

Summary
Sporing subtile endringer i progresjon og kinetikk av cellesyklus stadier kan oppnås ved bruk av en kombinasjon av metabolsk merking av nukleinsyrer med BrdU og total genomisk DNA farging via propidium jodid. Denne metoden unngår behovet for kjemisk synkronisering av sykling celler, og dermed hindre innføring av ikke-spesifikk DNA-skade, som igjen påvirker cellesyklus progresjon.
Abstract
Nøyaktig kontroll av initiering og påfølgende progresjon gjennom de ulike fasene av cellesyklus er av avgjørende betydning i voksende celler. Cellesyklus divisjon er en integrert del av vekst og reproduksjon og deregulering av sentrale cellesyklus komponenter har vært innblandet i de utløsende hendelsene i kreftutvikling 1,2. Molekylære agenter i anti-kreft terapier ofte målrette biologiske mekanismer er ansvarlige for regulering og koordinering av cellesyklus divisjon tre. Selv cellesyklus kinetikk tendens til å variere i henhold til celletype, er fordelingen av celler blant de fire fasene av cellesyklus heller konsekvent innenfor en bestemt cellelinje grunn konsistent mønster av mitogen og vekstfaktor uttrykk. Gentoksiske hendelser og andre cellulære stressfaktorer kan resultere i en midlertidig blokk med cellesyklusprogresjon, noe som resulterer i arrest eller en midlertidig pause i en bestemt celle syklus fase for å tillate instinavigasjonsknappene av riktig svar mekanisme.
Muligheten til å eksperimentelt observere atferden til en celle befolkning med referanse til deres cellesyklusprogresjon stadium er et viktig fremskritt i cellebiologi. Vanlige prosedyrer som mitotisk riste av, differensial sentrifugering eller flowcytometri-baserte sortering brukes til å isolere cellene på bestemte faser av cellesyklus 4-6. Disse Oppdelte, cellesyklus fase-beriket populasjoner blir deretter utsatt for eksperimentell behandling. Yield, renhet og levedyktighet av de utskilte fraksjoner kan ofte bli kompromittert ved hjelp av disse fysiske separasjonsmetoder. I tillegg kan tidsforløp mellom separasjon av celle populasjoner og i starten av eksperimentell behandling, der de fraksjonert cellene kan utvikle seg fra den valgte cellen syklus scenen, stiller betydelige utfordringer i en vellykket gjennomføring og tolkning av disse eksperimentene.
Andre tilnærminger til study cellesyklus stadier inkluderer bruk av kjemikalier for å synkronisere celler. Behandling av celler med kjemiske inhibitorer av sentrale metabolske prosesser for hver celle syklus stadium er nyttige i blokkerer progresjon av cellesyklus til neste stadium. For eksempel, de ribonukleotidreduktase hemmer hydroksyurea stopper cellene ved G1 / S tidspunkt ved å begrense tilførselen av deoxynucleotides, byggesteinene i DNA. Andre kjente kjemikalier omfatter behandling med aphidicolin, til en polymerase alfa-hemmer for G1 arrest, behandling med kolkisin og nocodazole, som begge forstyrre mitotisk spindel formasjon stanse celler i M fase og til slutt, til behandling med DNA-kjeden terminator 5-fluorodeoxyridine initiere S fase arrest 7-9. Behandling med disse kjemikaliene er et effektivt middel for å synkronisere en hel populasjon av celler på et bestemt fase. Med fjerning av kjemiske, celler gjenoppta cellesyklus i kor. Behandling av testen agenten følgende utgivelsefra cellen syklus blokkerer kjemisk sikrer at stoffet responsen utløses er fra en uniform, cellesyklus scene-spesifikk befolkningen. Men siden mange av de kjemiske synchronizers kjent gentoksiske stoffer, erting hverandre deltakelse av ulike respons pathways (til synchronizers vs test agentene) er utfordrende.
