1. Yeast Strains
Strains with wild-type SGS1 TOP3 (WT; W303-1A, genotype, MATa ade2-1 canl- 100 his3-11,15 leu2-3,112 trpl-l ura3-1) [2], a sgs1 mutant (W1292-3C; genotype MATa SUP4-o::URA3 sgs1-25 ade2-1 can1-100 his3-11,15 leu2-3,112 trp1-1 ura3-1 rad5-535) and a sgs1 top3 mutant ( W1058-11C, genotype, MATa SUP4-o::URA3 sgs1-25 top3-2::HIS3 ade2-1 can1-100 his3-11,15 leu2-3,112 trp1-1 ura3-1 rad5-535) have been characterized [3] and were kindly provided by Dr. Rodney Rothstein (Columbia University).
2. Plasmid DNA constructions
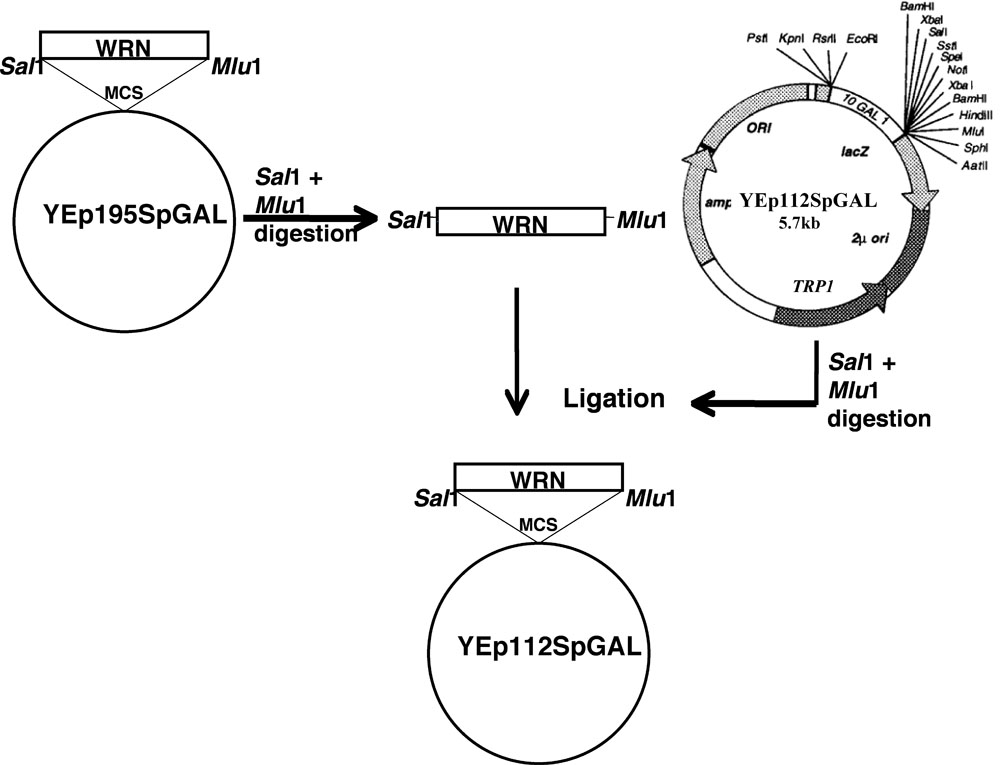
The WRN gene was cloned in YEp112SpGAL following the scheme as discussed.
- Site-directed mutations of WRN (E84A exonuclease dead, K577M helicase dead, R834C, and K1016A) in the plasmid YEp195SpGAL-WRN [4] were constructed using mutagenic primers and a standard protocol from Quickchange II XL site-directed mutagenesis kit (Stratagene) by Lofstrand labs (Gaithersburg, MD).
- DNA fragments encoding WRN and its associated variants were gel purified from the respective YEp195SpGAL-WRN constructs after double digestion with SalI and MluI.
- Gel purified fragments were then cloned into the SalI MluI sites of vector YEp112SpGAL, a 2 μm multi-copy plasmid containing a TRP1 selectable marker to construct YEp112SpGAL WRN or WRN variants under the control of a gal-inducible promoter as shown in Figure 1.
