

Summary
Hücre döngüsü aşamaları ilerleme ve kinetikleri izleme ince değişiklikler Propidium iyodid ile BrdU ve toplam genomik DNA boyama yöntemi ile, nükleik asit metabolik etiketlenmesi bir kombinasyonu kullanılarak gerçekleştirilebilir. Bu yöntem, dolayısıyla da hücre siklus progresyonu etkileyen non-spesifik DNA hasarına, geliştirilmesinin önlenmesi, bisiklet hücrelerin kimyasal senkronizasyonu ihtiyacını ortadan kaldırır.
Abstract
Hücre döngüsünün çeşitli aşamalarında başlaması ve sonraki ilerleme Hassas kontrol hücreleri prolifere olarak büyük öneme sahiptir. Hücre döngüsü bölünmesi, büyüme ve önemli hücre döngüsü bileşenlerinin üretimi ve deregülasyon karsinogenez 1,2 tetikleyici faktörler sorumlu tutulmuştur ayrılmaz bir parçasıdır. Anti-kanser tedavileri Moleküler ilaçlar sıklıkla hücre döngüsü bölünmesi 3 düzenlenmesi ve koordinasyonundan sorumlu biyolojik yollar hedef. Hücre döngüsü kinetikleri hücre türüne göre değişir olmalarına rağmen, hücre döngüsünün dört aşamada arasında hücrelerinin dağılımını mitojen ve büyüme faktörü ifade tutarlı bir desenine bağlı olarak, belirli bir hücre hattı içinde daha çok tutarlıdır. Genotoksik olayları ve diğer hücre-stres katali için izin vermek için yakalanması veya belirli bir hücre döngüsü aşamasında geçici bir duraklama sonuçlanan, hücre döngüsü progresyon geçici blok sonuçlanabiliruygun tepki mekanizması kolayca dolaşmak mümkündür.
Deneysel olarak da hücre döngüsü ilerleme aşamasında referans ile bir hücre popülasyonunun davranışlarını gözlemlemek için yeteneği hücre biyolojisinde önemli bir ilerlemedir. Gibi mitotik olarak ortak prosedürlerin hücre döngüsü 4-6 belirli aşamalarında hücrelerin izole etmek için kullanılır, diferansiyel santrifüj veya akım sitometri tabanlı sıralama kurtulamıyorum. Bunlar, fraksiyonlara, hücre döngüsü faz daha sonra popülasyonlar zenginleştirilmiş deneysel tedavi tabi tutulur. Ayrılmış fraksiyonların Verim, saflık ve canlılık sıklıkla bu fiziksel ayırma yöntemleri kullanarak tehlikeye girebilir. Aynı zamanda, hücre popülasyonlarının ayırma ve fraksiyonlara hücreleri seçili hücre döngüsü aşamasından ilerleyebilir sayede deneysel tedavi başlangıcı arasındaki zaman atlamalı, başarılı bir şekilde uygulanması ve bu deneylerin yorumunda önemli sorunlar oluşturabilmektedir.
St diğer yaklaşımlarKamuoyu iliĢkileri programları hücre döngüsü aşamalarında hücreleri senkronize etmek kimyasalların kullanımını içerir. Her bir hücre döngüsü aşaması için anahtar metabolik işlemler kimyasal inhibitörleri ile hücre tedavisi sonraki aşamaya hücre döngüsünün ilerlemesini engelleme faydalıdırlar. Örneğin, tarafından G1 / S kavşakta ribonükleotid redüktaz inhibitörü hidroksiüre durur hücreleri deoxynucleotides, DNA'nın yapıtaşlarını tedarik sınırlayıcı. Diğer önemli kimyasallar aphidicolin tedavisi dahil, mitotik iğ oluşumunu engelleyebilir, her ikisi de G1 tutuklama polimeraz alfa inhibitörü, kolşisin ve Nokodazol ile tedavi, M fazındaki hücrelerin durdurmak için ve nihayet, DNA zinciri terminatör 5-fluorodeoxyridine ile tedavi başlatmak için S fazı tutuklama 7-9. Bu kimyasallar ile tedavi belirli bir aşamasında hücre bütün bir nüfusun eşitleyerek etkili bir araçtır. Kimyasal kaldırılması ile, hücreler ağızdan hücre döngüsü katın. Test maddesi aşağıdaki sürümü Tedavisihücre döngüsü engelleme kimyasal noktalarında ortaya çıkan madde, bir yanıt üniform, hücre döngüsü aşaması spesifik popülasyondan olmasını sağlar. Ancak, kimyasal eşzamanlayıcılar birçok beri meydan okuyor çeşitli yanıt yollar (test ajanlar vs synchronizers için) katılımı dışında alay, genotoksik bilinen bileşiklerdir.
