

Summary
Ufikserede frosne vævsprøver indlagt i Optimal Cutting Temperatur medium (OCT) kan anvendes til undersøgelse af naturlige fordeling og glycosylering af secerneret slim. I denne tilgang vævsbehandling er minimal, og den naturlige præsentation af glycolipider, muciner og glycan-epitoper er bevaret. Vævssnit kan analyseres ved immunohistokemi ved anvendelse af fluorescens eller kromogen detektion.
Abstract
Muciner er komplekse og stærkt glycosyleret O-bundne glycoproteiner, som indeholder mere end 70% carbohydrat vægt 1-3. Secernerede muciner, produceret af slimceller og maveslimhinden, giver stilladset en mikrometer tykt slimlag at linjer epiteler i tarmen og luftvejene 3,4. Ud over muciner, også slim lag indeholder antimikrobielle peptider, cytokiner og immunglobuliner 5-9. Det slim lag er en vigtig del af vært medfødte immunitet, og danner den første linje i forsvaret mod invaderende mikroorganismer 8,10-12. Som sådan er slim genstand for talrige interaktioner med mikroorganismer, både bakterier og symbionter, og secernerede muciner danner en vigtig grænseflade for disse interaktioner. Undersøgelsen af sådanne biologiske vekselvirkninger involverer sædvanligvis histologiske metoder til vævs indsamling og farvning. De to mest almindeligt anvendte histologiske metoder til væv indsamling og preservtion i klinikken og i forskningslaboratorier er: formalinfiksering efterfulgt af paraffin embedding, og væv frysning, efterfulgt af indstøbning i kryo-beskyttelsesmiddel medier.
Paraffinindlejrede vævsprøver producere sektioner med optimale egenskaber til histologisk visualisering, herunder klarhed og veldefineret morfologi. Men i løbet af paraffin indlejring er en række epitoper er blevet ændret, og for at studere disse epitoper, vævssnittene skal bearbejdes yderligere med en af mange epitopgenfinding metoder 13. Secernerede muciner og lipider ekstraheres fra vævet i paraffin-indlejring clearing trin, som kræver forlænge inkubation med organiske opløsningsmidler (xylen eller Citrisolv). En sådan fremgangsmåde er optimal for undersøgelser, der fokuserer på arten og fordelingen af muciner og slim in vivo.
I modsætning hertil indlejring fryse væv i Optimal Cutting Temperatur (OLT) medium enhulrum dehydrering og clearing af prøven og opretholder prøven hydrering. Dette giver mulighed for bedre bevarelse af hydratiseret mucuslaget, og således tillader studiet af de mange roller muciner i epitel biologi. Da denne fremgangsmåde kræver minimal forarbejdning af vævet er vævet opbevares i en mere naturlig tilstand. Derfor frosne væv sektioner kræver ikke yderligere forarbejdning før farvning og kan let analyseres ved hjælp af immunhistokemi metoder.
Vi viser bevarelsen af mikrometer tykt udskilte slim lag i frosne kolon prøver. Dette lag reduceres drastisk i de samme væv indlejret i paraffin. Vi viser også immunfluorescensfarvning af glycan epitoper præsenteret på muciner hjælp plante lectiner. Fordelen ved denne fremgangsmåde er, at den ikke kræver anvendelse af specielle fiksativer og tillader anvendelse af frosne væv, der allerede kan være konserveret i laboratoriet.
Protocol
1. Tissue Indlejring i OLT
- Der fremstilles en frysebadet ved tilsætning af tøris til 2-methylbutan i en overfladisk Styrofoam box.
- Høste væv og forsigtigt fugtigt på tissuepapiret til at tørre overskydende væske. Hvis der anvendes snap-frosset væv (væv, som blev nedfrosset i flydende nitrogen), tillade vævet at varme op til -20 ° C ved at placere det i en cryo-microtome kammer.
- Tilføje en lille mængde af OCT til Peel-A-Way frysning form, lige nok til at dække bunden af formen.
