

Summary
שילוב של immunoprecipitation הכרומטין ורצף אולטרה תפוקה גבוהה (שבב ואילך) יכול לזהות ולמפות DNA חלבונים אינטראקציות בקו רקמה או תא נתון. התווה הוא איך ליצור תבנית באיכות גבוהה עבור סידור שבב שלאחר מכן, תוך שימוש בניסיון עם TCF7L2 גורם השעתוק לדוגמא ירושלים.
Abstract
שבב רצף (Chip-אילך) שיטות להציע ישירות כיסוי הגנום כולו, שבו שילוב של הכרומטין immunoprecipitation (שבב) ויכול להיות מנוצל רצף מקביל מסיבי כדי לזהות את הרפרטואר של רצפי DNA יונקים מאוגדים על ידי גורמי שעתוק in vivo. "דור הבא" טכנולוגיות רצף הגנום לספק 1-2 סדרי גודל גידול בסך של רצף שיכול להיות זול ויעיל שנוצר על פני טכנולוגיות ישנות יותר ובכך מאפשר לשיטות שבב seq כדי לספק כיסוי הגנום כולו ישירות לפרופיל יעיל של יונקים אינטראקציות חלבון ה-DNA.
למוצלחים שבב seq גישות, צריך ליצור תבנית ה-DNA שבב באיכות גבוהה כדי להשיג את התוצאות הטובות ביותר רצף. התיאור מבוסס סביב ניסיון עם מוצר החלבון של הגן הכי חזק מעורבים בפתוגנזה של סוכרת מסוג 2, כלומר גורם שעתוק גורם השעתוק דמוי 7 2 (TCF7L2). גורם זה גם היה מעורב בסוגים שונים של סרטן.
התווה הוא איך ליצור תבנית ה-DNA שבב באיכות גבוהה הנגזרת משורת תאי קרצינומה של המעי הגס, HCT116, על מנת לבנות את המפה ברזולוציה גבוהה באמצעות רצף כדי לקבוע את הגנים מאוגדים על ידי TCF7L2, נותן תובנה נוספת לתפקיד המרכזי שלה בפתוגנזה של תכונות מורכבות.
Introduction
במשך שנים רבות יש כבר צורך מסופק לזהות הסט של גנים כפותים ומוסדרים על ידי חלבון נתון הגנום רחב, במיוחד אלה בכיתה גורם שעתוק.
אודום et al. 1 משמש הכרומטין immunoprecipitation (שבב) בשילוב עם microarrays האמרגן לזהות באופן שיטתי את הגנים שנכבשו על ידי רגולטורים תעתיק שהוגדרו מראש בכבד האנושי ואיי לבלב. כתוצאה מכך, ג'ונסון ואח'. 2 פיתחה assay immunoprecipitation הכרומטין בקנה מידה גדולה המבוסס על רצף ישיר אולטרה תפוקה גבוהה DNA (שבב ואילך) על מנת למפות באופן מקיף אינטראקציות-DNA חלבון על פני הגנום כולו יונקים. כמקרה מבחן, הם מיפו in vivo המחייב של הגורם מגביל משתיק נוירון (NRSF) עד 1946 מקומות בגנום האנושי. הנתונים מוצגים ברזולוציה חדה של עמדה מחייבת (+ 50 זוגות בסיסים), שאפשרה גם ליsolation של מוטיבים וזיהוי של מוטיבים NRSF מחייבים. גם היו נתונים שבב seq אלה רגישות גבוהה וספציפיות וביטחון סטטיסטי (p -4 <10), תכונות שחשובות להסקת מסקנות אינטראקציות מועמד חדשה.