Burada bölünme ve kızları hücre bölünmeye kadar, DNA'nın kopyalanması aşamasında onların ilerlemesini aktif olarak bisiklet hücrelerinin bir alt takip etmek için metabolik bir etiketleme yöntemi açıklar. Sitometrisi miktar ile birleştiğinde, bu protokol yokluğunda hücre döngüsü kinetik ilerleme ölçümü için olanak ya mekanik veya kimyasal kaynaklı hücresel genellikle diğer hücre döngüsü senkronizasyon metodolojiler 10 ile ilişkili olduğunu vurguluyor. Aşağıdaki bölümlerde biz de biyomedikal araştırmalarda bazı uygulamalarını olarak, metodoloji tartışacağız.
Protocol
1. Hücre Hazırlık

- Yaklaşık% 60 konfluent bir yoğunluğa ulaşmak üzere plakası hücreleri. Hücreler toplama zamanda log fazında olmalıdır. Mcf7 hücreleri için, bu 5 x 10 5 hücre / uygun bir ortamda 10 cm plakasına tohumlama gerçekleştirilir. HT29 ve LS180 hücreleri 6 x 10 5 hücre / 6 cm plakasına tohumlanmıştır. Biz, DMEM ortam% 10 FBS ve 1 x penisilin / streptomisin ile desteklenmiş kullanılır. Doğru pozitif ve negatif kontroller için tohum plakaları emin olunTest numunesinin yanında. Bunlar, aşağıdakileri içerir:
| ben | pozitif kontrol | Sadece BrdU |
| ii | pozitif kontrol | PI sadece |
| iii | negatif kontrol | BrdU negatif, PI negatif |
(Bu tür Bu protokol anlatılan biri olarak birden timepoints sahip bir ön deneyi gelecek koleksiyon yürütmek için bu süre boyunca zaman çerçevesi sınırlamak için kullanılabileceğini belirtmek gerekir.)
- Kaplamalı hücreler 37 C'de inkübe edilir ° C'de,% 5 24-48 saat boyunca CO 2, böylece hücreler kurtarma ve bağlamak için olanak sağlar.
- Bromodeoxyuridine (BrdU) ile etiket hücreleri nabızlamak için, DMEM wi değiştirilir10 uM BrdU içeren inci taze medya. Hücreler ° C'de,% 5 CO2 DNA'ya BrdU dahil edilmesi için izin vermek üzere 37 az 1 saat süre ile inkübe edilir. Negatif kontrol olarak hareket etmek işlenmemiş bir tabak bıraktığınızdan emin olun.
- Puls-etiketleme ortam kaldırılır ve hücreler 1X PBS ile kısaca durulanır.
- Taze büyüme ortamı eklenir (eksi BrdU) ve hücreler ° C'de,% 5 CO2 toplama için uygun timepoint ulaşılıncaya kadar, 37 inkübe devam izin verilir.
2. Hasat ve Fiksasyon
- Adım 1.3 ila tedavi edilmemiş hücrelerin sıfır timepoint olarak kabul edilir. Bu örnek, BrdU tedavisi hemen sonra toplanır 1 saat timepoint birlikte toplanabilirler.
- Hücreler hasat etmek için, ortam kaldırılır ve plakalar 1X bir kez PBS ile durulanır.
