., Finkenzeller, G., Peintner, L. Automated spheroid generation, drug application and efficacy screening using a deep learning classification: a feasibility study. Scientific Reports. 10 (1), 11071 (2020).
- Mittler, F., et al. High-content monitoring of drug effects in a 3D spheroid model. Frontiers in Oncology. 7, 293 (2017).
- Brand, M. D., Nicholls, D. G.
- Benz, R., McLaughlin, S. The molecular mechanism of action of the proton ionophore FCCP (carbonylcyanide p-trifluoromethoxyphenylhydrazone). Biophysical Journal. 41 (3), 381-398 (1983).
- Kasianowicz, J., Benz, R., McLaughlin, S. The kinetic mechanism by which CCCP (carbonyl cyanide m-chlorophenylhydrazone) transports protons across membranes. The Journal of Membrane Biology. 82 (2), 179-190 (1984).
- Kenwood, B. M., et al. Identification of a novel mitochondrial uncoupler that does not depolarize the plasma membrane. Molecular Metabolism. 3 (2), 114-123 (2013).
- Mitchell, P. Coupling of phosphorylation to electron and hydrogen transfer by a chemi-osmotic type of mechanism. Nature. 191, 144-148 (1961).
- Alexopoulos, S. J., et al. Mitochondrial uncoupler BAM15 reverses diet-induced obesity and insulin resistance in mice. Nature Communications. 11 (1), 2397 (2020).
- Chen, S. -Y., et al. Mitochondrial uncoupler SHC517 reverses obesity in mice without affecting food intake. Metabolism - Clinical and Experimental. 117, 154724 (2021).
- Goedeke, L., Shulman, G. I. Therapeutic potential of mitochondrial uncouplers for the treatment of metabolic associated fatty liver disease and NASH. Molecular Metabolism. 46, 101178 (2021).
- Hill, B. G., et al. Integration of cellular bioenergetics with mitochondrial quality control and autophagy. Biological chemistry. 393 (12), 1485-1512 (2012).
- Demine, S., Renard, P., Arnould, T. Mitochondrial uncoupling: a key controller of biological processes in physiology and diseases. Cells. 8 (8), 795 (2019).
- Wang, J., et al. Uncoupling effect of F16 is responsible for its mitochondrial toxicity and anticancer activity. Toxicological Sciences. 161 (2), 431-442 (2018).
- Tretter, L., Chinopoulos, C., Adam-Vizi, V. Plasma membrane depolarization and disturbed Na+ homeostasis induced by the protonophore carbonyl cyanide-p-trifluoromethoxyphenyl-hydrazon in isolated nerve terminals. Molecular Pharmacology. 53 (4), 734-741 (1998).
- Connop, B. P., Thies, R. L., Beyreuther, K., Ida, N., Reiner, P. B. Novel effects of FCCP [carbonyl cyanide p-(trifluoromethoxy) phenylhydrazone] on amyloid precursor protein processing. Journal of neurochemistry. 72 (4), 1457-1465 (1999).
- Stöckl, P., et al. Partial uncoupling of oxidative phosphorylation induces premature senescence in human fibroblasts and yeast mother cells. Free Radical Biology and Medicine. 43 (6), 947-958 (2007).
- Firsov, A. M., et al. Protonophoric action of BAM15 on planar bilayers, liposomes, mitochondria, bacteria and neurons. Bioelectrochemistry. 137, 107673 (2021).
- Dranka, B. P., Hill, B. G., Darley-Usmar, V. M. Mitochondrial reserve capacity in endothelial cells: The impact of nitric oxide and reactive oxygen species. Free Radical Biology and Medicine. 48 (7), 905-914 (2010).
- Eilenberger, C., Rothbauer, M., Ehmoser, E. K., Ertl, P., Kupcu, S. Effect of spheroidal age on sorafenib diffusivity and toxicity in a 3D HepG2 spheroid model. Scientific Reports. 9 (1), 4863 (2019).
- vanden Brand, D., Veelken, C., Massuger, L., Brock, R. Penetration in 3D tumor spheroids and explants: Adding a further dimension to the structure-activity relationship of cell-penetrating peptides. Biochimica et Biophysica Acta (BBA) - Biomembranes. 1860 (6), 1342-1349 (2018).
- Niora, M., et al. Head-to-head comparison of the penetration efficiency of lipid-based nanoparticles into tumor spheroids. ACS Omega. 5 (33), 21162-21171 (2020).