- Placere væv i formen, sørge vævet hviler på bunden af støbeformen på den ønskede orientering. Når frosset, vil vævsblok være sektioneret enten fra bunden eller fra siderne.
- Dæk væv med OLT, og placer formen i frysebadet. Den OCT-forbindelse vil blive hvid som vævet fryser.
- Når frosne, skræl formen fra den frosne blok og sted i en markant frysepose.
- De frosne blokke kan holdesved -80 ° C indtil anvendelse.
2. Vævssektionering
- Sted vævsblokke i cryo-mikrotom kammer, og tillade dem at nå -20 ° C (ca. 30 minutter).
- Skæres et 3-5 um tykke sektion og placere en positivt ladet objektglas oven på sektionen. Vævet vil klæbe til diaset.
- Air tørre væv i 30-60 min.
- Præparatglassene kan anvendes på dette trin, eller de kan opbevares ved -80 ° C til senere brug.
3. Vævsfarvning
- Objektglas, som blev opbevaret ved -80 ° C: giv objektglassene at tø og lufttørre ved stuetemperatur i 30 minutter.
- Fix objektglassene med 10% pufret formalin i 30 minutter ved stuetemperatur.
- Vaskes tre gange i PBS eller TBST puffer, for hver vask dyppe slides 10 gange i 250 ml puffer. PBS-buffer kan anvendes til immunfluorescensfarvning imidlertid for chromogen detektion med alkalisk phosphatase-conjugated antistof, bør TBST anvendes, da fosfat i PBS hæmmer alkalisk phosphatase-aktivitet.
- Vævssnit slides er nu klar til at blive farvet.
4. Brug Histokemiske Stains såsom Alcian Blue og periodiske Acid Schiff at Detect Slim
- Alcian Blue stain:
- Glassene skylles i vand, inkuberes i 3% eddikesyre i 3 minutter ved stuetemperatur.
- Farvet med Alcian Blue pH 2,5 opløsning i 30 minutter ved stuetemperatur.
- Vaske objektglassene i rindende ledningsvand i 10 minutter, skylles i DI-vand.
- Kontrastfarve i Nuclear Fast Red i 5 minutter ved stuetemperatur.
- Wash slides tre gange i destilleret vand.
- Periodisk Acid Schiff plet:
- Skyl dias i vand, inkuberes i frisk fremstillet 1% periodisk syre i 5 min.
- Vask tre gange i DI vand, dyppe en gang i miliQ vand.
- Stain med Schiff-reagens i 15 minutter ved rumtemperature.
- Vaske objektglassene i rindende ledningsvand i 10 minutter, skylles i DI-vand.
- Kontrastfarve i Surgipath Hematoxylin for 30 sekunder ved stuetemperatur.
- Wash slides tre gange i destilleret vand.
- Inkuberes slides 30 sek i Scotts ledningsvand ved stuetemperatur.
- Vask tre gange i destilleret vand.
- Dehydrere og fjerne objektglas ved at inkubere 1 min i 95% ethanol, efterfulgt af tre hurtige ændringer i 100% ethanol, og tre ændringer i Citrisolv, 2 min hver. Alle ved stuetemperatur.
- Monter lysbilleder på dækglas med harpiksholdige medium (Cytoseal 60).
5. Anvendelse af lectiner og antistoffer (tabel 1) for at påvise glycan epitoper ved histokemiske metoder
- Til fluorescenspåvisning af tre glycan epitoper under anvendelse lectiner, blokere objektglassene med 1% BSA i PBS i 10-30 minutter ved stuetemperatur.
- Da en biotinyleret lectin anvendes, blokere endogent biotin ved inkubering 15 minutter med 0,1%Avidin, efterfulgt af 15 minutters inkubering med 0,01% biotin ved stuetemperatur.
- Vask dias i PBS efter hvert bloktrinet.