- Hücreler 5 dak Sup için 1500 rpm'de santrifüje edilerek, tripsinize kültür ortamı içinde toplanmış ve daha sonra topak haline edilirernatant atılır.
- 5 ml buz gibi soğuk 1X PBS ile 1-10 x 10 6 hücre durulayın. 5 dakika Fazı için 1500 rpm'de santrifüj atılır.
- Hücre topağı sonra buz gibi soğuk PBS /% 1 FBS 100 ul içinde tekrar süspanse edilir. Hücre topaklanma önlemede PBS yardımcıları% 1 FBS eklenmesi.
- -20 ° C, hücreler 70 sabitlemek için% 5 ml etanol ile damla damla ekleme bu hücreler.
- 4 ° C'da 30 dakika veya saklamak için buz üzerinde inkübe. Bu, çeşitli timepoints örneklerinin toplanması durdurmak için hangi ideal bir adımdır. Bunların örnekleri protokolünün kalanı ile eş zamanlı olarak işleme izin vererek, birkaç gün boyunca etanol içinde bırakılabilir.
3. BrdU ve PI Boyama
- 1500 rpm'de 5 dakika için santrifüjleme ile pelet hücreleri.
- Sabitleştirici çıkarın, ancak ~ 50 ul vortekslendi tarafından pelet gevşetmek için hangi bırakın.
- DNA denatüre yavaşça 2N HCI / Triton X-100 ile 1 mLvorteks ederken damla damla. 30 dakika oda sıcaklığında inkübe örnekleri.
- 1500 rpm'de 5 dakika örnekleri santrifüj edilerek pelet hücreleri. Aspire ve supernatant atın.
- Denatürasyon adım nötralize etmek için, 0.1M sodyum tetraborat, pH 8.5 1 ml hücre pelletini.
- 1500 rpm'de 5 dakika hücreleri santrifüjleyin. Aspire ve supernatant atın.
- BrdU boyama karışımı 75 ul (50 ul% 0.5 tween 20/1% BSA / PBS + 20 ul FITC konjuge anti-BrdU + 5 ul 10 mg / ml RNase) içerisinde hücre pelletini.
- Işıktan korunan 45 dakika oda sıcaklığında inkübe edilir.
- Pelet hücreleri, 1500 rpm'de 5 dakika boyunca örnekleri santrifüjlenir. Aspire ve supernatant atın.
- PBS 5 ug / ml propidium iyodür ihtiva eden 1 ml hücre pelletini.
4. Akım Sitometri
- Hücrelerin analiz için, bir akış sitometresi 488 nm lazer ve donanımlıUygun filtreler gereklidir. Böyle CellQuest gibi analiz yazılımı aşağıda özetlenen araziler yaratmak için gereklidir.
- Akış sitometresi ile örnek çalıştırırken, hücrelerin düzgün boyut dağılımı (Şekil 1A) sağlamak için bir forward scatter (SSC-H) karşı tarafında dağılım (FSC-H) arsa oluşturun. Bu parametreler her iki doğrusal bir ölçekte çizilmiştir.
- Aynı zamanda FL3-H (PI leke) vs FL2-W (Şekil 1B) bir arsa görüntülemek. Bu plot hücre döngüsünün normal dağılım deseni dahilinde hücre fraksiyonu izole etmek için bir geçit (R3) oluşturmak için kullanılır. Genel olarak, bu dağılım deseni sırasıyla hücre döngüsünün G1 ve G2 fazlar, temsil PI boyamanın 2N ve 4N DNA içeriği hücreleri iki yığın ile karakterize edilir. Bu 2 kümeleri arasında bulunan hücrelerin bir dize hücre döngüsünün S fazında meydana devam eden DNA replikasyonu temsilcisidir.
