

Summary
Bijhouden kleine veranderingen in de progressie en kinetiek van celcyclus stadia worden door gebruik van een combinatie van metabolische labeling van nucleinezuren met BrdU en totaal genomisch DNA kleuring via propidium jodide. Deze methode wordt voorkomen dat van de chemische synchronisatie van fietsen cellen, waarbij de introductie van niet-specifieke DNA-schade die op zijn beurt invloed celcyclus.
Abstract
Nauwkeurige controle van de inleiding, het doorlopen van de verschillende fasen van de celcyclus zijn van het grootste belang in prolifererende cellen. Celcyclus divisie is een integraal onderdeel van de groei en voortplanting en deregulering van de belangrijkste celcyclus componenten zijn betrokken bij de precipiterende gebeurtenissen van carcinogenese 1,2. Moleculaire agenten in anti-kanker therapieën vaak richten op biologische routes die verantwoordelijk zijn voor de regulering en de coördinatie van de celcyclus divisie 3. Hoewel celcycluskinetiek algemeen afhankelijk van celtype is de verdeling van cellen tussen de vier stadia van de celcyclus is vrij consistent binnen een cellijn te wijten aan de patroon mitogeen en groeifactor expressie. Genotoxische gebeurtenissen en andere cellulaire stressfactoren kan een tijdelijke blok celcyclusprogressie, waardoor arrestatie of een tijdelijke onderbreking in een bepaalde fase van de celcyclus mogelijk te maken instellingenplichting van de juiste reactie mechanisme.
De mogelijkheid om experimenteel observeren het gedrag van een celpopulatie met betrekking tot hun voortgang van de celcyclus fase is een belangrijke stap vooruit in de celbiologie. Gemeenschappelijke procedures zoals mitotische af te schudden, differentieel centrifugeren of flowcytometrie op basis van sorteren worden gebruikt om cellen te isoleren in specifieke fasen van de celcyclus 4-6. Deze gefractioneerd fase van de celcyclus verrijkte populaties vervolgens onderworpen aan een experimentele behandeling. Opbrengst, zuiverheid en levensvatbaarheid van de gescheiden fracties kunnen vaak in gevaar worden gebracht met behulp van deze fysieke scheiding methoden. Eveneens, kan de tijd die verstrijkt tussen de scheiding van de cel bevolking en de start van de experimentele behandeling, waarbij de gefractioneerde cellen kunnen evolueren van de geselecteerde cel cyclus fase, vormen belangrijke uitdagingen in de succesvolle implementatie en interpretatie van deze experimenten.
Andere benaderingen van stUdy celcyclus fasen omvatten het gebruik van chemicaliën om cellen te synchroniseren. Behandeling van cellen met chemische remmers belangrijke metabolische processen voor elke celcyclus fase nuttig bij het blokkeren van de progressie van de celcyclus de volgende fase. Bijvoorbeeld, de ribonucleotidereductase remmer hydroxyurea stopt cellen in de G1 / S moment door het beperken van deoxynucleotiden, de bouwstenen van DNA. Andere belangrijke chemicaliën omvatten behandeling met aphidicolin, een polymerase alfa-inhibitor voor G1 arrestatie behandeling met colchicine en nocodazole, beide verstoren mitotische spindel vorming stoppen cellen M en tot slot de behandeling met de DNA ketenterminator 5 fluorodeoxyridine te leiden S-fase arrestatie 7-9. Behandeling met deze stoffen is een doeltreffend middel synchroniseren een gehele populatie van cellen in een bepaalde fase. Met het verwijderen van de chemische, cellen opnieuw de celcyclus in koor. Behandeling van het testmiddel volgende releasevan de celcyclus blokkeren chemische zodat het geneesmiddel zou opleveren is van een uniform celcyclus stadium-specifieke populatie. Echter, omdat veel van de chemische synchronizers bekende genotoxische stoffen, plagen elkaar de deelname van de verschillende bestrijdingsstrategieën wegen (op de synchronizers vs de test agenten) is een uitdaging.