- The constructs so obtained were named as YEp112SpGAL-WRN, YEp112SpGAL-WRN E84A, YEp112SpGAL-WRN K577M, YEp112SpGAL-WRN K1016A, and YEp112SpGAL-WRN R834C.
3. Transformation
Yeast cultures were grown using standard protocol and transformations were performed using a Lithium Acetate-based protocol by Gietz et al [5]. Briefly, the following steps were performed.
- Cultures of the above mentioned strains were grown overnight in 5 ml YPD (yeast extract-peptone-dextrose) medium. Cultures were reinoculated in 50 ml YPD medium and were allowed to grow until an OD600 of 1.0 – 1.2 was reached.
- Cultures were harvested, washed with 20 ml water, and resuspended in 1 ml of 100 mM lithium acetate (LiAc). Cells were incubated at 30°C for 10 min.
- Cell pellet was resuspended in 100 mM LiAc to a final volume of 500 μl, which is enough to do ten transformations. 50 μl of the cell suspension was aliquoted and cell pellet was collected by centrifugation.
- To the cell pellet was added in the following order: 250 μl 50% Polyethylene glycol (PEG), 36 μl of 1 M LiAc, 25 μl of 2 mg/ml ssDNA (salmon sperm DNA), 5 ml of plasmid DNA (100 ng 1000 ng), and 50 μl water. Cell pellet was vortexed for 1 min.
- Cells were then incubated at 30°C for 30 min followed by a heat shock at 42°C for 20 min.
- Cells were harvested and resuspended in 1 ml water. Since the plasmid YEp112SpGAL contains trp as a selectable marker, all the mutant strains harboring the construct would be trp proficient. Therefore, transformants were selected by plating the suspension on SC (Synthetic complete) glucose media lacking Trp and incubating the plates at 30°C for 3-4 days until the colonies appear.
4. Genetic analysis of the slow growth phenotype restoration in WRN transformed sgs1 top3 strain.
1. Streak analysis:
- To examine the effect of WRN expression on the growth of wild-type parental (W303-1A), sgs1, or sgs1 top3 strains, the corresponding strains with YEp112SpGAL or YEp112SpGAL-WRN were streaked onto SC minus Trp plates containing either 2% glucose (glu) or 2% galactose (gal). As a control, sgs1 top3/YEp112SpGAL-SGS1 was included. Plates were incubated at 30°C for 2 days.
- To determine the ability of WRN to affect growth rate of sgs1 top3 strain at lower WRN protein expression levels, the sgs1 top3 strain transformed with YEp112SpGAL, YEp112SpGAL-WRN, or YEp112SpGAL-SGS1 was streaked onto SC minus Trp plates containing varying concentrations of gal from as low as 0.005% to as high as 2%. Plates were then incubated at 30°C for 2 days.
- To assess the functional requirements of WRN to restore the slow growth phenotype in the sgs1 top3 background, sgs1 top3 transformed with YEp112SpGAL-WRN (E84A, K577M, R834C, and K1016A) were streaked onto SC minus Trp plates containing either glu or gal at the indicated concentrations. For controls, sgs1 top3 strain transformed with YEp112SpGAL, YEp112SpGAL-WRN or YEp112Sp GAL-SGS1 was included. Plates were then incubated at 30°C for 2 days.
2. Liquid culture analysis:
- To evaluate the growth of transformed sgs1 top3 strains in liquid culture, yeast cells were grown in SC raf minus Trp at 30°C overnight. Cultures were reinoculated in SC raf minus Trp and grown at 30°C to early log phase (OD600 of ~ 0.5). Cultures were then reinoculated at OD0.05 in SC media containing 2% gal and growth was followed by measuring absorbance at the indicated time intervals.
5. Genetic analysis of hydroxyurea or methylmethane-sulfonate sensitivity in WRN transformed yeast strains.
- To determine the effect of WRN expression on the MMS and HU sensitivity of sgs1 or sgs1 top3 strains, strains transformed with YEp112SpGAL or YEp112SpGAL-WRN were grown in SC raf minus Trp at 30°C.
- Cultures were reinoculated in SC raf minus Trp and grown at 30°C to early log phase (OD600 of ~0.6 to 0.8).