רוברטסון et al. 3 משמשים גם שבב ואילך על מנת למפות STAT1 מטרות באינטרפרון-γ (IFN-γ) מגורה וunstimulated תאי S3 הלה אדם in vivo. על ידי שבב ואילך, באמצעות 15.1 ו12.9 מיליון רצף ממופה ייחודי קורא, ושיעור גילוי שקר משוער של פחות מ 0.001, הם זיהו 41,582 11,004 ואזורים STAT1 מחייבים משוערת בתאי מגורה וunstimulated, בהתאמה. מ -34 הלוקוסים ידועים STAT1 4-8 אתרי קישור אינטרפרון מגיבים, שבב אילך מצא 24 (71%). מטרות שבב seq היו מועשרים ברצפים דומים לSTAT1 מוטיבים מחייבים ידועים. השוואות עם שני נתונים שבב ה-PCR קיימים מגדירה הציעושרגישות שבב אילך הייתה בין 70% ו 92% וסגוליות היה 95% לפחות. בנוסף, היה ברור שהשבב ואילך מציע גם מורכבות אנליטית נמוכות ורגישות שעולות עם רצף עומק.
כמו טכנולוגיות רצף הגנום כגון, "הדור הבא" לספק 1-2 סדרי גודל גידול בסך של רצף שיכול להיות זולה ויעיל שנוצר על פני טכנולוגיות ישנות 9. שיטות שבב seq לכן ישירות לספק כיסוי הגנום כולו לאפיון יעיל של אינטראקציות-DNA חלבון 3 יונקים.
בשנת 2006, קשר חזק של גרסאות בגורם השעתוק דמוי 7 2 (TCF7L2) גן עם סוכרת מהסוג 2 התגלה 10. חוקרים אחרים כבר משוכפלים באופן עצמאי את הממצא הזה בקבוצות אתניות שונות ומעניין, מן הגנום מחקרי העמותה הרחב הראשונים של סוכרת מהסוג 2 שפורסמו בכתב עת Nature 11,12 מדע 13-15 ובמקומות אחרים 16,17, הקשר החזק ביותר אכן היה עם TCF7L2, זה נחשב כיום הממצא הגנטי המשמעותי ביותר בסוכרת מהסוג 2 עד כה 18-20. בנוסף, TCF7L2 נקשר לסיכון לסרטן 21,22, ואכן, הקשר הזה היה ברור יותר, כאשר מוקד 8q24 מתגלה על ידי מחקרי עמותה רחבות הגנום של מספר סוגי הסרטן, כולל קרצינומה של מעי גס, הוצג להיות בשל הזרם קיצוני אלמנט TCF7L2 מחייב נהיגה השעתוק של MYC 23,24. ככזה, יש עניין רב בקביעת גנים במורד הזרם מוסדרים על ידי גורם שעתוק זה מפתח.
בהתבסס על ניסיון של עם TCF7L2 כדוגמה למתודולוגיה, מאמר זה מתאר כיצד ליצור תבנית ה-DNA שבב באיכות גבוהה. צ'יפ היה מתבצע בשורת תאי סרטן המעי הגס, HCT116, עבור סידור שלאחר מכן על מנת לבנות גבוה Resolמפת ution מהגנים מאוגדים על ידי TCF7L2 25 במאמץ להניב תובנה נוספת לתפקיד המרכזי שלה בפתוגנזה של תכונות מורכבות.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. הכרומטין חוצה קישור
- לגדל תאים בתרבית תאי 100x20mm מנות. כמות התאים יכולה לנוע בין 1 ל -10 מיליון תאים לכל מנה בהתאם לסוג התא. כ -2 מיליון תאים מספיקים לimmunoprecipitation אחד.
- תאי צולבות קישור בפורמלין 1% למשך 10 דקות בטמפרטורת חדר עם נדנדה מדי פעם.
- להרוות את הצלב מקשר על ידי הוספת ריכוז סופי של 125 מ"מ גליצין ו דגירה במשך 5 דקות בטמפרטורת חדר.
- שטוף תאים עם פוספט 1X שנאגרו מלוח (PBS) פעמיים, למזוג PBS, ולאחר מכן להוסיף 0.2 מ"ל של PBS.