Hier beschrijven we een metabolische labeling methode voor het volgen van een subpopulatie van actief delende cellen door middel van hun progressie van de DNA-replicatie fase, tot en met de verdeling en de scheiding van hun dochter cellen. In combinatie met flowcytometrie kwantificering, dit protocol maakt voor het meten van kinetische progressie van de celcyclus in de afwezigheid van een mechanisch of chemisch-geïnduceerde cellulair benadrukt vaak geassocieerd met andere celcyclus synchronisatie methodologieën 10. In de volgende paragrafen bespreken we de methodologie, evenals een aantal van haar toepassingen in biomedisch onderzoek.
Protocol
1. Celpreparaat

- Plaat cellen een dichtheid van ongeveer 60% confluentie bereiken. De cellen moeten log fase bij het verzamelen. Voor MCF7 cellen wordt dit bereikt door enten van 5 x 10 5 cellen / 10 cm plaat in de geschikte media. HT29 en LS180 cellen worden uitgezaaid in 6 x 10 5 cellen / 6 cm plaat. We gebruikten DMEM medium aangevuld met 10% FBS en 1x penicilline / streptomycine. Zorg ervoor dat zaad platen voor de juiste positieve en negatieve controles inNaast uw test monsters. Zij omvatten het volgende:
| ik | positieve controle | BrdU alleen |
| ii | positieve controle | PI alleen |
| iii | negatieve controle | BrdU negatief, PI negatief |
(Er wordt opgemerkt dat een eerste experiment met meerdere tijdstippen zoals die beschreven in dit protocol kan worden gebruikt om een beperking van de tijd gedurende welke uitvoeren toekomstige verzamelingen.)
- De plated cellen worden geïncubeerd bij 37 ° C, 5% CO2 gedurende 24-48 uur, waardoor cellen te herstellen en bevestigen.
- Om label cellen pulseren met broomdeoxyuridine (BrdU), wordt DMEM vervangen with vers medium met 10 pM BrdU. Cellen worden gedurende 1 uur bij 37 ° C, 5% CO2 mogelijk te maken BrdU opname in het DNA. Zorg ervoor dat u een plaat onbehandeld op te treden als de negatieve controle verlaten.
- De puls-etikettering media wordt verwijderd en de cellen worden in het kort gespoeld met 1X PBS.
- Verse groeimedia toegevoegd (minus de BrdU) en cellen kunnen blijven incuberen bij 37 ° C, 5% CO2 totdat de gewenste meetpunt voor de oogst is bereikt.
2. Oogst en fixatie
- Onbehandelde cellen uit stap 1.3 worden beschouwd als de nul tijdpunt. Dit monster kan worden geoogst, samen met de 1 uur tijdpunt die onmiddellijk wordt verzameld na BrdU behandeling.
- Om de cellen te oogsten, wordt media verwijderd en de platen worden eenmaal gespoeld met 1X PBS.
- Cellen worden getrypsiniseerd, verzameld in kweekmedia en vervolgens gepelleteerd door centrifugeren bij 1500 rpm gedurende 5 minuten Supernatant wordt weggegooid.
- Spoel 1-10 x 10 6 cellen in 5 ml ijskoud 1X PBS. Centrifugeer bij 1500 rpm gedurende 5 minuten supernatans verwijderd.
- De celpellet wordt geresuspendeerd in 100 ul ijskoud PBS / 1% FBS. Toevoeging van 1% FBS in PBS steun voorkomen cel samenklontering.
- Voeg deze cellen druppelsgewijs 5 ml van -20 ° C, 70% ethanol om de cellen te lossen.
- Incubeer op ijs gedurende 30 minuten of op te slaan bij 4 ° C gedurende de nacht. Dit is een ideale stap voor stap om het verzamelen van monsters te stoppen uit de verschillende tijdstippen. De monsters kunnen worden gelaten in ethanol enkele dagen, waardoor ze tegelijkertijd worden verwerkt door de rest van het protocol.
3. BrdU en PI kleuring
- Pellet cellen door centrifugeren gedurende 5 min bij 1500 rpm.
- Verwijder het fixatief, maar laat ~ 50 ul in om de pellet los te maken door vortexen.
- Om de DNA denatureren langzaam 1 ml 2N HCl / Triton X-100druppelsgewijs terwijl vortexen. Incubeer monsters bij kamertemperatuur gedurende 30 minuten.