Her beskriver vi en metabolsk merking metode for å følge en undergruppe aktivt sykling celler gjennom sin progresjon fra DNA replikasjon fasen, gjennom til deling og utskillelse av deres datter celler. Sammen med flowcytometri kvantifisering, gjør denne protokoll for måling av kinetisk progresjon av cellesyklus i fravær av enten mekanisk eller kjemisk indusert mobilnettet understreker ofte assosiert med andre cellesyklus synkronisering metoder 10. I de følgende avsnittene vil vi diskutere metodikk, samt noen av sine applikasjoner i biomedisinsk forskning.
Protocol
1. Celleforberedelsen

- Plate celler for å oppnå en tetthet på ca 60% confluency. Celler må være i loggen fase på tidspunktet for samlingen. For mcf7 cellene, gjøres dette ved såing på 5 x 10 5 celler / 10 cm plate i de aktuelle mediene. HT29 og LS180 cellene blir seedet på 6 x 10 5 celler / 6 cm plate. Vi brukte DMEM media supplert med 10% FBS og 1x penicillin / streptomycin. Pass på å frø plater for de riktige positive og negative kontroller itillegg til stikkprøvene. De omfatter følgende:
| jeg | positiv kontroll | BrdU bare |
| ii | positiv kontroll | PI bare |
| iii | negativ kontroll | BrdU negativ, PI negativ |
(Det bør bemerkes at en foreløpig eksperiment med flere tidspunkt som skissert i denne protokollen kan brukes til å begrense tidsperiode som å utføre fremtidige samlinger.)
- De gullbelagte Cellene inkuberes ved 37 ° C, 5% CO 2 i 24-48 timer, og dermed lar cellene for å gjenopprette og fest.
- For å pulsere label celler med bromodeoxyuridine (BrdU), er DMEM erstattet with ferske medier som inneholder 10 mM BrdU. Celler inkuberes i 1 time ved 37 ° C, 5% CO 2 for å tillate BrdU innlemmelse i DNA. Sørg for å la en plate ubehandlet å fungere som den negative kontrollen.
- De puls-merking media fjernes og cellene blir skylt kort med 1X PBS.
- Ferske vekstmedier tilsettes (minus BrdU) og celler får lov til å fortsette å ruge ved 37 ° C, 5% CO 2 til den aktuelle endepunktet for høsting er nådd.
2. Høsting og fiksering
- Ubehandlede celler fra trinn 1,3 vurderes det null endepunktet. Denne prøven kan høstes sammen med en h endepunktet som samles inn umiddelbart etter BrdU behandling.
- Å høste cellene, blir media fjernet og platene skylles straks med 1X PBS.
- Celler er trypsinized, samles i dyrking media og senere pelletert ved sentrifugering ved 1500 rpm i 5 min Supernatant blir forkastet.
- Skyll 1-10 x 10 6 celler i 5 ml iskald 1X PBS. Sentrifuger ved 1500 rpm i 5 min supernatanten blir forkastet.
- Cellen pellet blir deretter resuspendert i 100 mL av iskald PBS / 1% FBS. Tilsetting av 1% FBS i PBS hjelpemidler i å forebygge celle klumper.
- Legg disse cellene dråpe for dråpe til 5 ml -20 ° C, 70% etanol for å fikse cellene.
- Inkuber på is i 30 min eller butikken ved 4 ° C over natten. Dette er en ideell skritt å stoppe innsamling av prøver fra de ulike tidspunkter. Prøvene kan stå i etanol i flere dager, slik at de kan behandles samtidig gjennom resten av protokollen.
3. BrdU og PI Farging
- Pellet celler ved sentrifugering i 5 min ved 1500 rpm.
- Fjern fiksativ, men la ~ 50 mL i å løsne pellet ved virvling.
- Slik denature DNA, sakte tilsett 1 ml av 2N HCL / Triton X-100dropwise mens virvling. Inkuber prøvene ved romtemperatur i 30 min.
- Pellet cellene ved sentrifugering prøvene i 5 min ved 1500 rpm. Aspirer og kast supernatanten.