- קציר תאים עם מגרד תא פלסטיק לתוך צינור microcentrifuge.
- ספין למטה תאים ב 2000 סל"ד במשך 5 דקות ב 4 ° C.
- לשאוב supernatant. תאי Resuspend במאגר תמוגה SDS (1% SDS, EDTA 10mm, 50mm טריס-HCl pH 8.1) לכל תא lysate או לשמור אותם כגלולה להפקת נשק גרעיני.
- ניתן לשמור תאים ב -80 ° C, או אחד יכול להמשיך immediatאיליי עם sonication.
2. הכן את הגרעינים (המשך שלב 3.5 לlysate תא השלם)
- להשלים את מאגר תמוגה סלולרי (5 מ"מ pH 8.0 צינורות, 85 מ"מ KCl, 0.5% NP-40) עם 1X אינהיביטור proteinase כל ניסוי.
- Resuspend מופשר תא גלולה בכ 10 פעמים בנפח גלולה עם הצפת תמוגה תא.
- Dounce-Homogenize 10 פעמים עם העלי אז לדגור על קרח במשך 10 דקות.
- צנטריפוגה מדגם ב -4,000 סל"ד במשך 5 דקות ב 4 ° C, להשליך supernatant, ולשמור גלולה גרעינית.
3. Sonication *
- לחמם את מאגר תמוגה SDS וסכום תוספת של חיץ לשימוש עם מעכבי proteinase.
- Resuspend גרעינית גלולה בSDS תמוגה המאגר (כ 0.5 מ"ל של חיץ ל1-10,000,000 תאים)
- לדגור על קרח במשך 10 דקות.
- הוסף 0.5 מ"ל aliquots של דגימות לצינורות microcentrifuge.
- Sonicate על קרח רטוב באמצעות sonicator Misonix עם 30 שניות על 45 שניות וליד הגדרת משרעת של 2. מספר המחזורים לגודל שבר אידיאלי ניתן לקבוע על ידי ראשון שניסה את מספרים שונים (מחזור 2 שמ ', 4, 8, 12, 16, 20 או יותר ומחזורים). מותג של sonicator שונה עשוי לשמש, לעומת זאת, התנאים ישתנו. ניסויים עם מספר המחזורים ואת כמות זמן וכיבוי חייבים להתבצע על מנת לקבוע את התנאים אידיאליים.
- אסוף 20 μl של כל דגימה כדי לבדוק את תוצאות sonication ולעשות כימות,. שאר המדגם ניתן לאחסן ב -80 ° C.
- לדלל את 20 μl של מדגם על ידי הוספת 30 μl של הצפת TE 0.1X.
- פנק את המדגם עם μl 1 של RNase על 37 מעלות צלזיוס במשך שעה 1 ולאחר מכן להוסיף μl 1 של proteinase K ו לדגור על 62 מעלות צלזיוס למשך 2 שעות.
- לרוץ 20 μl של המדגם על 2% ג'ל agarose.
- לטהר את הסכום הנותר של מדגם עם ערכת טיהור PCR QIAquick אז לכמת באמצעות ספקטרופוטומטר NanoDrop.
* לצ'י המקוריP, עיכול nuclease microccocal ניתן להשתמש לחלופין גזירת ה-DNA.
4. חרוזים agarose לחסום *
- אם חרוזים כבר חסומים, המשך לשלב 5.1.
- השתמש בחלבון או חלבון G agarose. במשך 5 immunoprecipitations (IPS), השתמש ב600 μl של 50% חרוז תרחיף (300 μl חרוז גלולה)
- כדי לשטוף את החרוזים, לסובב אותם ב 800 סל"ד דקות 1 על 4 מעלות צלזיוס וזורקים supernatant. הוסף מעט יותר מ -2 חיץ דילול שבב מ"ל (0.01% SDS, EDTA 1.2 מ"מ, 167 מ"מ NaCl, 1.1% טריטון X-100, 16.7 מ"מ טריס-HCl pH 8.1) ומערבב על ידי לאט צינור היפוך 10X. ספין למטה שוב ב800 סל"ד דקות 1 על 4 מעלות צלזיוס וזורקים supernatant. חזור על לשטוף פי 2 יותר.
