

Summary
Kombinationen af kromatin immunopræcipitation og ultra-high-throughput sekventering (Chip-seq) kan identificere og kortlægge protein-DNA interaktioner i en given vævs-eller cellelinie. Skitseret, er hvordan man kan generere en høj kvalitet chip skabelon til efterfølgende sekventering, ved hjælp erfaring med transkriptionsfaktor TCF7L2 som et eksempel.
Abstract
Chip-sekventering (Chip-seq) metoder direkte tilbyde hel-genom dækning, hvor kombinerer kromatin immunfældning (chip) og massivt parallelle sekventering kan udnyttes til at identificere repertoire af mammale DNA-sekvenser bundet af transkriptionsfaktorer in vivo. "Næste generation" genomsekvensering teknologier giver 1-2 størrelsesordener stigning i mængden af sekvensen, der kan være omkostningseffektivt genereret over ældre teknologier og dermed giver mulighed for chip-seq metoder til direkte at give hel-genom dækning for en effektiv profilering af pattedyr protein-DNA interaktioner.
For vellykkede chip-seq tilgange, må man generere høj kvalitet chip DNA skabelon til at opnå de bedste sekventering resultater. Beskrivelsen er baseret omkring erfaring med proteinet af genet stærkest impliceret i patogenesen af type 2-diabetes, nemlig transkriptionsfaktor transkriptionsfaktor 7-lignende 2 (TCF7L2). Denne faktor er også blevet impliceret i forskellige cancerformer.
Skitseret, er hvordan man kan generere høj kvalitet chip DNA-skabelon stammer fra kolorektal carcinom cellelinie HCT116, for at opbygge en høj opløsning kort ved sekventering til at bestemme de gener der er bundet af TCF7L2, hvilket giver yderligere indsigt i sin centrale rolle i patogenesen af komplekse egenskaber.
Introduction
I mange år har der været et udækket behov for at identificere det sæt af gener bundet og reguleret af et givet protein genom, navnlig i transkriptionsfaktoren klassen.
Odom et al. 1. brugte kromatin immunpræcipitering (chip) kombineret med promoter microarrays på systematisk at kortlægge de gener besat af forud specificerede transskriptionsregulatorer i menneskets lever og pancreas holme. Efterfølgende udviklede Johnson et al. 2 et storstilet kromatin immunfældning assay baseret på direkte ultra high-throughput DNA-sekventering (Chip-seq) for på udtømmende kortlægge protein-DNA interaktioner tværs hele pattedyrs genomer. Som en prøvesag, kortlagt de in vivo bindingen af neuron-begrænsende lyddæmper faktor (NRSF) til 1946 steder i det humane genom. De data viste skarp opløsning af bindende position (+ 50 basepar), som gjorde det lettere både isolation af motiver og identifikation af NRSF-bindende motiver. Disse chip-seq data havde også høj følsomhed og specificitet og statistisk selvtillid (P <10 -4), egenskaber, som er vigtige for at udlede ny kandidat interaktioner.
Robertson et al. 3 anvendes også chip-seq for at kortlægge Stat1 mål i interferon-γ (IFN-γ)-stimulerede og ustimulerede humane HeLa S3-celler in vivo. Af Chip-seq, ved hjælp 15,1 og 12,9 mio entydigt kortlagt sekvens læser, og en estimeret falsk opdagelse på mindre end 0,001, de identificerede 41.582 og 11.004 formodede STAT1-bindende regioner stimulerede og ustimulerede celler. Af de 34 loci vides at indeholde STAT1 interferon-responsiv bindingssteder 4-8, fandt chip-seq 24 (71%). Chip-seq mål blev beriget i sekvenser svarende til kendte Stat1 bindende motiver. Sammenligninger med to eksisterende chip-PCR datasæt foreslåetden chip-seq sensitivitet var mellem 70% og 92% og specificitet var mindst 95%. Derudover var det tydeligt, at chip-seq tilbyder både lav analytisk kompleksitet og følsomhed, der stiger med sekventering dybde.
Som sådan "næste generation" genomsekvensering teknologier giver 1-2 størrelsesordener stigning i mængden af sekvensen, der kan være omkostningseffektivt genereret over ældre teknologier 9. Chip-seq metoder derfor direkte give hel-genom dækning for en effektiv profilering af mammale protein-DNA interaktioner 3.