- Ten-fold serial dilutions of these strains were prepared using SC medium lacking Trp. From an exponentially growing culture, 5 x 106 cells were taken and diluted serially up to 10,000-fold. Four microliters of each dilution was spotted onto SC gal minus Trp plates containing the indicated concentrations of HU or MMS. Plates were incubated at 30°C.
- As a control, wild-type parental strain (W303-1A) transformed with YEp112SpGAL or YEp112SpGAL-WRN, or sgs1 top3/ YEp112SpGAL-SGS1 was treated as described above.
6. Cell cycle distribution of WRN transformed sgs1 top3 strain.
- To study the cell cycle distribution of WRN transformed sgs1 top3 strain, cultures of sgs1 top3 transformed with YEp112SpGAL, YEp112SpGAL-WRN, or YEp112SpGAL-SGS1, and the vector-transformed wild-type parental strain (W303-1A) were grown in SC raf minus trp at 30°C for overnight.
- Cultures were reinoculated at OD0.05 and allowed to grow til early log phase (OD600 of 0.5 -0.6). WRN expression was induced by adding gal to 2% final concentration. Cultures were then allowed to grow at 30°C for 6 h.
- Cultures were then processed for DAPI (4′, 6-diamidino-2-phenylindole) staining.
- Cultures were harvested by centrifugation at 5800 x g for 10 min at room temperature (RT).
- Cells were washed once with 1X phosphate-buffered saline (PBS) and were then fixed in 70% ethanol for 20 min at RT.
- Fixed cells were washed twice with 1 X PBS, and finally resuspended in 1X PBS.
- Cells were placed on slide and mounted in Vectashield mounting medium with DAPI (1 μg/ml) (Vector,USA). Cells were examined with an Axiovert 200 M microscope (Zeiss; 100x lens) and composite differential interference contrast (DIC). Fluorescence images were analyzed using AxioVision, version 3.0 program (Zeiss).
7. Western blot analyses.
1. To determine the protein expression of WRN or the WRN variants in transformed sgs1 or sgs1 top3 strains, lysates were prepared by the following method and proteins analyzed by immunblotting.
- Transformed sgs1 or sgs1 top3 strains were grown in SC raf minus Trp at 30°C for overnight.
- Cultures were reinoculated and incubated at 30°C until early log phase (OD600 of 0.5 – 0.8) and then induced by adding gal to 2% final concentration. Cultures were then allowed to grow at 30°C for 6 h.
- Cells (3 ml) were collected by centrifugation and washed with PBS. Cell pellet was resuspended in alkaline lysis buffer [50 mM NaOH, pH 10.5, 2 mM EDTA, 1 mM phenylmethylsulfonyl fluoride (PMSF), 2% SDS, 10% glycerol, 5% 2-mercaptoethanol and protease inhibitors (Roche Molecular Biochemicals)], boiled for 5 min, clarified by centrifugation, and neutralized with 1 M HCl.
- Proteins from equivalent amounts of cell lysate were resolved on 8-16% polyacrylamide SDS gels.
2. To determine the level of expression of WRN protein under the influence of varying concentrations of galactose, cultures were processed as follows.
- Transformed sgs1 top3 strains were grown in SC raf minus Trp at 30°C for overnight.
- Cultures were reinoculated and incubated at 30°C until early log phase (OD600 of 0.5 – 0.8) and then induced by adding gal to 2% final concentration. Cultures were then allowed to grow at 30°C for 6 h.
- Cells were harvested by centrifugation and resuspended in buffer (50 mM Tris-Cl, pH 7.5, 1% sodium deoxycholate, 1% triton X-100, 0.1% sodium dodecyl sulfate [SDS]) containing protease inhibitors (0.05% PMSF, 0.05 μg/ml leupeptin).
- Glass beads (425 to 600 μm) were added to the mixture, and the cells were broken by vortexing.
- The resulting homogenate was centrifuged for 5 min at 15000 x g and the supernatant fraction was collected.
- Protein in the lysate fractions was estimated by Bradford method.
- Equal concentration of protein from different lysate fractions was resolved on 8-16% polyacrylamide SDS gels.
3. Expression of WRN or WRN mutant proteins was determined by Western blot using a WRN mouse monoclonal antibody directed against an epitope in a purified C-terminal fragment of WRN [6] (1:1000, Spring Valley Labs).
4. Primary antibody incubation was followed by incubation with a secondary (horseradish peroxidase-conjugated) antibody (1:5000). Blots were processed with an ECL Plus Western detection system per the manufacturer’s protocol.