- Frisk fremstille en blanding af 1 ug / ml rhodamin-konjugeret succinylerede hvedekimagglutinin (sWGA), 1,3 ug / ml biotinyleret Sambucus nigra agglutinin (SNA) og 5 pg / ml fluorescein-konjugeret Jacalin i HEPES / NaCl puffer (10 mM HEPES, 150 mM NaCI pH 7,5).
- Glassene anbringes på en flad overflade eller i et farvning boks, og laget lectinet blandingen på toppen. Blandingen volumen kan reduceres ved forsigtigt at placere parafilm på væsken, parafilmen flader væsken og forhindrer fordampning.
- Inkuber 1 time ved stuetemperatur i mørke.
- Wash slides tre gange med PBS.
- Lag objektglassene med 0,7 ug / ml streptavidin-konjugeret CY5 (til påvisning af biotinylerede-SNA), og der inkuberes i 30 minutter ved stuetemperatur i mørke.
- Wash slides tre gange med PBS.
- Kontrastfarve kerner med 0,1 & mu, g / ml DAPI.
- Montere lysbilleder på dækglas med vandigt medium, såsom VectaMount monteringsmedie (eller enhver vandige medier).
6. Styring for Lectin Staining Specificitet
- Lectin farvning specificitet styres enten ved specifik enzymatisk spaltning af target glycan epitop inden farvning eller ved konkurrence med små molekyler.
- Enzymatisk spaltning for Sambucus nigra agglutinin (SNA) bindingsspecificitet
- Fortynd Arthrobacter ureafaciens sialidase (AUS) til 250 mU / ml i 50 mM natriumacetat pH 5,5.
- Der tilsættes vand til bunden af en tom tip-box, vil dette danne et fugtigt kammer under inkubation.
- Anbring objektglassene med billedsiden op på den øverste bakke på tip-box, lag 150-200 pi AUS løsning på objektglasset og dækkes med et dækglas. Undgå luft bobledannelse.
- Luk æskens låg og inkuberes ved 37 ° C i 2,5 timer.
- Wash slides tre gange i PBS ved rumtemperaratur at fjerne alle frie sialsyrer. Disse slides bør være negativt for SNA farvning.
- Kompetitive inhibitorer for Jacalin og succinyleret hvedekimagglutinin (sWGA) specificitet
- Aliquot 200 ul af lectinet blanding fremstillet i trin 5.4 til to Eppendorf hætteglas.
- Tilføj 200 mM melibiose (Jacalin inhibitor) til et af hætteglassene og Chitin-hydrolysatet ved 1:10 fortynding (sWGA inhibitor) til den anden hætteglas.
- Overlay negative kontrolglas med inhibitorer indeholdende blandingen og inkuberes 1 time ved stuetemperatur på samme tidspunkt som resten af gliderne. Disse slides bør være negativ for Jacalin eller sWGA farvning hhv.
7. Repræsentative resultater
En sammenligning mellem vævsprøver indlejret i paraffin til frosne væv indlejret i cryo-beskyttelsesmiddel medier (OLT) viste slående forskel i bevarelsen og kvaliteten af farvning for mucin glycoproteiner. Vævsfarvning med histokemiske farvestoffer, såsom Alcian Blue og periodiske Acid Schiff, producerer meget forskellige resultater i sammenlignelige vævssnit fra frosne eller paraffin indlejret prøver (figur 1). Det fremgår, at det organiske opløsningsmiddel (xylen eller Citrisolv), der bruges i paraffin embedding proces påvirker fordelingen af secernerede muciner på epiteler og fjerne meget af glycolipider fra prøverne (figur 2). Som følge heraf forekommer slimlaget sammen på slimhinden celler og findes hovedsageligt i slimceller. Flash frysning af væv i cryo-beskyttelsesmiddel medier (OLT) vedligeholdt prøve hydrering og bevarede de secernerede muciner lag dimensioner. The paraffin embedding proces påvirkes andre mucus-associerede glycaner og glycolipider på samme måde. Glycan fordeling blev undersøgt under anvendelse af lectiner, der rutinemæssigt anvendes til glycan detektering (fig. 3) og antistoffer mod epitoper der findes påmuciner og glycolipider (fig. 6). Fordi lectin binding ikke er veldefineret, og er påvirket af den rumlige fordeling af glycaner samt glycan struktur 14,15, er det vigtigt at anvende de rette metoder til lectin farvning. Her viser vi to metoder til styring lectin farvning på de testede væv: enzymatisk spaltning og kompetitiv hæmning. Spaltning af de glycan epitoper blev gjort ved fordøjelse af vævssnittet med glycan-specifikke enzymer, f.eks bakteriel sialidase som kontrol for sialinsyre binding af SNA (figur 4). I tilfælde, hvor specifikke enzym (f.eks glykosidase) er ikke tilgængelig for fjernelse af glycan-epitopen undersøgt, kan lectin specificitet bekræftes ved tilsætning af en kompetitiv inhibitor, såsom melibiose for Jacalin farvning eller Chitin-hydrolysatet for sWGA farvning (fig. 5).