I 2006 blev en stærk sammenslutning af varianter i transskription faktor 7-lignende 2 (TCF7L2) genet med type 2-diabetes opdaget 10.. Andre forskere har allerede uafhængigt gentaget denne konstatering i forskellige etniske grupper, og interessant fra de første genom dækkende forening undersøgelser af type 2-diabetes i Nature 11,12 Science 13-15 og andre steder 16,17, var den stærkeste forening faktisk med TCF7L2, og dette er nu betragtes som den mest betydningsfulde genetiske fund i type 2-diabetes til dato 18-20. Desuden har TCF7L2 været forbundet med risiko for kræft 21,22, ja, denne forbindelse blev mere indlysende, når den 8q24 locus afsløret af genom dækkende forening undersøgelser af en række kræftformer, herunder kolorektale carcinomer, blev vist at skyldes en ekstrem opstrøms TCF7L2-bindende element kørsel transskription af MYC 23,24. Som sådan er der stor interesse i at fastlægge de nedstrøms gener reguleres af denne nøgle transskription faktor.
Baseret på erfaringer med TCF7L2 som et eksempel på den metode, skitserer dette papir, hvordan at generere høj kvalitet chip DNA-skabelon. Chip blev udført i kolorektal carcinom cellelinie HCT116, til efterfølgende sekventering på at opbygge en høj-resolution kort over de gener der er bundet af TCF7L2 25 i et forsøg på at frembringe yderligere indsigt i sin centrale rolle i patogenesen af komplekse egenskaber.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. Cross-link Chromatin
- Grow celler i 100x20mm celle dyrkningsskåle. Mængden af celler kan variere fra 1 til 10 millioner celler pr skålen afhængig celletype. Ca. 2 millioner celler er tilstrækkeligt til en immunfældning.
- Cross-link celler i 1% formaldehyd i 10 minutter ved stuetemperatur med lejlighedsvis rokkende.
- Quench tværbinding ved tilsætning af en slutkoncentration på 125 mM glycin og inkuberes i 5 minutter ved stuetemperatur.
- Vask cellerne med 1X phosphatpufret saltvand (PBS) to gange dekanteres PBS, og derefter tilsættes 0,2 ml PBS.
- Høst celler med en plastik celleskraber i et mikrocentrifugerør.
- Spin ned cellerne ved 2.000 rpm i 5 minutter ved 4 ° C.
- Aspirere supernatanten. Resuspender celler i SDS Lysis buffer (1% SDS, 10 mM EDTA, 50 mM Tris-HCI pH 8,1) til helcellelysat eller holde dem som en pellet for nuklear ekstraktion.
- Celler kan gemmes ved -80 ° C eller man kan fortsætte immediately med sonikering.
2. Forbered Kerner (Fortsæt til Trin 3.5 for helcellelysat)
- Supplement Cell Lysis Buffer (5 mM PIPES pH 8,0, 85 mM KCI, 0,5% NP-40) med 1X proteinaseinhibitor hvert eksperiment.
- Resuspenderes optøet cellebundfaldet på cirka 10 gange den pellet volumen med Cell Lysis Buffer.
- Dounce-homogenisere 10 gange med pistil derefter inkuberes på is i 10 min.
- Centrifuger prøven ved 4.000 rpm i 5 minutter ved 4 ° C, kasseres supernatanten, og gemme kernepellet.
3. Sonikering *
- Varm op SDS Lysis Buffer og ekstra mængde puffer, der skal anvendes med proteinaseinhibitor.
- Resuspender kernepellet i SDS Lysis Buffer (ca. 0,5 ml puffer pr 1-10.000.000 celler)
- Inkuber på is i 10 min.
- Tilsæt 0,5 ml alikvoter af prøver til mikrocentrifugerør.
- Sonikeres på våd is hjælp Misonix sonikator med 30 sek på og 45 sekunderoff på en amplitude indstilling på 2. Antal cykler til ideal fragment størrelse kan bestemmes ved først afprøve forskellige cyklus numre (ex. 2, 4, 8, 12, 16 og 20 eller flere cykler). Et andet mærke sonikatoren kan anvendes, dog vil de betingelser varierer. Eksperimenter med antallet af cyklusser og mængden af tid og slukkes skal udføres for at bestemme de ideelle betingelser.