5. For quantitative Western blot analysis, increasing concentrations of purified recombinant His-tagged full-length WRN protein were included on gels such that upon quantification of the blot a standard linear curve could be generated. Further, cell extract from equivalent amount of cells or 20 μg of the yeast lysate samples were loaded on gels to estimate the concentration of WRN.
6. ImageQuant analysis was then performed on gels and standard linear curve was generated.
7. Concentration of WRN protein at varying gal concentrations was estimated using standard linear curve generated above.
8. Representative Results
All strains used in this study are Trp auxotrophs. Therefore, sgs1, sgs1 top3 and wild type W303-1A strains transformed with YEp112SpGAL or YEp112SpGAL-WRN were selected on the basis of their ability to grow in presence of SC minus Trp media. To examine the effect of WRN expression on the growth of transformed sgs1 top3 mutant cells, the cultures were streaked onto plates containing SC minus Trp media with 2% gal to induce WRN expression. Top3 decatenates intertwined DNA molecules generated by Sgs1 helicase during replication [1, 2]; therefore, in the absence of Top3, torsional stress is not relieved resulting in slow growth and hyper-recombination. The genetic function of sgs1 is to suppress the slow growth phenotype of a top3 mutant. If WRN could substitute for SGS1 in genetic interaction with Top3, restoration of top3 slow growth phenotype in sgs1 top3 would be expected.
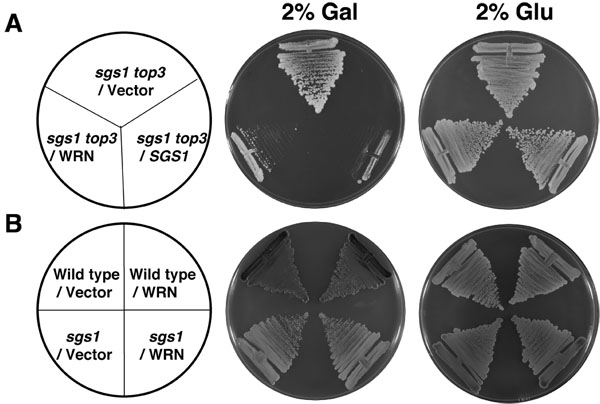
As shown in Figure 2A, the WRN transformed sgs1 top3 strain grew significantly more slowly compared to the vector transformed sgs1 top3 strain. sgs1 top3 transformed with the YEp112SpGAL-SGS1 plasmid was included as a positive control (Figure 2A), demonstrating that wild-type Sgs1 expressed in the sgs1 top3 mutant was able to genetically complement the growth phenotype. The transformed sgs1 top3 strains grew similarly in the absence of gal as shown for 2 days (Figure 2A). Expression of WRN had no effect on the growth of parental wild-type (W3031A) or sgs1 strains (Figure 2B), indicating that the effect of WRN on cell growth was specific to that observed in the sgs1 top3 mutant background. Genetic studies performed using WRN variants demonstrated that WRN helicase but not exonuclease activity was required for the restoration of top3 growth phenotype (Figure 3B). A naturally occurring missense polymorphism in WRN that interferes with helicase activity abolished its ability to restore top3 slow growth phenotype.
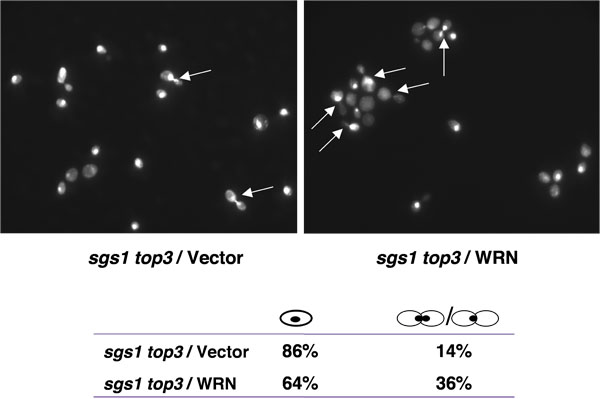
top3 mutant strains are delayed in the late S/G2 phase of the cell cycle [1], a characteristic that may account for their slow growth. Mutation of the SGS1 gene in the top3 background suppresses the delay in the S/G2 phase of the cell cycle. If WRN expression could restore the delay in the S/G2 phase of the cell cycle in sgs1 top3, an elevated population of large budded cells with undivided nuclei would be expected for sgs1 top3 mutant cells expressing WRN. As shown in Figure 4, the percentage of large budded cells was higher for sgs1 top3/WRN than sgs1 top3/vector, suggesting the restoration of the S/G2 delay characteristic of top3.