Vi viser her, at snap-frosne vævsprøver, som er routinely opnået i klinikken og i forskningslaboratorier, kan yderligere indlejret i OCT og anvendes til at undersøge mucin-glycoproteiner og de mange glycans stede på dem.
| Lectin / Ab | Kilde | Major specificitet |
| LFA | Limax flavus (Gul slug) | Terminal Sia |
| MAA * | Maackia amurensis (Amur Maackia) | Siaα2-3Galβ1-R / 3-O-sulfat på Galβ1-R |
| SNA | Sambucus nigra (Elderberry) | Siaα2-6Gal / Siaα2-6GalNAc |
| WGA | Triticum vulgaris (Hvedekim) | GlcNAcβ1-4GlcNAcβ1-4GlcNAc / Sia |
| sWGA | Succinyleret Triticum vulgaris (Hvedekim) | GlcNAc & beta; 1-4GlcNAcβ1-4GlcNAc |
| PNA | Arachis hypogaea (peanut) | Galβ1-3GalNAc (umodificeret T-antigen) |
| Jacalin | Artocarpus integrifolia (Jacalin) | Galβ1-3GalNAc fundet på O-bundne glycans |
| ECA | Erythrina cristagalli (Coral træ) | Galβ1-4GlcNAc |
| TKH2 | Antistof | Siaa2-6GalNAc (STN) på O-bundne glycans |
| CA19-9 | Antistof | Siaa2-3Galβ1-4 (Fuca1-3) GlcNAc (SLE a) |
| SNH3 | Antistof | Siaa2-3Galβ1-3 (Fuca1-4) GlcNAc (SLE x) |
Forkortelser: Ab, antistof, Sia, Sialinsyre, Gal, Galactose; GaINAc,
Tabel 1. En delvis liste over lectiner og antistoffer til glycan epitoper.
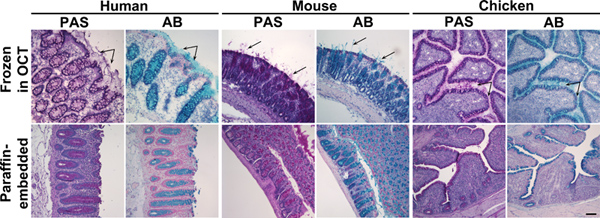
Figur 1. Alcian Blue og periodisk syre Schiff farvning af frosne og paraffinindlejret kolon væv. Vævssnit af human, mus eller kylling kolon prøver frosset i OCT (øverste panel) eller indlejret i paraffin (nederste panel) blev farvet med periodisk syre Schiff (PAS) eller Alcian Blue (AB). Disse reagenser plette slim pink eller blå, hhv. Upper panel: I frosne væv, ud over muciner i slimceller var secerneret slim også overenelig (pile). Nederste panel: I paraffinindlejrede væv, blev slim farvning begrænset til slimceller. Kerner blev modfarvet med Surgipath (PAS) eller Mayers (AB) hematoxylin. Scale bjælke angiver 100 um. Klik her for at se større figur .