- Pellet cellen door centrifugeren monsters 5 min bij 1500 rpm. Zuig Verwijder de bovenstaande vloeistof.
- Resuspendeer de celpellet in 1 ml 0,1 M natriumtetraboraat, pH 8,5, de denaturatiestap neutraliseren.
- Centrifugeer cellen gedurende 5 min bij 1500 rpm. Zuig Verwijder de bovenstaande vloeistof.
- Resuspendeer de celpellet in 75 pi BrdU kleuring mix (50 pi 0,5% Tween 20/1% BSA / PBS + 20 pi FITC-geconjugeerd anti-BrdU + 5 pi 10 mg / ml RNase).
- Incuberen bij kamertemperatuur gedurende 45 min van licht.
- Om pellet cellen gecentrifugeerd monsters 5 min bij 1500 rpm. Zuig Verwijder de bovenstaande vloeistof.
- Resuspendeer de celpellet in 1 ml PBS met 5 pg / ml propidiumjodide.
4. Flowcytometrie
- Om de cellen te analyseren, een flowcytometer met een 488 nm laser enpassende filters nodig. Analyse software zoals CellQuest nodig is om de kavels hieronder beschreven maken.
- Bij het uitvoeren van het monster door de flowcytometer, maak een voorwaartse verstrooiing (SSC-H) vs zijwaartse verstrooiing (FSC-H) complot om de juiste grootte verdeling van de cellen (figuur 1A) te verzekeren. Beide parameters zijn uitgezet op een lineaire schaal.
- Tegelijkertijd bekijken van een perceel van FL3-H (PI kleuring) vs FL2-W (Figuur 1B). Deze plot wordt gebruikt om een poort (R3) van de fractie van cellen die in de normale verdeling patroon van de celcyclus isoleren maken. In het algemeen is deze verdeling kenmerkt door twee clusters van cellen van de 2N en 4N DNA inhoud van PI kleuring die de G1 en G2 fasen van de celcyclus respectievelijk. Een reeks van cellen zich tussen deze twee clusters representatief is lopende DNA replicatie die optreedt in de S-fase van de celcyclus.
- Maak een plot displaying van de gated cellen (R3) in stap 4.3, met FL1-H (FITC log plot) op de y-as en PI (lineaire grafiek) op de x-as (figuur 1c). Dit perceel wordt gebruikt om de overige parameters met de verschillende bedieningsorganen beschreven in 1.1 en de gegevens voor analyse verzamelen stellen.
- Plaats de PI enige gebrandschilderde monster op de cytometer. Stel de winst voor G1 plaatsen op ~ 200 op de x-as. Dit zal het gemakkelijker zijn om te visualiseren in een histogram plot van de PI vlek.
- Een tweede controle omvat het uitvoeren van een BrdU-only monster op de flowcytometer. Stel de winst, zodat de twee populaties van cellen (BrdU positieve versus negatieve BrdU) verschijnen op het perceel. Idealiter wordt de BrdU negatieve cellen gepositioneerd weergegeven net onder 10 -1.
- De laatste controle is de BrdU / PI negatief monster. Voer deze negatieve controle opdat cellen niet in een van de bovenste of rechts kwadranten.
- Zodra de parameters van de verschillende percelen zijn ingesteld, deflowcytometer is gekalibreerd en klaar voor de verwerking van BrdU en PI gekleurde cellen. Een minimaal 10.000 cellen die gated in de juiste PI fractie (zie 4,3 stap) worden gelezen per verzameling.
- Om de verzamelde gegevens te analyseren, worden software zoals FlowJo of FacsDiva benut. Elk van de bovengenoemde percelen kan worden gecreëerd in deze programma's en kwantitatieve statistische analyse uitgevoerd.
- Het eindpunt van de analyse omvat verschillende opeenvolgende stappen. Het creëren van een perceel van FITC vs PI met cellen gated als PI positief uit de PI vs FL2-W perceel maakt het mogelijk om onderscheid te maken het fietsen bevolking. Een tweede poort is gemaakt op basis van dit perceel door het isoleren van de BrdU positieve populatie. Het uiten van deze bijzondere populatie op een histogram perceel met PI op de x-as maakt het mogelijk om bij te houden het tijdsverloop van de voortgang van de celcyclus. Dit kan verder zichtbaar getrokken waarbij het aantal BrdU-positieve cellen met G1 en G2 inhoud als functie van de tijd.