- Resuspender cellepelleten i 1 ml 0.1m natrium tetraborate, pH 8,5, for å nøytralisere den denaturering trinnet.
- Sentrifuger cellene i 5 min ved 1500 rpm. Aspirer og kast supernatanten.
- Resuspender cellen pellet i 75 mL av BrdU flekker mix (50 mL 0,5% Tween 20/1% BSA / PBS + 20 mL FITC konjugert anti-BrdU + 5 mL 10 mg / ml RNase).
- Inkuber ved romtemperatur i 45 min beskyttet mot lys.
- Til pelletskaminer celler, sentrifuger prøvene i 5 min ved 1500 rpm. Aspirer og kast supernatanten.
- Resuspender cellepelleten i 1 ml PBS inneholder 5 mikrogram / ml propidium jodid.
4. Flowcytometrisystemer
- For å analysere cellene, en flowcytometer utstyrt med en 488 nm laser oghensiktsmessige filtre er nødvendig. Analyse programvare som CellQuest er nødvendig for å skape tomtene er skissert nedenfor.
- Når du kjører prøven gjennom strømningscytometeret, skape et framtidsrettet scatter (SSC-H) vs side scatter (FSC-H) plottet for å sikre riktig størrelse fordeling av celler (figur 1A). Begge disse parametrene er plottet på en lineær skala.
- Samtidig viser en tomt på FL3-H (PI flekker) vs FL2-W (Figur 1B). Denne tomten brukes til å lage en gate (R3) å isolere brøkdel av celler som er innenfor den normale fordelingen mønster av cellesyklus. Generelt er denne fordelingen mønsteret preget av to klynger av celler fra 2N og 4N DNA innhold av PI flekker representerer G1 og G2 faser av cellesyklus, henholdsvis. En streng av celler lokalisert mellom disse 2 klynger er representativ for pågående DNA replikasjon som skjer i S-fasen av cellesyklus.
- Lag et plott displaying de gated celler (R3) fra trinn 4,3, med FL1-H (FITC, log plott) på y-aksen og PI (lineær tomten) på x-aksen (figur 1C). Denne tomten vil bli brukt til å sette de resterende parametere med ulike kontrollene skissert i 1.1, samt å samle inn data for analyse.
- Plasser PI eneste fargede prøve på cytometeret. Juster forsterkningen å plassere G1 på ~ 200 på x-aksen. Dette vil være lettere å visualisere i et histogram plott av PI flekken.
- En annen kontroll innebærer å kjøre en BrdU bare prøve på strømningscytometeret. Juster forsterkningen slik at de 2 populasjoner av celler (BrdU positive vs BrdU negativ) vises på tomten. Ideelt sett er de BrdU negative cellene posisjonert til å vises rett under 10 -1.
- Den endelige kontrollen er BrdU / PI negativ prøve. Kjør denne negativ kontroll for å sikre at cellene ikke vises i noen av de øvre eller høyre hånd kvadranter.
- Når parametrene i de forskjellige tomter har blitt innstilt, detflowcytometeret er kalibrert og klar for behandling av BrdU og PI fargede celler. Et minimum på 10.000 celler som er inngjerdet i riktig PI fraksjonen (se trinn 4.3) må leses per samling.
- Å analysere de innsamlede dataene, er programvare som FlowJo eller FacsDiva utnyttet. Hver av de ovenfor nevnte tomter kan gjenskapes i disse programmene og kvantitativ og statistisk analyse utført.
- Endepunktet av analysen involverer flere suksessive trinn. Opprette en tomt på FITC vs PI med celler gated som PI positiv fra PI vs FL2-W tomten tillater en å skille sykling befolkningen. En annen gate er opprettet fra denne tomten ved å isolere BrdU positive befolkningen. Uttrykke dette karakteristiske befolkningen på en histogram tomt med PI på x-aksen gjør det mulig å spore tiden løpet av cellesyklusprogresjon. Dette kan være ytterligere visualisert ved å plotte antall BrdU-positive celler med G1 eller G2 innhold som en funksjon av tid.