- Saml 20 ul af hver prøve for at kontrollere lydbehandling resultater og gøre kvantificering. Resten af prøven kan opbevares ved -80 ° C.
- Fortyndes 20 ul prøve ved at tilsætte 30 pi 0,1 X TE buffer.
- Behandle prøven med 1 ml af RNase A ved 37 ° C i 1 time og derefter tilsættes 1 ml af proteinase K og inkuberes ved 62 ° C i 2 timer.
- Kør 20 ul af prøven på en 2% agarosegel.
- Rense den resterende mængde prøve med QIAquick PCR Oprensning kit derefter kvantificere bruge NanoDrop spektrofotometer.
* For indfødte CHIP, kan microccocal nuclease fordøjelse anvendes alternativt at forskyde DNA'et.
4.. Block agaroseperler *
- Hvis perler allerede er blokeret, skal du fortsætte til trin 5.1.
- Brug Protein A eller protein G agarose. For 5 immunopræcipitationer (IP), skal du bruge 600 ml 50% perle gylle (300 gl kuglepelleten)
- At vaske perlerne, spinde dem ned ved 800 rpm i 1 min ved 4 ° C og kasseres supernatanten. Tilføj lidt mere end 2 ml chip Fortyndingsbuffer (0,01% SDS, 1,2 mM EDTA, 167 mM NaCl, 1,1% Triton X-100, 16,7 mM Tris-HCI pH 8,1) og blandes ved langsomt at vende røret 10X. Spin ned igen ved 800 rpm i 1 min ved 4 ° C og kasseres supernatanten. Gentag vask 2 gange mere.
- Bloker perler ved at dreje ved 4 ° C natten over i blokerende opløsning. Se Tabel 1 for opskrift blokerende opløsning.
5.. Pre-klar Chromatin
- Thaw lydbehandlet kromatin på is.
- Spin ned ved 12.000 rpm feller 10 min ved 4 ° C og derefter sat på is straks at fjerne SDS (hvid pellet).
- Saml supernatanten kasseres pellet, og kombinere prøverne, hvis nødvendigt.
- Tag de beløb, der er nødvendige for eksperimentet baseret på beregninger (1-10 ug kromatin per IP).
- Fortynd kromatin 10X chip fortyndingsbuffer suppleret med proteinaseinhibitor.
- Tilføj 100 ul blokerede perler pr IP.
- Rotere ved 4 ° C i 1 time.
6.. Immunpræcipitation
- Spin ned prøver ved 800 rpm i 1 min og overføre supernatanten til et frisk rør.
- Spin ned supernatant ved 800 rpm i 1 min og overførsel til et andet rent glas.
- Gem 20 ul af supernatanten, at tjene som indgangs kontrol, ved -20 ° C.
- Alikvot kromatin til antallet af IP-adresser, der skal gøres i eksperimentet.
- Tilsæt 2 ug af antistof pr 1-10 ug kromatin til hver prøve.
- Inkuber natten over 4 ° C med rotation.
- Tilføj 100 ul blokerede perler til hver IP prøve.
- Der inkuberes i 1 time ved 4 ° C med rotation.
- Pelletere perlerne ved spinding ned ved 800 rpm i 1 min og bortskaffe så meget af supernatanten som muligt.
- Vask perlerne en gang med lav Salt immunkompleks vaskebuffer. Tilsæt 1 ml buffer til hvert rør, roterer ved stuetemperatur i 5-8 min, spin ned ved 800 rpm i 1 min, og derefter kassere supernatanten. Gentag vask en gang med High Salt immunkompleks vaskebuffer og LiCl immunkompleks vaskebuffer og to gange med TE buffer til i alt 5 vaske (tabel 2).
7.. Eluering
- Thaw input prøver fra foregående dag skal behandles med elueringsmidler.
- Gør Elution Buffer frisk (tabel 3).
- Lav en master blanding af nok Elution Buffer nødvendig for IP og input kontrolprøver plus 1-2 ekstra prøver.