- Bir komplo displayi oluşturmang y-ekseni ve x-ekseni (Şekil 1C) üzerindeki PI (lineer arsa) üzerinde FL1-H ile adım 4.3, (FITC, günlük arsa) gelen kapılı hücreleri (R3). Bu plot analizi için veri toplamak için çeşitli 1,1 belirtilen kontrol yanı sıra kullanılarak geri kalan parametreleri belirlemek için kullanılır.
- Sitometresinin üzerine PI sadece lekeli örneği yerleştirin. X-ekseninde ~ 200 G1 yerleştirmek için kazanç ayarlayın. Bu leke PI bir histogram grafiği de görselleştirmek için daha kolay olacaktır.
- İkinci bir denetim akış sitometresinde bir BrdU-tek numune çalışan içerir. Hücrelerin 2 popülasyonları (BrdU pozitif vs BrdU negatif) arsa üzerinde görünür ki kazanç ayarlayın. İdeal olarak, BrdU negatif hücrelerin sadece 10 -1 altında görüntülenmesini konumlandırılmış.
- Son kontrol BrdU / PI negatif örneğidir. Hücreleri üst veya sağ kadran herhangi görünmez olmasını sağlamak için bu negatif kontrol çalıştırın.
- Çeşitli araziler parametreleri ayarlandıktan sonra,Akım sitometrisi BrdU ve PI boyalı hücrelerin işlenmesi için kalibre ve hazırdır. Doğru PI fraksiyonu (4.3 adıma bakınız) ortasında korunaklı olan 10.000 hücrelerin en az tahsilat okumak gerekir.
- Toplanan verileri analiz etmek, bu FlowJo veya FacsDiva gibi yazılımları kullanılmaktadır. Yukarıda bahsedilen parselleri her bu programlara yeniden ve niceliksel ve İstatistiksel analiz gerçekleştirilebilir.
- Analizin son nokta art arda birkaç adım içerir. PI vs FL2-W arsa olumlu PI kapılı hücrelerle FITC vs PI bir komplo oluşturma bir bisiklet nüfus ayırmasına olanak sağlar. İkinci bir kapı BrdU pozitif nüfusu izole ederek bu komplo oluşturulur. X ekseni üzerinde PI ile bir histogram arsa üzerinde bu farklı nüfus ifade bir hücre döngüsü ilerleme zamanı ders izlemenize olanak sağlar. Bu G1 veya zamanın bir fonksiyonu olarak G2 içeriğine sahip BrdU-pozitif hücrelerin sayısı çizilmesi ile daha da görüntülenmiştir olabilir.
5.. Temsilcisi Sonuçlar
Normalde propidyum iyodür ile boyanmış bisiklet hücrelerin sırasıyla, 2N ve 4N DNA içeriği içeren hücreleri (Şekil 2) karşılık gelen G1 ve G2 farklı zirveleri var. BrdU Darbe etiketleme aktif DNA (yani S fazı) sentezleyerek hücrelerden bir alt nüfusun seçici etiketleme için izin verir. Kısa bir süre BrdU reaktifi uzaklaştırıldıktan sonra, tüm etiketli hücreler S fazı (Şekil 3) bulunmaktadır. Onlar hücrenin döngüsünün sonraki aşamalarında geçerken kısa bir darbe etiketleme kısıtlayarak bir çok kez puan boyunca hücrelerin bu şimdi ayrı alt-nüfus takip edebiliyor. Bu, bir G1 ayrı olmaması ve G2 zirveleri Şekil 3'te 1 saat bir zaman noktasında görüntülenmiştir olabilir.