- Millard, M., et al. Drug delivery to solid tumors: the predictive value of the multicellular tumor spheroid model for nanomedicine screening. International Journal of Nanomedicine. 12, 7993-8007 (2017).
- Ruas, J. S., et al. Underestimation of the maximal capacity of the mitochondrial electron transport system in oligomycin-treated cells. PLoS One. 11 (3), 0150967 (2016).
- Benton, G., DeGray, G., Kleinman, H. K., George, J., Arnaoutova, I. In vitro microtumors provide a physiologically predictive tool for breast cancer therapeutic screening. PLoS One. 10 (4), 0123312 (2015).
- Hirpara, J., et al. Metabolic reprogramming of oncogene-addicted cancer cells to OXPHOS as a mechanism of drug resistance. Redox Biology. 25, 101076 (2019).
- Ware, M. J., et al. Generation of homogenous three-dimensional pancreatic cancer cell spheroids using an improved hanging drop technique. Tissue Engineering. Part C, Methods. 22 (4), 312-321 (2016).
- Song, Y., et al. TGF-β-independent CTGF induction regulates cell adhesion mediated drug resistance by increasing collagen I in HCC. Oncotarget. 8 (13), 21650-21662 (2017).
- Wrzesinski, K., et al. HepG2/C3A 3D spheroids exhibit stable physiological functionality for at least 24 days after recovering from trypsinisation. Toxicology Research. 2 (3), 163-172 (2013).
- Gaskell, H., et al. Characterization of a functional C3A liver spheroid model. Toxicology Research. 5 (4), 1053-1065 (2016).
- Takahashi, Y., et al. 3D spheroid cultures improve the metabolic gene expression profiles of HepaRG cells. Bioscience Reports. 35 (3), 00208 (2015).
- Hendriks, D. F. G., Puigvert, L. F., Messner, S., Mortiz, W., Ingelman-Sundberg, M. Hepatic 3D spheroid models for the detection and study of compounds with cholestatic liability. Scientific Reports. 6, 35434 (2016).
- Leung, B. M., Lesher-Perez, S. C., Matsuoka, T., Moraes, C., Takayama, S. Media additives to promote spheroid circularity and compactness in hanging drop platform. Biomaterials Science. 3 (2), 336-344 (2015).
- Cavo, M., et al. A synergic approach to enhance long-term culture and manipulation of MiaPaCa-2 pancreatic cancer spheroids. Scientific Reports. 10 (1), 10192 (2020).
- Carlsson, J., Yuhas, J. M.
- Costa, E. C., Gaspar, V. M., Coutinho, P., Correia, I. J. Optimization of liquid overlay technique to formulate heterogenic 3D co-cultures models. Biotechnology and Bioengineering. 111 (8), 1672-1685 (2014).
- Lundholt, B. K., Scudder, K. M., Pagliaro, L. A simple technique for reducing edge effect in cell-based assays. Journal of Biomolecular Screening. 8 (5), 566-570 (2003).
- Zhang, X. D., et al. The use of strictly standardized mean difference for hit selection in primary RNA interference high-throughput screening experiments. Journal of Biomolecular Screening. 12 (4), 497-509 (2007).
- Yepez, V. A., et al. OCR-Stats: Robust estimation and statistical testing of mitochondrial respiration activities using Seahorse XF Analyzer. PLoS One. 13 (7), 0199938 (2018).
- Silva, L. P., et al. Measurement of DNA concentration as a normalization strategy for metabolomic data from adherent cell lines. Analytical Chemistry. 85 (20), 9536-9542 (2013).
Tags
Recherche sur le cancer numéro 180Erratum
Formal Correction: Erratum: Exploring Mitochondrial Energy Metabolism of Single 3D Microtissue Spheroids using Extracellular Flux Analysis
Posted by JoVE Editors on 03/11/2022.
Citeable Link.
An erratum was issued for: Exploring Mitochondrial Energy Metabolism of Single 3D Microtissue Spheroids using Extracellular Flux Analysis. The Representative Results section was updated.