5. Representative Resultater
Normalt sykling celler farget med propidium iodide har forskjellige topper på G1 og G2, tilsvarende celler som inneholder 2N og 4N DNA innhold, henholdsvis (figur 2). Pulse merking med BrdU tillater selektiv merking av en sub-populasjon av celler som er aktivt syntetisere DNA (dvs. S-fasen). Kort tid etter fjerning av BrdU reagens, alle merket celler er i S-fasen (figur 3). Ved å begrense merking til en kort puls man er i stand til å følge dette nå distinkt sub-populasjon av celler i hele utallige tidspunkter når de passerer gjennom de påfølgende fasene av cellesyklus. Dette kan visualiseres på en t tidspunkt i figur 3 som en tydelig mangel på G1 og G2 topper.
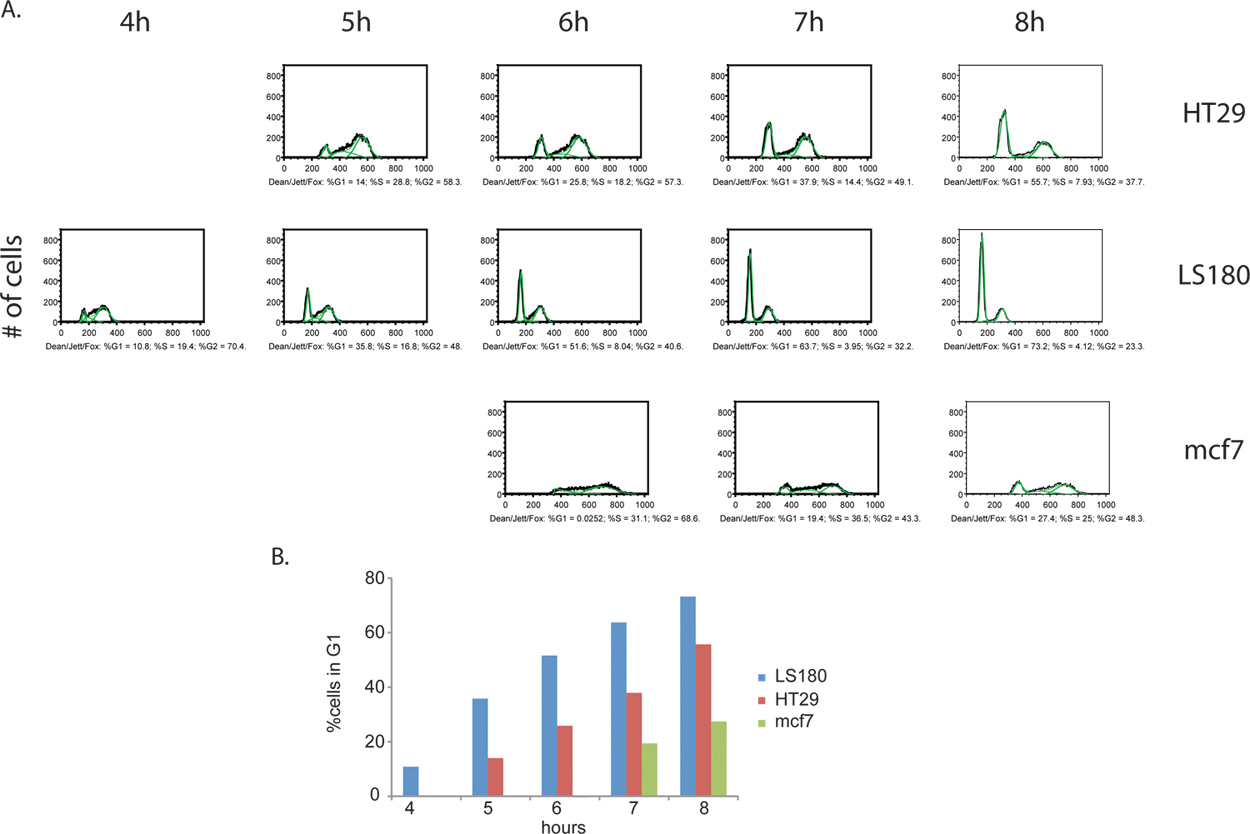
Ytterligere informasjon kan utledes fra BrdU merking trinnet. Ikke bare kan den andelen av aktivt delende celler måles, men en estimate av cellesyklus scenen fordeling mellom to prøvene kan bestemmes også. Ved å samle cellene ved jevne mellomrom mellomrom etter fjerning av BrdU reagens, kan cellene spores som de fortsetter å sykle, fremdrift til G2 og til slutt gå gjennom til celledeling av de opprinnelige cellene, endelig frem som BrdU-positive datterceller med G1 DNA-innhold (figur 4). Et eksempel på cellesyklus fase kvantifisering og kinetisk analyse er gitt i figur 5.