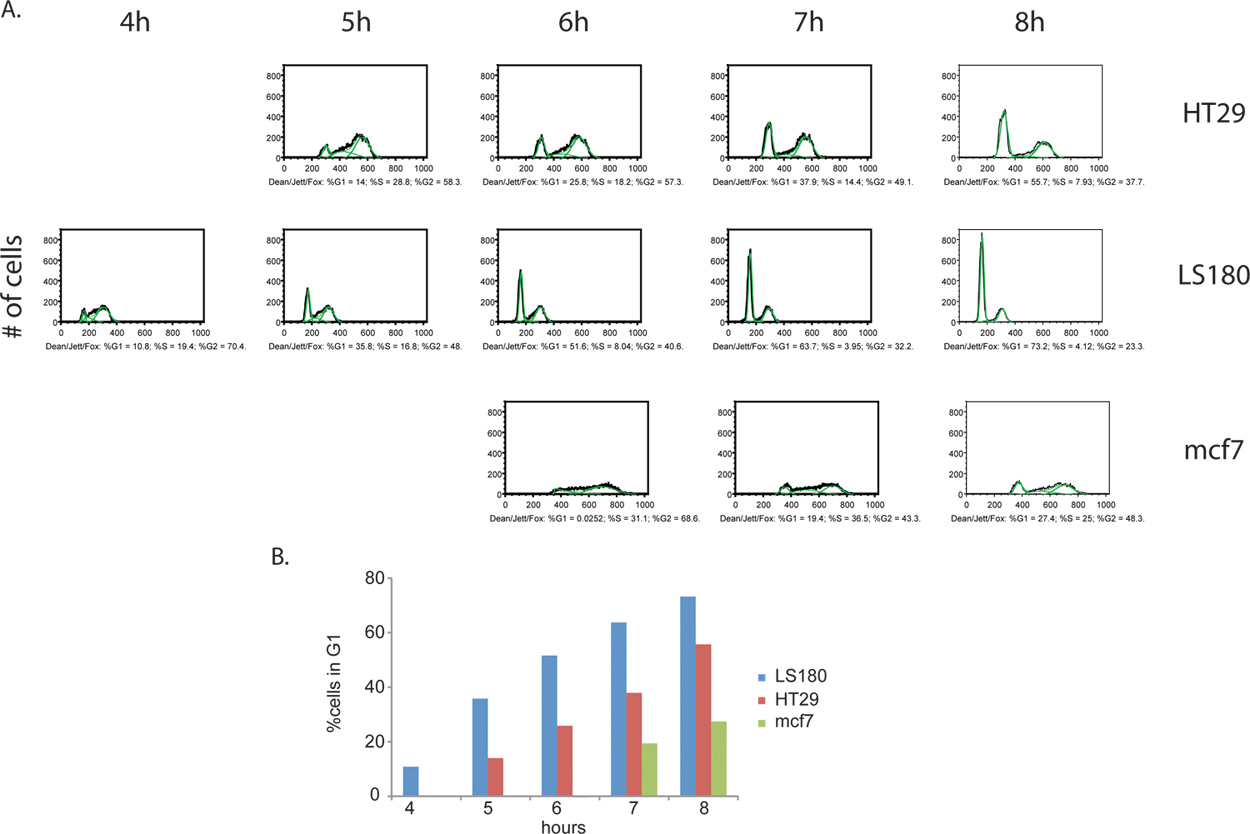
Ek bilgi BrdU etiketleme adım elde edilebilir. Bölünen hücre oranı, bir est ölçülen, fakat sadece edilebiliriki örnek arasındaki hücre döngüsü aşamasında dağıtım mak de tespit edilebilir. Onlar G2 döngüsü, ilerleme devam ettikçe BrdU reaktif çıkarılmasını takiben eşit aralıklı aralıklarla hücrelerin toplanması ile, hücreler izlenebilir ve sonunda nihayet G1 ile BrdU pozitif kızı hücreler olarak ortaya çıkan, orijinal hücre hücre bölünmesi için hareket DNA içeriği (Şekil 4). Hücre döngüsü faz miktar ve kinetik analizi için bir örnek Şekil 5 'de verilmiştir.

Şekil 1. Temsilcisi mcf7 hücre popülasyonunun (A) Forward Scatter karşı Side Scatter Plot. Bir temsilci mcf7 hücre popülasyonunun (B) PI karşı Genişlik (FL-W) Arsa. Perdeleme son analizde (R3) hücre çiftler çifti çıkarmak için gösterilmiştir. Bu lazer ışını ve Şeref geçmesi için daha uzun sürer gibi hücre çiftler çifti, tek bir hücre daha büyük puls genişliğine sahip olacakcevher analize dahil edilebilir. Bir temsilci mcf7 hücre popülasyonunun (C) PI karşı FITC (BrdU) Arsa. Perdeleme yalnızca FITC (BrdU) pozitif hücrelerin (R4) içerecek şekilde gösterilmiştir.