- חסום את חרוזים על ידי סיבוב על 4 מעלות צלזיוס למשך הלילה בחסימת פתרון. עיין בטבלה 1 למתכון של פתרון החסימה.
5. הכרומטין מראש ברור
- הפשירו sonicated הכרומטין על קרח.
- ספין למטה בסל"ד ו 12,000או 10 דקות ב 4 ° C ואז לשים על קרח מייד כדי להסיר SDS (לבן גלולה).
- איסוף supernatant, להשליך גלולה, ולשלב דוגמאות במידת צורך.
- להוציא את הסכומים דרושים לצורך הניסוי המבוסס על חישובים (1-10 UG של הכרומטין ל-IP).
- לדלל הכרומטין 10X בחיץ דילול שבב בתוספת מעכב proteinase.
- הוסף 100 μl של חרוזים חסומים ל-IP.
- סיבוב על 4 מעלות צלזיוס במשך שעה 1.
6. Immunoprecipitation
- ספין למטה דגימות ב 800 סל"ד דקות 1 ולהעביר את supernatant לצינור טרי.
- ספין למטה supernatant ב 800 סל"ד דקות 1 והעברה לצינור נקי אחר.
- לחסוך 20 μl של supernatant, לשמש כשליטת קלט, ב -20 ° C.
- Aliquot הכרומטין למספר כתובות IP שצריך לעשות בניסוי.
- הוסף 2 UG של נוגדן לUG 1-10 של הכרומטין לכל דגימה.
- דגירה הלילה 4 ° C עם רוטציה.
- הוסף 100 μl של חרוזים חסומים לכל דגימת ה-IP.
- דגירה במשך שעה 1 ב 4 ° C עם רוטציה.
- גלולה החרוזים על ידי שצנח ב 800 סל"ד דקות 1 ולבטל כמה שיותר את supernatant ככל האפשר.
- לשטוף חרוזים פעם עם חיץ לשטוף קומפלקס חיסוני דל מלח. הוסף 1 מ"ל של חיץ לכל צינור, לסובב בטמפרטורת חדר למשך 5-8 דקות, ספין למטה בסל"ד 800 דקות 1, ולאחר מכן להשליך supernatant. חזור על לשטוף פעם עם חיץ גבוה מלח חיסוני קומפלקס לשטוף ולשטוף חיץ קומפלקס חיסוני LiCl ופעמים עם חיץ TE עבור סכום כולל של 5 כביסות (טבלה 2).
7. Elution
- דגימות קלט להפשיר מהיום קודם לכן כדי להיות מעובד עם eluants.
- הפוך Elution הצפת טרי (טבלה 3).
- הפוך את תערובת הורים של ההצפה elution מספיק נחוצה לשב"ס ודגימות בקרת קלט בתוספת 1-2 דוגמאות נוספות.
- הוסף 100 ההצפה elution μl לכל דגימת ה-IP ולדגור על טמפ 'החדר למשך 15 מ'בעם סיבוב.
- ספין למטה בסל"ד 800 דקות 1 ולהוסיף supernatant לצינור טרי.
- הוסף 100 μl נוסף של הצפת elution על צינור אחד של חרוזים ולדגור על טמפ 'החדר במשך 15 דקות עם רוטציה.
- וורטקס במשך 15 שניות לאחר דגירה; ספין למטה ב 5000 סל"ד דקות 1, ולאחר מכן לשלב את supernatant עם supernatant מelution הראשון. (ודא שאין חרוזים שנשארו בsupernatants. אם אינך בטוח, ספין למטה supernatant שוב ב 5000 סל"ד דקות 1 ולאסוף את supernatant בצינור חדש.
- הוסף 180 μl של ההצפה elution ל20 μl של דגימות בקרת קלט.