5. Representatieve resultaten
Normaal delende cellen gekleurd met propidium jodide hebben verschillende pieken bij G1 en G2, overeenkomend met cellen die 2N en 4N DNA inhoud respectievelijk (figuur 2). Pulse labeling met BrdU zorgt voor een selectieve etikettering van een subpopulatie van cellen die actief zijn synthese van DNA (dat wil zeggen de S-fase). Kort nadat de BrdU reagens, alle gelabelde cellen in S phase (figuur 3). Door beperken etikettering een korte puls men kan dit nu verschillende subpopulatie van cellen gevolgd gedurende vele tijdstippen als ze door de volgende fasen van de celcyclus. Dit kan worden gevisualiseerd op het 1 uur tijdstip in figuur 3 als een afzonderlijke weinig G1 en G2 pieken.
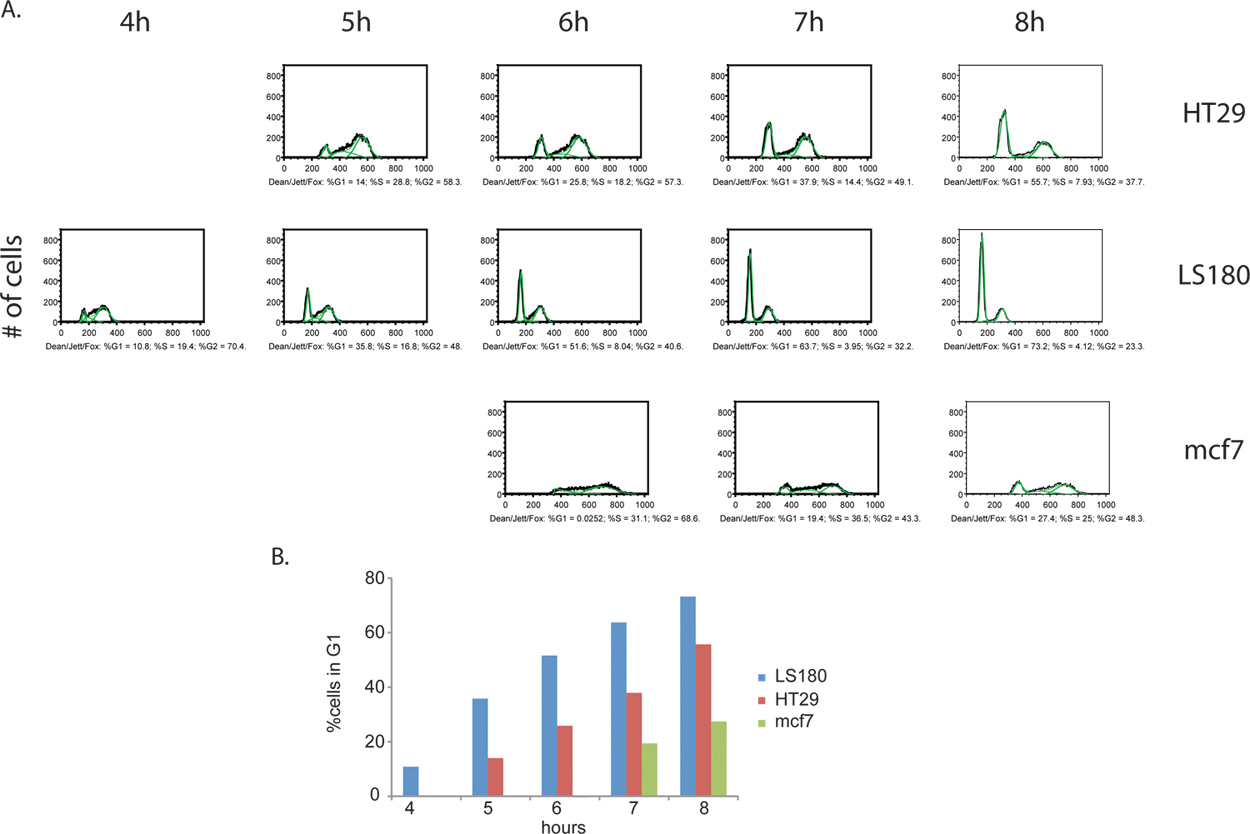
Aanvullende informatie kan worden afgeleid uit de BrdU etiket stap. Niet alleen kan de hoogte van actief delende cellen worden, maar een estiMate van celcyclus stadiumverdeling tussen twee monsters kunnen ook worden vastgesteld. Door het verzamelen van cellen op gelijke afstand van elkaar intervallen na het verwijderen van de BrdU reagens, kunnen cellen worden opgespoord als ze blijven om te fietsen, vooruitgang G2 en uiteindelijk gaan door naar de celdeling van de originele cellen, uiteindelijk naar voren als BrdU-positieve dochter cellen met G1 DNA-gehalte (figuur 4). Een voorbeeld van de celcyclus fase kwantificering en kinetische analyse wordt in figuur 5.