Figur 1. (A) Forward Scatter versus Side Scatter Plot av en representant mcf7 celle befolkningen. (B) PI versus Bredde (FL-W) Plot av en representant mcf7 celle befolkningen. Gating er vist å utelukke celle dubletter i den endelige analysen (R3). Cell dubletter vil ha større pulsbredde enn en enkelt celle, som de tar lengre tid å passere gjennom laserstrålen og therefmalm kan utelukkes fra analysen. (C) PI versus FITC (BrdU) Plot av en representant mcf7 celle befolkningen. Gating vises bare å inkludere FITC (BrdU) positive celler (R4).

Figur 2. Representant histogram handlingen i en samlet befolkning på normalt sykling celler.

Figur 3. Histogram tomt av celler som har blitt samlet 1t etter fjerning av BrdU puls, etter gating for FITC (BrdU)-positive celler. BrdU-positive celler i en tidlig endepunktet etter fjerning av etiketten viser PI profiler som tilsvarer prøvene viser DNA-innhold i samsvar med celler som er i S-fasen av cellesyklus, bekrefter vellykket merking av celler bare under DNA-syntese.
 lt = "Figur 4" />
lt = "Figur 4" />
Figur 4. Sammenligning av cellesyklusprogresjon kinetikk mellom kreft cellelinjer, kolorektalkreft HT29 (A) og LS180 (B), samt brystkreft mcf7 (C). Celler ble samlet hver time i 8 timer, etter fjerning av BrdU puls. I dette forsøket, observerte vi en klar profil av fremskyndet cellesyklusprogresjon gjennom G2 fasen av cellesyklus i kolorektal cellelinje LS180. Sammenligning kinetikken mellom de to profilene av kolorektal kreft celler, er fremveksten av en G1 topp tydelig ved T = 4t post-BrdU i LS180 cellene, sammenlignet med tilsvarende tidspunkt for HT-29 cellelinjen som mangler denne toppen. Sammenlignet med ett av de to kolorektal cellelinjer, er mcf7 celler sykling på et betydelig redusert sats. Klikk her for å se større figur .
re 5 "/>
Figur 5. (A) Kvantitativ cellesyklus fase analyse av BrdU-merket kreftceller. Dekan / Jett / Fox algoritme ble brukt til HT29, LS180 og mcf7 (illustrert i grønt). De resulterende cellesyklus fase utdelinger fra hver prøve er uttrykt som% totalt BrdU-positive celler for hver fase. Bare velg tidspunkter for hver celle linje vises, som analysene for noen av de tidligere tidspunkter produsert ugyldige resultater. I disse tidligere tidspunkter, alle BrdU-positive celler er i S-fase og derfor mangler tydelig G1 og G2 topper, som er nødvendige for algoritmen programmet. (B) histogrammer av kreftceller viser kinetikken av progresjon gjennom G2 / M faser av cellesyklus. Progresjon gjennom G2 / M faser er raskest for LS180 celler, etterfulgt av HT29 og mcf7. Klikk her for å se større figur .
jpg "alt =" Figur S1 "/>
Figur S1. Flowcytometri kontroller. mcf7 celler er vist som (A) negativ kontroll der cellene er verken PI eller BrdU beiset. (B) PI bare flekkete. (C) BrdU-FITC merket prøver.