Figur 2. Citrisolv inkubation resulterer i væsentlig tab af muciner. Seriesnit af frosne kylling ileum prøver blev fikseret i 10% pufret formalin og holdt hydratiseret (venstre panel), dehydreret i ethanol ved sekventiel inkubering i 70%, 90% og 100% ethanol i 20 minutter hver (midterste panel), eller dehydreret i ethanol og godkendt med Citrisolv i 1 time (højre panel). Tørrede prøver blev rehydreret tilbage til PBS før hematoxylin og eosin (H & E) eller Alcian Blue farvning. Ethanol dehydrering og Citrisolv clearing forbedret væv morphology (f.eks øverste række, midterste og højre billeder versus venstre billede). Ethanol dehydrering havde ingen signifikant effekt på Alcian blå farvning (midterste række, sammenligne venstre og midterste billeder). I modsætning hertil reducerede Citrisolv inkubation Alcian Blue farvning, og kun den til slimceller (midterste række, højre billede) i et mønster, der svarede til den, set i paraffinindlejrede væv (figur 1). Disse data indebærer, at den kraftige farvning af slim granula i paraffinindlejrede prøver skyldes krympning og kondensering af slim i slimceller under clearing trin. Den svagere og mindre tæt farvning af slim i frosne oktober-indlejrede væv afspejler en mere naturlig fordeling af slim i vævet. Højere forstørrelser af boxed områder er markeret med pile. Skala søjler angiver 50 pm (øverste og midterste rækker) og 10 um (nederste række).

Figur 3. Binding aflektiner til glycan epitoper på frosne og paraffinindlejret væv. Chicken tyndtarmen (ileum) prøver frosset i OCT (øverste paneler) eller indlejret i paraffin (nedre paneler) blev undersøgt med Jacalin (blå), sWGA (grøn) og SNA (rød ). Højere forstørrelser af boxed områder er markeret med pile. I frosne væv Jacalin binding til O-bundne glycaner afslørede strukturer, der syntes at være udsivning fra slimceller ind i lumen (Jacalin, øvre panel, pile). I modsætning hertil blev Jacalin binding til paraffinindlejrede væv begrænset til slimceller (Jacalin, nederste panel, pile) og til villi brush border (nederste venstre billede, pil). sWGA farvning af β1-4GlcNAc delvist co-lokaliseret med bindingen af Jacalin lektin i begge frosne væv (øverste panel, pile), og paraffinindlejret væv (nedre panel, pile). I modsætning hertil er SNA lectin binding til α2-6 forbundne sialinsyrer intracellulært (SNA, pilehoveder), og som ikke co-lokalisere med Jacalin (farvebillede, SNA i rød markeret med pilespids, Jacalin i blåt markeret med stiplede pil). Scale søjler angiver 100 um (venstre billeder) og 20 um (forstørrede boxed områder).

Figur 4. Enzymatisk spaltning kontrol for sialinsyre farvning med SNA. Chicken tyndtarmen prøver blev inkuberet med 250 mU / ml Arthrobacter ureafaciens sialidase (AUS) eller med 50 mM natriumacetat pH 5,5 puffer i 2,5 timer ved 37 ° C. AUS behandling ophæver farvning med biotinyleret SNA, bekræfter SNA bindingsspecificitet til sialsyrer. Scale bjælke angiver 100 um.

Figur 5. Kompetitiv inhibering kontrol for glycan farvning med Jacalin og sWGA. Chicken tyndtarmen (ileum) prøver blev inkuberet med en Jacalin og sWGA blandingen i nærvær af specifikke LECtin inhibitorer: melibiose (midterste søjle), Chitin-hydrolysat (højre kolonne) eller uden inhibitor (A og D). Øverste panel: (venstre) Jacalin farvning uden inhibitorer. (I midten) Jacalin farvning blev hæmmet af melibiose. (Højre) Chitin-hydrolysat inhiberede ikke Jacalin farvning. Nederste panel: (venstre) sWGA farvning uden inhibitor. (I midten) melibiose hæmmede ikke sWGA farvning. (Højre) sWGA farvning blev inhiberet af Chitin-hydrolysatet. Denne inhibering bekræfter den specifikke interaktion af lectiner med vævene. Asterisker mark billede af farvning inhibering. Scale bjælke angiver 100 um.