Figuur 1. (A) Forward Scatter versus Side Scatter Plot van een vertegenwoordiger MCF7 celpopulatie. (B) PI versus Breedte (FL-W) plot van een vertegenwoordiger MCF7 celpopulatie. Gating is aangetoond dat cellen wambuizen uit te sluiten in de uiteindelijke analyse (R3). Cell doubletten zullen grotere pulsbreedte dan een enkele cel, zoals het langer duurt voordat door het laserstraal en thereferts kunnen worden uitgesloten van de analyse. (C) PI versus FITC (BrdU) plot van een vertegenwoordiger MCF7 celpopulatie. Gating wordt alleen de FITC (BrdU) positieve cellen (R4) omvatten.

Figuur 2. Vertegenwoordiger histogram plot van een totale bevolking van normaal delende cellen.

Figuur 3. Histogram plot van cellen die zijn 1h verzameld na verwijdering van de BrdU puls na gating voor FITC (BrdU)-positieve cellen. BrdU-positieve cellen in een vroeg tijdstip bepaald na verwijdering van het etiket tonen PI profielen die overeenkomen met monsters getoond DNA inhoud overeenstemming met cellen die in de S-fase van de celcyclus, hetgeen de succesvolle labeling van de cellen alleen tijdens DNA synthese.
 lt = "Figuur 4" />
lt = "Figuur 4" />
Figuur 4. Vergelijking van de celcyclus kinetiek tussen cellijnen, colorectale kanker HT29 (A) en LS180 (B), alsmede borstkanker MCF7 (C). Cellen werden verzameld elk uur 8 uur na de verwijdering van de BrdU puls. In dit experiment, zagen we een duidelijk profiel van een versnelde voortgang van de celcyclus door de G2 fase van de celcyclus in de colorectale cellijn LS180. Vergelijking van de kinetica van de profielen van colorectale kanker cellen, het ontstaan van een G1 piek is duidelijk op T = 4h na BrdU de LS180 cellen, vergeleken met de overeenkomstige tijdstip voor de HT-29 cellijn die deze piek vertoont. In vergelijking met een van de twee colorectale cellijnen, worden MCF7 cellen fietsen tegen een aanzienlijk lagere prijs. Klik hier om een grotere afbeelding te bekijken .
Ad 5 "/>
Figuur 5. (A) Quantitative celcyclus fase analyse BrdU gelabelde kankercellen. De decaan / Jett / Fox-algoritme werd toegepast op HT29, LS180 en MCF7 (geïllustreerd in het groen). De resulterende celcyclus fase verdelingen van elk monster wordt uitgedrukt als% van de totale BrdU-positieve cellen per fase. Alleen de tijd punten te selecteren voor elke cel lijn worden weergegeven, zoals de analyses voor een aantal van de eerdere tijdstippen geproduceerd ongeldige resultaten. In deze eerdere tijdstippen, alle BrdU-positieve cellen in S-fase en daardoor geen afzonderlijke G1 en G2 pieken, die noodzakelijk zijn voor toepassing algoritme. (B) histogrammen van kankercellen met de kinetiek van doorlopen van G2 / M fasen van de celcyclus. Progression via G2 / M fasen is het snelst voor de LS180 cellen, gevolgd door HT29 en MCF7. Klik hier om een grotere afbeelding te bekijken .
jpg "alt =" Figuur S1 "/>
Figuur S1. Flowcytometrie controles. MCF7 cellen worden weergegeven als (A) negatieve controle waar de cellen zijn niet PI, noch BrdU gekleurd. (B) PI slechts gekleurd. (C) BrdU-FITC gemerkte monsters.