8. היפוך חוצה קישור
- לμl 200 של eluants ובקרות קלט, להוסיף 8 μl של 5 M NaCl.
- צינורות חותם עם parafilm ודגירה באמבט מים ב 65 ° C במשך הלילה.
9. טיהור ה-DNA
- טיפול בכל דגימה עם μl 1 של RNase עבור שעה 1 ב 37 ° C.
- הוסף 4 μl של 0.5 M EDTA, 8 1M μl טריס-HCl, לערבב, ואז להוסיף μl 1 של proteinase K לכל דגימה ולדגור על 45 מעלות צלזיוס למשך 2 שעות.
- לטהר את הדגימות באמצעות ערכת טיהור PCR QIAquick. ניתן לשמור את הדגימות ב -20 ° C ו-PCR בדיקה יכולה להיעשות במועד מאוחר יותר.
* לחלופין, חרוזים מגנטיים שבב כיתה ניתן להשתמש במקום של agarose עבור חלק immunoprecipitation.
10. סימון ה-PCR
- לבדיקת PCR, השתמש פריימרים לאזורים ידועים להיות מחויבים לחלבון של עניין. כמו כן, השתמש פריימרים לאזורים בלתי מחייבים כביקורת שלילית.
- מערבבים את חומרים כימיים לתגובה. לדלל מדגם קלט ב1:100 (טבלה 3).
- הפעל תגובה. תכנית ה-PCR:
שלב 1: 94 ° C 3 דקות
שלב 2: 94 מעלות צלזיוס 20 שניות
59 ° C 30 שניות
72 ° C 30 שניות
(חזור על שלב 2 עבור לפחות 30 cycles)
שלב 3: 72 מעלות צלזיוס 2 דקות
- הפעל דוגמאות על 1% agarose ג'ל.
- העשרה גם ניתן לקבוע כמותית עם PCR בזמן אמת.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
ברגע שהכרומטין כבר sonicated וטופל עם RNase וproteinase, הדגימות לרוץ על 2% ג'ל agarose צריכה להציג למרוח עם חלק הארי של ה-DNA בגודל הרצוי. אם מספר מחזורים שונים נבדקים, ירידה הדרגתית בגודלו יש לראות במספר מחזורי הגידול (איור 2).
לאחר שסיים את חלק immunoprecipitation של הפרוטוקול או ניתן לבדוק את ההעשרה על ידי PCR או PCR בזמן אמת. לקבלת דוגמיות PCR לרוץ על ג'ל agarose צריכה להיות בלהקות הקלט והשבב (באמצעות נוגדן לחלבון של עניין, שהוא TCF7L2 במקרה זה) לדוגמה נתיבים ולא שום דבר או לכל היותר, להקה קלושה מאוד (רעש רקע) ב נתיב ביקורת (שלילי) IgG לאזור המחייב החיובי. לאזור המחייב השלילי לא צריכה להיות מאוד קלוש, אם בכלל, להקה לשליטה IgG ונתיבי שבב. לא צריך להיות להקה בנתיב הקלט (איור 3).
איור 4 מראה את אותן דגימות שנבדקו על ידי בזמן אמת PCR. כמו עם הדמות הקודמת, לא אמורה להיות העשרת קיפול משמעותית של האזור מחייב החיובי לדוגמא השבב על שליטת IgG. כמו כן, לא אמורה להיות העשרה מעט מאוד, אם בכלל, ראתה באזור מחייב השלילי.

איור 1. תרשים זרימה של תהליך שבב. לחץ כאן לצפייה בדמות גדולה.
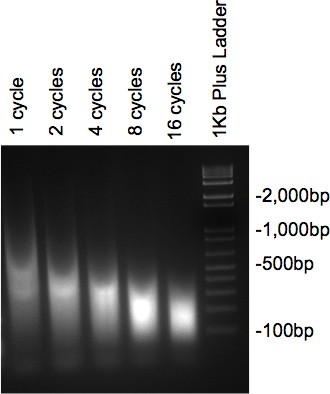
איור 2. ג'ל לבדוק sonication של ה-DNA.