Figur 6. Påvisning af sekretoriske muciner, glycolipider og glycan epitoper i frosne humane kolorektal cancer væv. Kolorektal cancer biopsier fra villous carcinom og slim carcinoma var lynfrosset i flydende nitrogen og indlejret i OCT. Vævssnit blev inkuberet i 1 timemed antistoffer mod det secernerede mucin MUC5AC, sialyl-Lewis a - glycan epitop fundet på gangliosider (kolorektal cancer markør CA 19-9), og sialyl-Tn - rigelige glycan epitop på muciner (detekteret med TKH2 antistof), efterfulgt af 30 minutters inkubation med biotinyleret æsel-anti-muse-IgG sekundært antistof, og 30 minutters inkubation med peroxidase-konjugeret streptavidin. Yderligere væv blev inkuberet i 1 time med det biotinylerede lectiner SNA og sWGA, efterfulgt af 30 minutters inkubation med peroxidase-konjugeret streptavidin. Peroxidase-farvning blev udviklet ved anvendelse AEC-kit. Sort skala bjælke angiver 200 um.
Discussion
Bevarelse af slim og glycan epitoper i frosne væv er overlegen i forhold til væv, som blev indlejret i paraffin. Vi viste bevarelse af secerneret mucuslaget (figur 1 og 3), og fordelingen af tre glycans strukturer (fig. 3) i frosne væv sammenlignet med paraffinindlejrede væv. Specialiserede fiksativer, såsom Carnoy opløsning (60% ethanol, 30% chloroform, 10% eddikesyre) 17 er blevet udviklet til optimal bevaring af slimlaget i vævsprøver. Optimalt set bør denne opløsning anvendes til at opsamle vævsprøver, der er dedikeret til slim undersøgelser og blev vist at opretholde et effektivt udseende af mucuslaget 16-17. Det slim lag i ufikserede frosne prøver indlejret i OCT forekommer robust og i nogle områder kan løsnes fra vævet, men den samlede lagtykkelse er indforstået med, der blev observeret i væv, der blev fastsat med Carnoy opløsning og embedded i paraffin 16-17. For eksempel er slimlaget i frosset human colon vævssnit ~ 100 um (fig. 1), hvilket er inden for det område rapporteret for Carnoy's-fikseret human colon prøve 55,4 ± 2,5 um (intervallet fra 7,7 til 204,8 um) 16.
Det har været kendt i årtier, at ethanol dehydrering resulterer i ~ 30% krympning af biologiske prøver 18, og at organiske opløsningsmidler, såsom xylen, Citrisolv og chloroformekstrakten lipider, glycolipider og i en vis udstrækning, proteiner fra vævene 13. Vævsbehandling for paraffin indlejring omfatter følgende trin: fiksering (10% pufret formalin), dehydrering (stigende ethanolkoncentration) og clearing (Citrisolv eller xylen). Ved at efterligne disse trin på ikke-fikserede frosne vævssektioner demonstrerede vi, at Citrisolv udtrækker slim fra frosne vævssektioner resulterer i vævsmorfologi, der ligner den af paraffinindlejrede væv (Figure 2, højre panel). I modsætning hertil blev slimlaget ikke ændret ved inkubation med formalin eller ethanol (figur 2, venstre og midterste paneler). Dette antyder, at clearing trinnet standard paraffin embedding procedure, som kræver forlænget inkubering i Citrisolv / xylen, resulterer i sammenbrud af mucuslaget. Formalinfiksering skader ikke slim lag og frosne væv sektioner, som blev fikseret med formalin kan let farvet med lectiner og antistoffer mod glycans, glycolipider og proteiner (figur 2, 3 og 6). Disse virkninger kan være ubetydelige til undersøgelse af membranbundne proteiner og vævs patologi, men de er ødelæggende for stærkt hydratiserede strukturer som det secernerede slimlag. Imidlertid histologiske undersøgelser af muciner stadig udføres mest med paraffinindlejrede prøver, hvor slimlaget konservering er suboptimal. Dybdegående analyser af mucuslaget sammensætning, såsom den nøjagtige identitet af secreted eller membranbundne MUC glycoprotein kombinationer kræver specifikke antistoffer og massespektrometri til identifikation af protein backbone-net. Konservering af slim lag er, men den indledende krav til sådanne undersøgelser.