Şekil 2. Normalde bisiklet hücrelerinin toplam nüfusu Temsilcisi histogram grafiği.

Şekil 3. FITC (BrdU)-pozitif hücreleri için perdeleme sonra, BrdU puls çıkarıldıktan sonra 1H toplanmıştır hücrelerinin histogram plot. Etiketin çıkarılmasını takiben bir erken timepoint BrdU-pozitif hücreler sadece DNA sentezi esnasında hücre başarılı bir etiketleme doğrulayan, hücre döngüsünün S-fazlı in olan hücreleri ile tutarlıdır DNA içeriğini gösteren örnekleri karşılık PI profillerini gösterir.
 lt = "Şekil 4" />
lt = "Şekil 4" />
Şekil 4. Kanser hücresi hatlarında, kolorektal kanserler HT29 (A) ve LS180 (B) yanı sıra, meme kanseri mcf7 (C) arasındaki hücre döngüsü ilerlemesi kinetikleri karşılaştırılması. Hücreler BrdU darbe çıkarılmasını takiben, 8 saat boyunca her saat toplanmıştır. Bu deneyde, kolorektal hücre hattı LS180 hücre döngüsünün G2 fazı ile hızlandırılmış hücre döngüsü sürecinde net bir profil gözlenmiştir. Kolorektal kanser hücrelerinin iki profil arasında kinetikleri karşılaştırarak, bir G1 pik ortaya çıkması, bu pik yoksun HT-29 hücre çizgisi için uygun bir zaman noktası ile karşılaştırıldığında, LS180 hücreleri, T = 4H post-BrdU belirgindir. İki kolorektal hücre hatları biri biriyle karşılaştırıldığında, mcf7 hücreler önemli ölçüde azalmayla bisiklet edilir. büyük rakam görmek için buraya tıklayın .
yeniden 5 "/>
Şekil 5. BrdU-etiketli kanser hücreleri (A) Kantitatif hücre döngüsü faz analizi. Dekan / Jett / Fox algoritma HT29, LS180 ve mcf7 (yeşil resimli) uygulandı. Her numuneden elde edilen hücre döngüsü faz dağılım her aşama için% toplam BrdU-pozitif hücreler olarak ifade edilir. Önceki zaman noktalarının bazı analizler geçersiz sonuç olarak sadece her hücre hattı için zaman noktaları seçmek, görüntülenir. Bu daha erken zaman noktalarında, tüm BrdU-pozitif hücreleri S-faz ve bu nedenle algoritması uygulama için gerekli olan ayrı G1 ve G2 zirveleri, yoksundur. (B), kanser hücrelerinin Histogramlar hücre döngüsünün G2 / M aşamalarında kinetiği progresyon gösteren. G2 / M aşamalarında İlerleme HT29 ve mcf7 takip LS180 hücreleri için hızlı. büyük rakam görmek için buraya tıklayın .
jpg "alt =" Şekil S1 "/>
Şekil S1. Flow sitometri kontrolleri. mcf7 hücreler PI ne BrdU boyanmış ne (A) negatif kontrol olarak gösterilir. (B) PI sadece boyandı. (C) BrdU-FITC etiketli örnekleri.

PI karşı FITC (BrdU) floresan emisyon spektrumu arasındaki ilişkiyi gösteren S2. Şematik gösterimi Şekil. Flüoresan kanalı 1 (FL1, FITC) ve flüoresan kanalı 3 (FL3, PI) için spektral toplama pencere karşılık gelen kutular içinde gösterilmiştir. FL1 ve FL3 algılama arasında fluorofor spektrumları örtüşme yoktur. PI ve FITC-BrdU gelen deneysel verileri eş-etiketli hücreler sırası FL1/FL3 kanallar toplanır ettiği bir floresans dengeleme gerekli değildir. Floresans spektrumları BD BioSciences web sitesinden elde edilmiştir:target = "_blank"> http://www.bdbiosciences.com/research/multicolor/spectrum_viewer. büyük rakam görmek için buraya tıklayın .