איור 3. בדוק PCR של שבב.
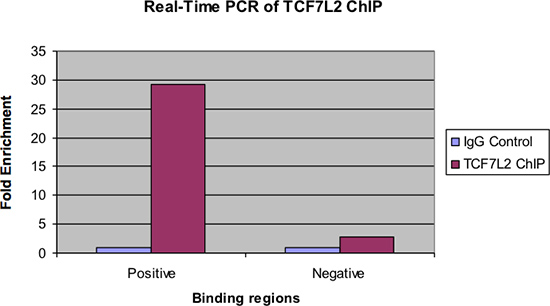
איור 4. Real-Time PCR של TCF7L2 שבב.
| מגיב | נפח |
| חרוז גלולה | 300 μl |
| ארגון ה-BSA (50 מ"ג / מ"ל) | 30 μl |
| אינהיביטור proteinase 100X | 10 μl |
| חיץ דילול שבב | 660 μl |
| סך הכל | 1,000 μl |
טבלת 1. מתכון לחסימת agarose.
| חוצץ | רכיבים |
| הצפת לשטוף קומפלקס חיסוני דלת מלח | 0.1% SDS 1% טריטון X-100 2 מ"מ EDTA 20 מ"מ טריס-HCl pH 8.1 150 מ"מ NaCl |
| הצפת לשטוף קומפלקס חיסוני מלח גבוה | 0.1% SDS 1% טריטון X-100 2 מ"מ EDTA 20 מ"מ טריס-HCl pH 8.1 500 מ"מ NaCl |
| הצפת לשטוף קומפלקס חיסוני LiCl | 0.25 מ 'LiCl 1% NP-40 1% Deoxycholate 1 mM EDTA 10 מ"מ טריס-HCl pH 8.1 |
| TE חיץ | 10 מ"מ טריס-HCl pH 8.1 1 mM EDTA pH 8.0 |
טבלת 2. מאגרים לשטוף שבב.
| מגיב | נפח |
| 10 μl | |
| 1 M NaHCO 3 | 20 μl |
| H 2 O | 170 μl |
לוח 3. מאגר elution לIP אחת.
| מגיב | 50 תגובת μl | 20 תגובת μl |
| מים | 27 μl | 10.8 μl |
| מאגר תגובת PCR 5X | 10 μl | 4 μl |
| MgCl 2 | 4 μl | 1.6 μl |
| dNTP (10 מ"מ) | μl 1 | 0.4 μl |
| לערבב פריימר (5 אום כל אחד) | 2 μl | 0.8 μl |
| תקי (Promega Hotstart) | <Μl 1/ P> | 0.4 μl |
| ה-DNA השבב | 5 μl | 2 μl |
לוח 4. כרכי תגובת PCR.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
עכשיו זה אפשרי לבצע את פרופיל הגנום של עמותת אינטראקציות חלבון דנ"א באמצעות שבב ואילך, כפי שהודגם לאחרונה עם גורמי שעתוק אחרים 2,3. המפתח לתוצאה מוצלחת הוא רצף של דור תבנית immunoprecipitation הכרומטין באיכות גבוהה ה-DNA.