Figuur S2. Schematische die de relatie tussen fluorescentie emissiespektrum van PI versus FITC (BrdU). De spectrale verzameling vensters voor fluorescentielampen kanaal 1 (FL1 voor FITC) en fluorescerende kanaal 3 (FL3 voor PI) wordt in overeenkomstige vakken. Er is geen fluorofoor spectra overlap tussen FL1 en FL3 detectie. Blijkbaar fluorescentie compensatie niet nodig als experimentele gegevens PI en FITC-BrdU co-gelabelde cellen verzameld in FL1/FL3 kanalen respectievelijk. Fluorescentie spectra werden verkregen van BD BioSciences website:target = "_blank"> http://www.bdbiosciences.com/research/multicolor/spectrum_viewer. Klik hier om een grotere afbeelding te bekijken .
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Door het combineren van flowcytometrie met BrdU incorporatie, hebben we de nodige tools om kinetiek van de celcyclus te bestuderen. De kenmerkende eigenschap van BrdU om te functioneren als een analoog van thymidine is wat zorgt voor een DNA-inhoud kwantificering van een fiets-cel. De opname van BrdU in steeds dochter DNA tijdens de synthese fase van de celcyclus is wat het mogelijk is een subpopulatie van delende cellen volgen door de replicatie van DNA in S fase, gedurende groei G2 en uiteindelijk celdeling . Dochter BrdU-positieve cellen continu gevolgd tot welk moment de BrdU label wordt verdund door deling van de mate waarin het signaal verminderd tot achtergrondniveaus.
Met behulp van propidiumjodide om de totale DNA-gehalte te bepalen, zijn we in staat om voortgang van de celcyclus volgt uit het Z → G2 / M → G1. De mogelijkheid om de fietsen van cellen te volgen is belangrijk en nuttig zijn in vele toepassingen, met name for pre-klinische studies met betrekking tot de celcyclus specifieke DNA schade inducerende medicijnen of tijdens het ontwikkelingsproces van geneesmiddelen aan de celcyclus stadium gevoeligheid van nieuwe verbindingen 11 te bepalen. Het grote voordeel van het gebruik van een metabole labeling techniek is het afvoeren van chemische synchronisatie cellen. Aangezien de meeste chemische behandelingen celcyclus induceren zelf genotoxisch kan ontleden van de celcyclus gevoeligheid werking van nieuwe verbindingen in aanwezigheid van deze stoffen niet haalbaar.
We toonden de veelzijdigheid van deze BrdU metabole etikettering techniek met PI co-kleuring. Met dit protocol hebben we met succes gedemonstreerd dat de toename in de celcyclus door G2 / M fase van de celcyclus kan worden waargenomen in telomerase-negatieve humane cellen die zijn getransformeerd met gedwongen enzymexpressie 10. Hoewel deze methode krachtig het meten van de kinetiek van de cel zoals cycli through meerdere fasen een beperking is het verlies van tijdsinformatie tijdens de overgangen en in de G2 en M fasen. Zoals cellen doorloop G2 M, kunnen wij niet niet alleen de twee fasen onderscheiden, maar ook de sub-G1 populatie uitsluitend op DNA inhoud. In toepassingen waarbij deze differentiatie nodig is mogelijk kunnen co-vlek BrdU gelabelde cellen met antilichamen tegen cyclines of andere cellulaire eiwitten die uitdrukkingen zijn aan de gewenste fase van de celcyclus 12,13. Extra optimalisatie nodig dat de cycline antilichamen verenigbaar zijn met de BrdU label en immuno-kleuring protocol. In het ideale geval adequaat op de verschillende fasen van de celcyclus te onderscheiden, kunnen verschillende cycline antilichamen nodig zijn om co-label een enkel monster tegelijk. Ons laboratorium werkt aan de post-etikettering van BrdU-puls gelabelde cellen met cycline B en fosfo histon 3-antilichamen om onderscheid te maken tussen cellenin G2 versus M fase van de celcyclus. Co-labeling met cycline antilichamen naast PI en FITC-BrdU kleuring verhoogt aanzienlijk het complicatie van flowcytometrie zowel hardware en-analyse (fluorescentie schadevergoedingskwesties) perspectief. Met de komst van de nieuwe generatie flowcytometers, moet de vooruitgang in de capaciteit en functionaliteit te verlichten deze technische problemen.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Wij hebben niets te onthullen.