Subscription Required. Please recommend JoVE to your librarian.
Discussion
BrdU eklenmesi ile akım sitometri birleştirerek, hücre döngüsü kinetiğini incelemek için gerekli araçlara sahip. Bir timidin analoğu olarak işlev BrdU en ayırt edici özelliği bir bisiklet hücrenin DNA içeriği ölçümü için olanak budur. Hücre siklusu sentez aşamasında büyüyen bir kızı DNA zincir haline BrdU ile birleşmesiyle bir G2 büyüme dönemi ve sonuçta hücre bölünmesi, S fazında DNA çoğaltma yoluyla bisiklet hücrelerinin bir alt izlemenizi sağlar nedir . BrdU etiketin sinyal seviyeleri arka indirgenir derecede hücre bölünmesi ile seyreltilir zamanda hangi noktaya dek Daughter-pozitif hücrelerin BrdU sürekli izlenebilir.
Toplam DNA içeriği belirlemek için propidyum iyodür kullanarak, S → G2 / M → G1 hücre döngüsünün ilerlemesini takip edebiliyoruz. Hücre döngüsü oranı izlemek için özellikle fo yeteneği, çok uygulamada kullanılabilir ve önemli olduğuyeni bileşiklerin 11 hücre döngüsü aşamasında duyarlılığı belirlemek için hücre döngüsü spesifik DNA hasarı oluşturan ilaçlar veya ilaç geliştirme sürecinde içeren r ön-klinik çalışmalar. Metabolik bir etiketleme tekniği kullanarak en belirgin avantajı hücrelerin kimyasal senkronizasyonu ortadan kaldırılmasıdır. Hücre siklusu durmasına neden en kimyasal işlemler genotoksik kendileri gibi, bu kimyasalların varlığında yeni bileşiklerin hücre döngüsü duyarlılığı etkileri kesme uygun olmayabilir.
Biz PI co-boyama ile bu BrdU metabolik etiketleme tekniğinin çok yönlülüğü. Bu protokolü kullanarak, başarılı hücre döngüsünün G2 / M faz üzerinden hücre döngüsünün ilerlemesinde artış enzim ifade 10 zorladı dönüştürülmüştür edilmiştir telomeraz negatif insan hücrelerinde de görülebiliyor göstermiştir. Bu yöntem, bir uçtan bir uca döngüleri olarak hücre ölçülerek kinetikleri güçlü ikenÇoklu aşamalar ugh, bir sınırlandırma geçişleri sırasında zamanlama bilgi kaybının yanı sıra, G2 ve M fazlar dahilindedir. Hücreleri M G2 boyunca ilerlerken, biz sadece DNA içeriğine göre, iki aşamada sadece ayırt edemez, aynı zamanda alt-G1 nüfus. Bu farklılaşma gerektiren uygulamalarda, bu siklinler veya olan ifadeler hücre döngüsü 12,13 arasında arzu edilen faz özgü başka bir hücresel protein karşı antikorları ile birlikte leke BrdU etiketli hücreler için potansiyel olarak mümkündür. Ek optimizasyon siklin antikorlar BrdU etiket ve immüno-boyama protokolü ile uyumlu olmasını sağlamak için gerekli olacak. İdeal olarak, yeterli hücre döngüsünün çeşitli aşamalarında ayırt etmek için, birkaç siklin antikorlar tek bir örnek aynı anda co-etiket gerekebilir. Laboratuvarımız hücreleri ayırt siklin B ve fosfor Histon 3 antikorlar ile BrdU-darbe etiketli hücrelerinin post-etiketleme üzerinde çalışıyorhücre döngüsünün G2 karşılık M fazında. PI ve FITC-BrdU boyama ek olarak siklin antikorlar ile Co-etiketleme, önemli bir donanım ve veri analizi (floresans tazminat konular) açısından her iki akım sitometri komplikasyon artırır. Flow sitometrinin yeni nesil ile birlikte, kendi kapasite ve işlevselliğe gelişmeler bu teknik kaygıları hafifletmek gerekir.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Biz ifşa hiçbir şey yok.