Figur S2. Skjematisk illustrasjon som viser forholdet mellom fluorescerende utslipp spekteret av PI versus FITC (BrdU). De spektrale samling vinduer for fluorescerende kanal 1 (FL1, for FITC) og fluorescerende kanal 3 (FL3, for PI) er vist i tilsvarende bokser. Det er ingen fluoroforen spektra overlapping mellom FL1 og FL3 deteksjon. Åpenbart er fluorescens kompensasjon ikke nødvendig når eksperimentelle data fra PI og FITC-BrdU co-merkede celler samles i FL1/FL3 kanalene hhv. Fluorescens spectrums ble innhentet fra BD Biosciences nettside:target = "_blank"> http://www.bdbiosciences.com/research/multicolor/spectrum_viewer. Klikk her for å se større figur .
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Ved å kombinere flowcytometri med BrdU innlemmelse, har vi de nødvendige verktøy for å studere cellesyklus kinetikk. Den karakteristiske egenskap BrdU å fungere som en tymidin analogt er det som gjør at for DNA innhold kvantifisering av en sykkel celle. Inkorporering av BrdU i et voksende datter DNA-tråden under syntese fasen av cellesyklus er det som gjør en til å følge en undergruppe sykling cellene gjennom replikasjon av DNA i S-fasen, til en periode med vekst på G2 og til slutt til celledeling . Datter BrdU-positive celler kan være kontinuerlig fulgt inntil noe tidspunkt BrdU etiketten er fortynnet ved celledeling i den grad der signalet er redusert til bakgrunnsnivå.
Bruke propidium iodide å bestemme total DNA innhold, er vi i stand til å følge progresjonen i cellesyklus fra S → G2 / M → G1. Muligheten til å spore sykling rate av celler er viktig og nyttig i mange programmer, spesielt FOr pre-kliniske studier med cellesyklus spesifikke DNA-skade induserende medikamenter eller under stoffet utviklingsprosessen for å fastslå cellesyklus stadium følsomhet av romanen forbindelser 11. Den mest åpenbare fordelen med å bruke en metabolsk merking teknikk er eliminering av kjemiske synkronisering av celler. Som de fleste kjemiske behandlinger som induserer cellesyklus er selv genotoksisk, kan dissekere cellesyklus følsomhet effekter av nye forbindelser i nærvær av disse kjemikaliene ikke være gjennomførbart.
Vi viste det allsidigheten til denne BrdU metabolsk merking teknikk med PI co-farging. Ved hjelp av denne protokollen, har vi demonstrert at økningen i cellesyklusprogresjon gjennom G2 / M fasen av cellesyklus kan observeres i telomerase-negative menneskelige celler som har blitt forvandlet til å ha tvunget enzymet uttrykk 10. Selv om denne metoden er kraftig i måle kinetikk i cellen som den sykluser through flere stadier, er en begrensning tap av timing informasjon i løpet av de overgangene, samt innenfor G2 og M faser. Som cellene gå gjennom G2 til M, kan vi ikke skille ikke bare de to fasene, men også den sub-G1 befolkningen, basert utelukkende på DNA innhold. I applikasjoner der dette differensiering er nødvendig, er det potensielt mulig å co-beis BrdU merkede cellene med antistoffer mot sykliner eller andre cellulære proteiner som uttrykk er spesifikke for den ønskede fasen av cellesyklus 12,13. Ytterligere optimalisering vil være nødvendig for å sikre at cyclin antistoffer er kompatible med BrdU etiketten og immuno-farging protokollen. Ideelt sett til tilstrekkelig skille de ulike fasene av cellesyklus, kan flere cyclin antistoffer være nødvendig for å co-etikett en enkelt prøve samtidig. Vårt laboratorium arbeider med post-merking av BrdU-Pulse merkede celler med cyclin B og fosfor histone 3 antistoffer å skille mellom celleri G2 kontra M fasen av cellesyklus. Co-merking med cyclin antistoffer, i tillegg til PI og FITC-BrdU flekker øker vesentlig komplikasjon flowcytometri fra både maskinvare og dataanalyse (fluorescens kompensasjon utgaver) perspektiv. Med ankomsten av den nye generasjonen av flowcytometere, bør fremskritt i sin kapasitet og funksjonalitet lindre disse tekniske bekymringer.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Vi har ingenting å avsløre.