Acknowledgments
Wij danken Andy Johnson van de Biomedical Research Center at UBC voor hulp bij FACS-analyse. Cancer Research financiering in de Wong laboratorium wordt geleverd door de Canadian Cancer Society Research Institute (exploitatiesubsidie # 019250), en van O Herbelegging fondsen van de Faculteit Farmaceutische Wetenschappen, UBC. JMYW wordt ondersteund door de Canada Research voorzitters en de Michael Smith Foundation for Health Research Career Development-programma's.
Materials
| Name | Company | Catalog Number | Comments |
| bromodeoxyuridine | BD Biosciences | 55089 | |
| propidium iodide | Sigma-Aldrich | 287075 | 1mg/ml stock |
| FITC anti-BrdU | BD Biosciences | 347583 | |
| sodium tetraborate | Fisher Scientific | S80172 | 0.1M, pH 8.5 |
| FACS Caliber | BD Biosciences |
References
- Musgrove, E. A., Caldon, C. E., Barraclough, J., Stone, A., Sutherland, R. L. Cyclin D as a therapeutic target in cancer. Nat. Rev. Cancer. 11, 558-572 (2011).
- Molchadsky, A., Rivlin, N., Brosh, R., Rotter, V., Sarig, R. p53 is balancing development, differentiation and de-differentiation to assure cancer prevention. Carcinogenesis. 31, 1501-1508 (2010).
- Dickson, M. A., Schwartz, G. K. Development of cell-cycle inhibitors for cancer therapy. Curr. Oncol. 16, 36-43 (2009).
- Banfalvi, G. Cell cycle synchronization of animal cells and nuclei by centrifugal elutriation. Nat. Protoc. 3, 663-673 (2008).
- van Opstal, A., Boonstra, J. Inhibitors of phosphatidylinositol 3-kinase activity prevent cell cycle progression and induce apoptosis at the M/G1 transition in CHO cells. Cell Mol. Life Sci. 63, 220-228 (2006).
- Gasnereau, I. Flow cytometry to sort mammalian cells in cytokinesis. Cytometry. A. 71, 1-7 (2007).
- Harper, J. V. Synchronization of cell populations in G1/S and G2/M phases of the cell cycle. Methods Mol. Biol. 296, 157-166 (2005).
- Pedrali-Noy, G. Synchronization of HeLa cell cultures by inhibition of DNA polymerase alpha with aphidicolin. Nucleic Acids Res. 8, 377-387 (1980).
- Merrill, G. F.
- Fleisig, H. B., Wong, J. M. Telomerase promotes efficient cell cycle kinetics and confers growth advantage to telomerase-negative transformed human cells. Oncogene. , (2011).
- Cai, D., Byth, K. F., Shapiro, G. I. AZ703, an imidazo[1,2-a]pyridine inhibitor of cyclin-dependent kinases 1 and 2, induces E2F-1-dependent apoptosis enhanced by depletion of cyclin-dependent kinase 9. Cancer Res. 66, 435-444 (2006).
- Pozarowski, P., Darzynkiewicz, Z. Analysis of cell cycle by flow cytometry. Methods Mol. Biol. 281, 301-311 (2004).
- Sherwood, S. W., Rush, D. F., Kung, A. L., Schimke, R. T. Cyclin B1 expression in HeLa S3 cells studied by flow cytometry. Exp. Cell Res. 211, 275-281 (1994).

{kind=link}
{kind=link}