ברגע שתבנית ה-DNA כבר נוצר והוברר להיות מועשרים במידה מספקת, ואז אחד יכול לקחת את זה בספריית הכנה עבור סידור שלאחר מכן. לדוגמה, ניתן להשתמש בפרוטוקול ספריית הרצף מסופק על ידי הספק, Illumina. בחירת הגודל של ספרייה זו יכולה להתבצע על ידי ג'ל אלקטרופורזה וכריתה שלאחר מכן וטיהור של ה-DNA ב~ 200 - לטווח של 700 נ"ב. הקטנת הגודל וצמצום טווח הגודל של DNAs שנאסף מטיהור ג'ל נועד לשפר את הרזולוציה של מיקומית שבב ואילך. על ידי העשרה לחתיכות קטנות יותר של דנ"א הקלט מאוגד לגורם של interest, ניתן היה לצפות שמיקום האתר יקבל החלטה. בחירת גודל הדוקה יותר גם משפרת את אחידות הגודל של מושבות מולקולריות המיוצרות על פלטפורמת Illumina. אחידות גודל מושבה כזו גם מגדילה את מספר הקריאה היעיל שהושג. גודל ה-DNA קלט קצר יותר גם מייצר מושבות חזקות יותר על פלטפורמת Illumina, וזה כנראה אומר שחלקי DNA קצרים יותר בתוך כל חלוקה מדגם קלט נתון יהיו מיוצגים בצורה יעילה יותר ברצף הפלט הסופי הן קלט מחתיכות ארוכות יותר מאותה ההתפלגות.
הגישות לניתוח bioinformatic רצף "דור הבא" ממשיכות להתפתח, עם ספקים רבים שהופכים את התוכנה שלהם בקוד פתוח לחידוד נוסף. אפשר לשנות את המפה שקוראה למקומות גנומית ייחודיים לתוך פרופיל חפיפה קטע DNA. ניתן לזהות על ידי פרופילי threshholding פסגות משמעותיות בגובה שווה ערך לשיעור גילוי משוער שווא. SP העמדהמטריצות תדר ecific הנגזרות מעבודה זו יכולות לשמש כדי לזהות ולמקם אתרי קישור DNA בגנום האנושי לגורם נתון.
אבל צריך להיות זהיר לגבי מה הגורם אחד רוצה ללמוד עם שבב ואילך. לפני היציאה למחקר אחד כזה צריך להעריך אם נוגדן הוא זמין בשוק כי הוא שמיש בהגדרת השבב, כנוגדן עני יכול להיות השפעות מזיקות על אחד מאוד של תוצאות ניסוי. בנוסף, יש לשקול אם יש isoforms האחו של החלבון הנחקר, ואכן, TCF7L2 ידוע יש isoforms רבים כל כך שהיינו זהירים במיוחד בבחירת נוגדן שקשר לחומצות אמינו בהווה באופן עקבי בכל isoforms העיקרי של גורם שעתוק זה 25.
לסיכום, שילוב של immunoprecipitation הכרומטין ורצף אולטרה תפוקה גבוהה (שבב ואילך) יכול לזהות ולמפות אינטראקציות-DNA חלבון ברקמה מסוימת או גקו ell. נציין כאן איך ליצור תבנית שבב באיכות גבוהה עבור סידור שלאחר מכן.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
החוקרים מצהירים כי אין להם אינטרסים כלכליים מתחרים.
Acknowledgments
העבודה נתמכת על ידי פרס מכון לפיתוח מבית החולים לילדים של פילדלפיה.
Materials
| Name | Company | Catalog Number | Comments |
| QIAquick PCR Purification Kit | Qiagen | 28104 | |
| EZ-ChIP Kit | Millipore | 17-371 | |
| GoTaq Hot Start Polymerase | Promega | M5001 | |
| Misonix Sonicator | Qsonica | XL-2000 | |
| NanoDrop 1000 Spectrophotometer | Thermo-Scientific | ||
| Positive control primer sequences (TCF7L2-1) Forward- 5'-TCGCCCTGTCAATAATCTCC-3' Reverse- 5'-GCTCACCTCCTGTATCTTCG-3' Negative control primer sequences (CTRL-1) Forward-5'-ATGTGGTGTGGCTGTGATGGGAAC-3' Reverse- 5'-CGAGCAATCGGTAAATAGGTCTGG-3' |
|||
References
- Odom, D. T., et al. Control of pancreas and liver gene expression by HNF transcription factors. Science. 303, 1378-1381 (2004).