Acknowledgments
Biz FACS analizi ile yardım için at UBC Biyomedikal Araştırma Merkezi Andy Johnson teşekkür ederim. Wong Laboratuvar Kanser Araştırma Fonu Kanada Kanser Topluluğu Araştırma Enstitüsü (işletme hibesi # 019250) tarafından ve İlaç Bilimleri, UBC Fakültesi Araştırma yeniden yatırım fonlarından sağlanır. JMYW Kanada Araştırma Sandalyeler ve Sağlık Araştırması Kariyer Geliştirme programları için Michael Smith Vakfı tarafından desteklenmektedir.
Materials
| Name | Company | Catalog Number | Comments |
| bromodeoxyuridine | BD Biosciences | 55089 | |
| propidium iodide | Sigma-Aldrich | 287075 | 1mg/ml stock |
| FITC anti-BrdU | BD Biosciences | 347583 | |
| sodium tetraborate | Fisher Scientific | S80172 | 0.1M, pH 8.5 |
| FACS Caliber | BD Biosciences |
References
- Musgrove, E. A., Caldon, C. E., Barraclough, J., Stone, A., Sutherland, R. L. Cyclin D as a therapeutic target in cancer. Nat. Rev. Cancer. 11, 558-572 (2011).
- Molchadsky, A., Rivlin, N., Brosh, R., Rotter, V., Sarig, R. p53 is balancing development, differentiation and de-differentiation to assure cancer prevention. Carcinogenesis. 31, 1501-1508 (2010).
- Dickson, M. A., Schwartz, G. K. Development of cell-cycle inhibitors for cancer therapy. Curr. Oncol. 16, 36-43 (2009).
- Banfalvi, G. Cell cycle synchronization of animal cells and nuclei by centrifugal elutriation. Nat. Protoc. 3, 663-673 (2008).
- van Opstal, A., Boonstra, J. Inhibitors of phosphatidylinositol 3-kinase activity prevent cell cycle progression and induce apoptosis at the M/G1 transition in CHO cells. Cell Mol. Life Sci. 63, 220-228 (2006).
- Gasnereau, I. Flow cytometry to sort mammalian cells in cytokinesis. Cytometry. A. 71, 1-7 (2007).
- Harper, J. V. Synchronization of cell populations in G1/S and G2/M phases of the cell cycle. Methods Mol. Biol. 296, 157-166 (2005).
- Pedrali-Noy, G. Synchronization of HeLa cell cultures by inhibition of DNA polymerase alpha with aphidicolin. Nucleic Acids Res. 8, 377-387 (1980).
- Merrill, G. F.
- Fleisig, H. B., Wong, J. M. Telomerase promotes efficient cell cycle kinetics and confers growth advantage to telomerase-negative transformed human cells. Oncogene. , (2011).
- Cai, D., Byth, K. F., Shapiro, G. I. AZ703, an imidazo[1,2-a]pyridine inhibitor of cyclin-dependent kinases 1 and 2, induces E2F-1-dependent apoptosis enhanced by depletion of cyclin-dependent kinase 9. Cancer Res. 66, 435-444 (2006).
- Pozarowski, P., Darzynkiewicz, Z. Analysis of cell cycle by flow cytometry. Methods Mol. Biol. 281, 301-311 (2004).
- Sherwood, S. W., Rush, D. F., Kung, A. L., Schimke, R. T. Cyclin B1 expression in HeLa S3 cells studied by flow cytometry. Exp. Cell Res. 211, 275-281 (1994).

{kind=link}
{kind=link}