Mange laboratorier har vævsprøver indefrosset i OCT, der blev indsamlet i fortiden til forskellige projekter, kan disse væv let bruges til at undersøge muciner, glycolipider og glycan fordeling eliminerer behovet for at indsamle væv i særlige fiksativer, der er designet unikt for slim konservering. Frosne væv undergår minimal forarbejdning og derfor den naturlige fordeling af glycaner, som er hydratiseret karakter, bevares. Dette er især vigtigt i realm af mikrobielle vært interaktioner. Viden om naturalistiske udbredelse og talrighed secernerede muciner og de mange glycan strukturer udsmykning disse "barriere" molekyler vil være afgørende i forståelsen vært forsvar, mikrobiel udnyttelse og patogenerer.
Disclosures
Ingen interessekonflikter erklæret.
Acknowledgments
Forfatterne vil gerne takke Nicole M. Nemeth (University of Georgia) og Jeanne M. Fair (LANL) for deres hjælp i høst kylling væv, og Steven A. Springer for hans hjælp under optagelserne. Pasning af alle fugle i denne undersøgelse var i overensstemmelse med National Institutes of Health retningslinjer for human brug af forsøgsdyr, og alle protokoller blev godkendt af Institutional Animal Care og brug udvalgene på Los Alamos National Security, LLC, operatøren af Los Alamos National Laboratory under kontrakt nr. DE-AC52-06NA25396 med det amerikanske Department of Energy. Pasning af mus i denne undersøgelse er i overensstemmelse med UCSD dyr godkendte protokol. Humane væv blev opnået som en del af UCSD godkendt IRB protokol. Dette arbejde blev støttet af tilskud 118.645 fra University of California Lab Fee President Program (PG) og tilskud NS047101 fra National Institute of Neurologiske og Stroke (Neuroscience Microscopy Delt Facility, UC San Diego).
Materials
| Name | Company | Catalog Number | Comments |
| 2-methyl butane | Fisher Scientific | 03551-4 | |
| AEC peroxidase substrate kit | Vector Labs | SK-4200 | |
| Alcian Blue | Sigma-Aldrich | A3157 | |
| Anti-CA 19-9 monoclonal antibody | Calbiochem | CA1003 | |
| Anti-MUC5AC monoclonal antibody | Millipore | MAB2011 | |
| Avidin-Biotin blocking kit | Vector Labs | SP-2001 | |
| Biotinylated donkey anti-mouse antibody | Jackson Immunoresearch | 90863 | |
| Biotinylated SNA | Vector Labs | B-1305 | |
| Bovine serum albumin | Sigma-Aldrich | A4503 | |
| Chitin-hydrolysate | Vector Labs | SP-0090 | |
| Cryostat microtome | Leica Microsystems | Leica CM 1800 | |
| Hematoxylin | Surgipath Medical Ind. | 3801570 | |
| Hydrogen peroxide 30% | Fisher Scientific | H325-100 | |
| Jacalin-FITC | Vector Labs | FL-1151 | |
| Mayer's Hematoxylin | Sigma-Aldrich | MHS32 | |
| Melibiose | Sigma-Aldrich | M5500 | |
| Nuclear Fast Red | Vector Labs | H-3403 | |
| OCT compound | VWR International | 25608-930 | |
| Peroxidase conjugated streptavidin | Jackson Immunoresearch | 94638 | |
| Schiff reagent | Electron microscopy sciences | 26052 | |
| sWGA-Rhodamine | Vector Labs | RL1022S | |
| TKH2 monoclonal antibody | ATCC | HB-9654 | |
| VectaMount aqueous mounting media | Vector Labs | H-5501 | |
| Cytoseal 60 | Thermo Scientific | 8310-4 | |
| Peel-A Way molds | Polysciences Inc. | 18646A-1 |
References
- Slayter, H. S., Wold, J. K., Midtvedt, T. Intestinal mucin of germ-free rats. Biochemical and electron-microscopic characterization. Carbohydr. Res. 222, 1-9 (1991).