- Johnson, D. S., Mortazavi, A., Myers, R. M., Wold, B. Genome-wide mapping of in vivo protein-DNA interactions. Science. 316, 1497-1502 (2007).
- Robertson, G., et al. Genome-wide profiles of STAT1 DNA association using chromatin immunoprecipitation and massively parallel sequencing. Nature Methods. 4, 651-657 (2007).
- Reich, N. C., Liu, L.
- Lodige, I., et al. Nuclear export determines the cytokine sensitivity of STAT transcription factors. The Journal of Biological Chemistry. 280, 43087-43099 (2005).
- Schroder, K., Sweet, M. J., Hume, D. A. Signal integration between IFNgamma and TLR signalling pathways in macrophages. Immunobiology. 211, 511-524 (2006).
- Vinkemeier, U. Getting the message across, STAT! Design principles of a molecular signaling circuit. The Journal of Cell Biology. 167, 197-201 (2004).
- Brierley, M. M., Fish, E. N.
- Bentley, D. R.
- Grant, S. F., et al. Variant of transcription factor 7-like 2 (TCF7L2) gene confers risk of type 2 diabetes. Nature Genetics. 38, 320-323 (2006).
- Sladek, R., et al. A genome-wide association study identifies novel risk loci for type 2 diabetes. Nature. 445, 881-885 (2007).
- Wellcome Trust Case Control Consortium. Genome-wide association study of 14,000 cases of seven common diseases and 3,000 shared controls. Nature. 447, 661-678 (2007).
- Saxena, R., et al. Genome-wide association analysis identifies loci for type 2 diabetes and triglyceride levels. Science. 316, 1331-1336 (2007).
- Zeggini, E., et al. Replication of genome-wide association signals in UK samples reveals risk loci for type 2 diabetes. Science. 316, 1336-1341 (2007).
- Scott, L. J., et al. A genome-wide association study of type 2 diabetes in Finns detects multiple susceptibility variants. Science. 316, 1341-1345 (2007).
- Steinthorsdottir, V., et al. A variant in CDKAL1 influences insulin response and risk of type 2 diabetes. Nature Genetics. 39, 770-775 (2007).
- Salonen, J. T., et al. Type 2 Diabetes Whole-Genome Association Study in Four Populations: The DiaGen Consortium. American Journal of Human Genetics. 81, 338-345 (2007).
- Zeggini, E., McCarthy, M. I. TCF7L2: the biggest story in diabetes genetics since HLA. Diabetologia. 50, 1-4 (2007).
- Weedon, M. N.
- Hattersley, A. T. Prime suspect: the TCF7L2 gene and type 2 diabetes risk. The Journal of Clinical Investigation. 117, 2077-2079 (2007).
- Yochum, G. S., et al. Serial analysis of chromatin occupancy identifies beta-catenin target genes in colorectal carcinoma cells. Proceedings of the National Academy of Sciences of the United States of America. 104, 3324-3329 (2007).
- Duval, A., Busson-Leconiat, M., Berger, R., Hamelin, R. Assignment of the TCF-4 gene (TCF7L2) to human chromosome band 10q25.3. Cytogenet. Cell Genet. 88, 264-265 (2000).
- Pomerantz, M. M., et al. The 8q24 cancer risk variant rs6983267 shows long-range interaction with MYC in colorectal cancer. Nature Genetics. 41, 882-884 (2009).
- Tuupanen, S., et al. The common colorectal cancer predisposition SNP rs6983267 at chromosome 8q24 confers potential to enhanced Wnt signaling. Nature Genetics. 41, 885-890 (2009).
- Zhao, J., Schug, J., Li, M., Kaestner, K. H., Grant, S. F. Disease-associated loci are significantly over-represented among genes bound by transcription factor 7-like 2 (TCF7L2) in vivo. Diabetologia. 53, 2340-2346 (2010).
- Benjamini, Y., Yekutieli, D. Quantitative trait Loci analysis using the false discovery rate. Genetics. 171, 783-790 (2005).