Acknowledgments
Vi takker Andy Johnson av Biomedical Research Center på UBC for hjelp med FACS analyse. Cancer Research Funding i Wong Laboratory er levert av Canadian Cancer Society Research Institute (driftstilskudd # 019250) og fra forskning reinvesteringer midler ved Fakultet for Pharmaceutical Sciences, UBC. JMYW støttes av Canada Research Stoler og Michael Smith Foundation for Health Research Career utviklingsprogrammer.
Materials
| Name | Company | Catalog Number | Comments |
| bromodeoxyuridine | BD Biosciences | 55089 | |
| propidium iodide | Sigma-Aldrich | 287075 | 1mg/ml stock |
| FITC anti-BrdU | BD Biosciences | 347583 | |
| sodium tetraborate | Fisher Scientific | S80172 | 0.1M, pH 8.5 |
| FACS Caliber | BD Biosciences |
References
- Musgrove, E. A., Caldon, C. E., Barraclough, J., Stone, A., Sutherland, R. L. Cyclin D as a therapeutic target in cancer. Nat. Rev. Cancer. 11, 558-572 (2011).
- Molchadsky, A., Rivlin, N., Brosh, R., Rotter, V., Sarig, R. p53 is balancing development, differentiation and de-differentiation to assure cancer prevention. Carcinogenesis. 31, 1501-1508 (2010).
- Dickson, M. A., Schwartz, G. K. Development of cell-cycle inhibitors for cancer therapy. Curr. Oncol. 16, 36-43 (2009).
- Banfalvi, G. Cell cycle synchronization of animal cells and nuclei by centrifugal elutriation. Nat. Protoc. 3, 663-673 (2008).
- van Opstal, A., Boonstra, J. Inhibitors of phosphatidylinositol 3-kinase activity prevent cell cycle progression and induce apoptosis at the M/G1 transition in CHO cells. Cell Mol. Life Sci. 63, 220-228 (2006).
- Gasnereau, I. Flow cytometry to sort mammalian cells in cytokinesis. Cytometry. A. 71, 1-7 (2007).
- Harper, J. V. Synchronization of cell populations in G1/S and G2/M phases of the cell cycle. Methods Mol. Biol. 296, 157-166 (2005).
- Pedrali-Noy, G. Synchronization of HeLa cell cultures by inhibition of DNA polymerase alpha with aphidicolin. Nucleic Acids Res. 8, 377-387 (1980).
- Merrill, G. F. Cell synchronization. Methods Cell Biol. 57, 229-249 (1998).
- Fleisig, H. B., Wong, J. M. Telomerase promotes efficient cell cycle kinetics and confers growth advantage to telomerase-negative transformed human cells. Oncogene. , (2011).
- Cai, D., Byth, K. F., Shapiro, G. I. AZ703, an imidazo[1,2-a]pyridine inhibitor of cyclin-dependent kinases 1 and 2, induces E2F-1-dependent apoptosis enhanced by depletion of cyclin-dependent kinase 9. Cancer Res. 66, 435-444 (2006).
- Pozarowski, P., Darzynkiewicz, Z. Analysis of cell cycle by flow cytometry. Methods Mol. Biol. 281, 301-311 (2004).
- Sherwood, S. W., Rush, D. F., Kung, A. L., Schimke, R. T. Cyclin B1 expression in HeLa S3 cells studied by flow cytometry. Exp. Cell Res. 211, 275-281 (1994).

{kind=link}
{kind=link}