- Lamblin, G. The carbohydrate diversity of human respiratory mucins: a protection of the underlying mucosa. Am. Rev. Respir. Dis. 144, S19-S24 (1991).
- Corfield, A. P., Carroll, D., Myerscough, N., Probert, C. S. Mucins in the gastrointestinal tract in health and disease. Front. Biosci. 6, D1321-D1357 (2001).
- Turner, J. R. Intestinal mucosal barrier function in health and disease. Nat. Rev. Immunol. 9, 799-809 (2009).
- Lievin-Le Moal, V., Servin, A. L. The front line of enteric host defense against unwelcome intrusion of harmful microorganisms: mucins, antimicrobial peptides, and microbiota. Clin. Microbiol. Rev. 19, 315-337 (2006).
- Kim, Y. S., Ho, S. B. Intestinal goblet cells and mucins in health and disease: recent insights and progress. Curr. Gastroenterol. Rep. 12, 319-330 (2010).
- Nochi, T., Kiyono, H. Innate immunity in the mucosal immune system. Curr. Pharm. Des. 12, 4203-4213 (2006).
- Vareille, M., Kieninger, E., Edwards, M. R., Regamey, N. The airway epithelium: soldier in the fight against respiratory viruses. Clin. Microbiol. Rev. 24, 210-229 (2011).
- McGuckin, M. A., Linden, S. K., Sutton, P., Florin, T. H.
- Knowles, M. R., Boucher, R. C. Mucus clearance as a primary innate defense mechanism for mammalian airways. J. Clin. Invest. 109, 571-577 (2002).
- Johansson, M. E. The inner of the two Muc2 mucin-dependent mucus layers in colon is devoid of bacteria. Proc. Natl. Acad. Sci. U.S.A. 105, 15064-15069 (2008).
- Hooper, L. V., Macpherson, A. J. Immune adaptations that maintain homeostasis with the intestinal microbiota. Nat. Rev. Immunol. 10, 159-169 (2010).
- Hayat, M. A. Microscopy, Immunohistochemistry, and Antigen Retrieval Methods: For Light and Electron Microscopy. Hayat, M. A. , Kluwer Academic/Plenum Publishers. New York, NY. 53-70 (2002).
- Cohen, M., Hurtado-Ziola, N., Varki, A. ABO blood group glycans modulate sialic acid recognition on erythrocytes. Blood. 114, 3668-3676 (2009).
- Cohen, M., Varki, A. The sialome--far more than the sum of its parts. OMICS. 14, 455-464 (2010).
- Ota, H., Katsuyama, T. Alternating laminated array of two types of mucin in the human gastric surface mucous later. Histochemical J. 24, 86-92 (1992).
- Matsuo, K., Ota, H., Akamatsu, T., Sugiyama, A., Katsuyama, T. Histochemistry of the surface mucous gel layer of the human colon. Gut. 40, 782-789 (1997).
- Boyde, A., Maconnachie, E. Treatment with lithium salts reduces ethanol dehydration shrinkage of glutaraldehyde fixed tissue. Histochemistry. 66, 181-187 (1980).