The sgs1 top3 double mutant is less sensitive to MMS or HU than the top3 single mutant [2]. Since WRN affected cell growth of sgs1 top3, we next examined its effect on sensitivity to methylmethane sulfonate (MMS, an alkylating agent) and hydroxyurea (HU, a replication inhibitor). sgs1 top3 / WRN displayed sensitivity to both the drugs comparable to sgs1 top3/ SGS1 (Figure 5). Genetic analysis of WRN variants revealed that WRN helicase /ATPase, but not WRN exonuclease activity, was required for its effect on sensitivity to MMS and HU.

Figure 1. Schematic presentation for cloning WRN / WRN variants in YEp112SpGAL vector. Please click here for a larger version of figure 1.

Figure 2. WRN expression in sgs1 top3 restores the slow growth phenotype of top3. Panel A, sgs1 top3 strain transformed with YEp112SpGAL or YEp112SpGAL WRN were streaked on an SC-Trp plate containing either 2% glu or 2% gal. As a control sgs1 top3 strain transformed with YEp112SpGAL SGS1 was streaked on both the plates. Plates were incubated at 30°C for 2 days and then photographed. Panel B, Wild type parental strain W303-1A or sgs1 strain transformed with YEp112SpGAL or YEp112SpGAL WRN were streaked on an SC-Trp plate either containing 2% glu or 2% gal. Plates were incubated at 30°C for 4 days and then photographed. Composition of the plates was as in Panel A and Panel B respectively. Please click here to see a larger version of figure 2.

Figure 3. WRN ATPase/helicase, but not exonuclease activity, is required to restore the slow growth phenotype of top3 in sgs1 top3 background. sgs1 top3 strain transformed with ATPase/helicase-dead (YEp195SpGAL WRN K577M), exonuclease-dead (YEp195SpGAL-WRN E84A), RQC mutant (YEp195SpGAL-WRN K1016A), or polymorphic mutant (YEp195SpGAL-WRN R834C) was streaked on SC-Trp plates containing either 2% glu (Panel C) or 2% gal (Panel B). Plates were incubated at 30°C for 2 days and then photographed. Composition of the plates was as in Panel A. Please click here to see a larger version of figure 3.

Figure 4. WRN expression induces S/G2 arrest in sgs1 top3 cells. Logarithmically growing cultures of sgs1 top3 strain transformed with YEp112SpGAL, YEp112SpGAL WRN, or YEp112SpGAL SGS1, and the vector-transformed wild-type parental strain were induced at 2% gal concentration for 6 h. Cultures were harvested, processed for DAPI staining as described in Materials and Methods and were observed using Axiovert 200 M microscope (Zeiss; 100x lens). Shown is the DAPI staining of the sgs1 top3 transformed with YEp112SpGAL (upper left) and with YEp112SpGAL WRN (upper right). Arrows show cells with undivided nuclei. Distribution of the cells in G1 (single cells) and S/G2 (budded cells) is shown in lower panel. Please click here to see a larger version of figure 4.

Figure 5. Effect of WRN expression on the MMS and HU sensitivity of sgs1 top3 strain. Logarithmically growing cultures of sgs1 top3 strain transformed with YEp112SpGAL , YEp112SpGAL WRN, exonuclease-dead (YEp112SpGAL-WRN E84A), ATPase/helicase-dead (YEp112SpGAL WRN K577M), RQC mutant (YEp112SpGAL-WRN K1016A), polymorphic mutant (YEp112SpGAL-WRN R834C), YEp112SpGAL SGS1 and vector transformed wild type parental strains were spotted in a ten-fold serial dilutions onto SC-Trp plates containing glu or gal and either MMS or HU at the indicated concentrations. Plates were incubated at 30°C for 2 days (control plates) and 4 days (MMS and HU plates) and then photographed. Please click here to see a larger version of figure 5.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}