Figure 5 was updated from:
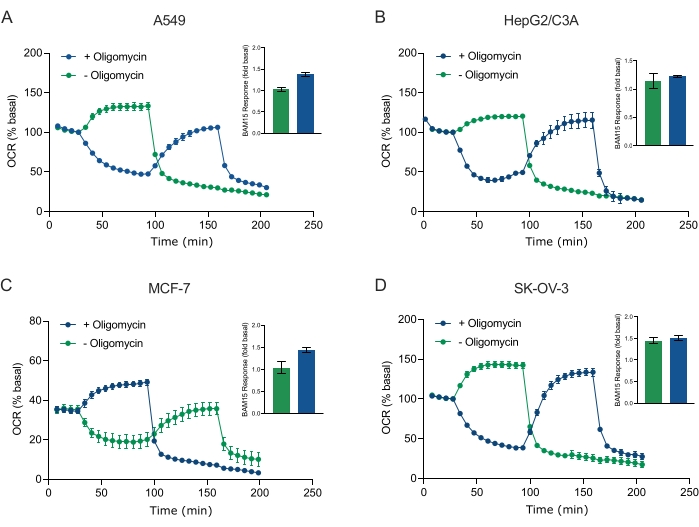
Figure 5: Single or sequential injection of mitochondrial respiratory compounds. Cancer-cell-derived spheroids of MCF-7, HEPG2/C3A, SK-OV-3, and A549 were placed into wells of an XFe96 spheroid microplate in XF RPMI and probed for OCR using the Agilent Seahorse XFe96 analyzer. OCR was measured 5x, after which 2 µg/mL oligomycin (injection Port A: green trace) or 5 µM BAM15 (injection Port A: blue trace or injection port B: green trace) to inhibit the mitochondrial ATP synthase and determine maximal respiratory capacity, respectively. Kinetic OCR data are expressed as % basal (A-D). Maximal respiratory capacity (OCRmax) was calculated as a factor of basal OCR by the equation: OCRmax = OCRBAM15 / OCRbasal. OCRmax was obtained from OCR averages across measurement cycles 8-10 post BAM15 injection with (green bars) and without (blue bars) oligomycin. Data are averages ± SEM from 3-8 individual well replicates across the spheroid assay microplate. Abbreviations: OCR = oxygen consumption rate. Please click here to view a larger version of this figure.
to:
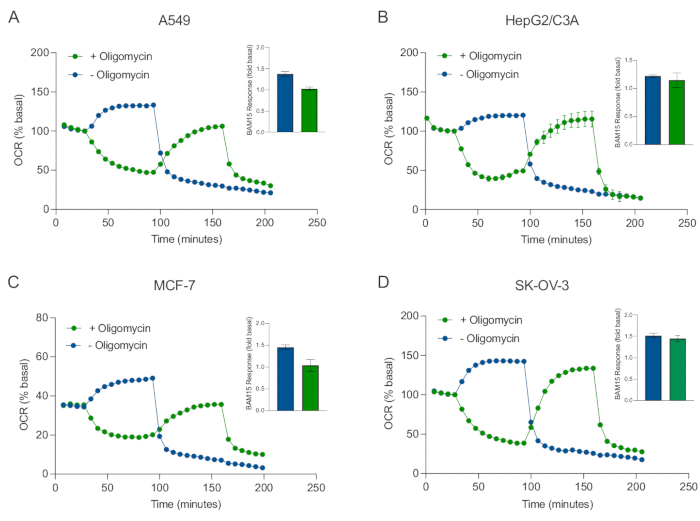
Figure 5: Single or sequential injection of mitochondrial respiratory compounds. Cancer-cell-derived spheroids of MCF-7, HEPG2/C3A, SK-OV-3, and A549 were placed into wells of an XFe96 spheroid microplate in XF RPMI and probed for OCR using the Agilent Seahorse XFe96 analyzer. OCR was measured 5x, after which 2 µg/mL oligomycin (injection Port A: green trace) or 5 µM BAM15 (injection Port A: blue trace or injection port B: green trace) to inhibit the mitochondrial ATP synthase and determine maximal respiratory capacity, respectively. Kinetic OCR data are expressed as % basal (A-D). Maximal respiratory capacity (OCRmax) was calculated as a factor of basal OCR by the equation: OCRmax = OCRBAM15 / OCRbasal. OCRmax was obtained from OCR averages across measurement cycles 8-10 post BAM15 injection with (green bars) and without (blue bars) oligomycin. Data are averages ± SEM from 3-8 individual well replicates across the spheroid assay microplate. Abbreviations: OCR = oxygen consumption rate. Please click here to view a larger version of this figure.