- Tilsæt 100 pi elueringspuffer til hver IP prøve og inkuber ved stuetemperatur i 15 mi med rotation.
- Spin ned ved 800 rpm i 1 min og tilføje supernatanten til et frisk rør.
- Tilføje en anden 100 ul elueringspuffer til hvert rør af perler og inkuber ved stuetemperatur i 15 min med rotation.
- Vortex i 15 sekunder efter inkubation spin ned ved 5000 rpm i 1 minut, derefter kombinere supernatanten med supernatanten fra den første eluering. (Sørg for at der er nogen tilovers perler i supernatanterne. Hvis du er usikker, spin ned supernatant igen ved 5.000 rpm i 1 min og indsamle supernatanten i et nyt rør.
- Tilføj 180 ul Elution Buffer til 20 ul af input kontrolprøver.
8. Reverse Cross-link
- Til 200 pi eluanter og input kontrol, 8 pi 5 M NaCl tilføje.
- Tætningsrørene med parafilm og inkuberes i vandbad ved 65 ° C natten over.
9.. DNA-oprensning
- Behandl hver prøve med 1 pi RNase A i 1 time ved 37 ° C.
- Tilsæt 4 μl 0,5 M EDTA, 8 pi 1M Tris-HCI, blandes, hvorefter der tilsættes 1 ml af proteinase K til hver prøve og inkuberes ved 45 ° C i 2 timer.
- Renses prøverne ved anvendelse QIAquick PCR Purification kit. Prøverne kan gemmes ved -20 ° C og PCR-kontrol kan gøres på et senere tidspunkt.
* Alternativt kan chip-grade magnetiske perler kan anvendes i stedet for agarose til immunopræcipitation del.
10. PCR kontrol
- Til PCR check, bruge primere for regioner vides at være bundet af proteinet af interesse. Også bruge primere til ikke-bindende regioner som negative kontroller.
- Bland reagenserne til reaktionen. Fortyndet Input prøve på 1:100 (tabel 3).
- Kør reaktion. PCR Program:
Trin 1: 94 ° C 3 min
Trin 2: 94 ° C 20 sek
59 ° C 30 sek
72 ° C 30 sek
(Gentag trin 2 for mindst 30 CYCLes)
Trin 3: 72 ° C 2 min
- Køre prøver på 1% agarosegel.
- Berigelse kan også bestemmes kvantitativt med real-time PCR.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Når kromatin er sonikeret og er blevet behandlet med RNase og proteinase bør prøverne kører på 2% agarosegel præsentere en overhældes med hovedparten af DNA ved den ønskede størrelse. Hvis flere forskellige cyklusser testes, bør en gradvis nedgang i størrelse ses som antallet af cyklusser stigning (figur 2).
Efter endt immunofældning del af protokollen berigelsen kan enten kontrolleres ved PCR eller real-time PCR. For PCR prøver kørt på en agarose gel bør der være bands i Input og chip (ved hjælp af antistof til protein af interesse, hvilket er TCF7L2 i dette tilfælde) prøve baner og intet eller højst en meget svag band (baggrundsstøj) i IgG (negativ) kontrol lane for den positive bindende region. For den negative bindende region bør der være meget svag eller ingen bånd for IgG kontrol og chip baner. Der bør være et band i Input Lane (Figur 3).
Figur 4 viser de samme prøver undersøgt af real-time PCR. Som med den foregående figur, bør der være en betydelig fold berigelse af positiv binding region for chip prøve gennem IgG kontrol. Desuden bør der være meget lidt berigelse, hvis nogen, set i den negative bindende region.

Figur 1.. Flow diagram chip proces. Klik her for at se større figur .
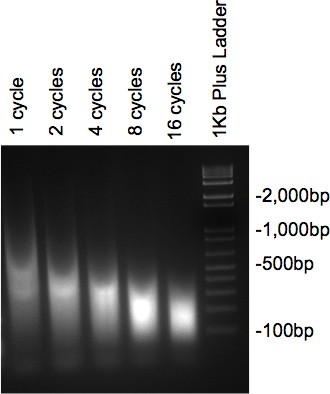
Figur 2. Gel kontrollere af DNA lydbehandling.

Figur 3. PCR Kontrol af chip.
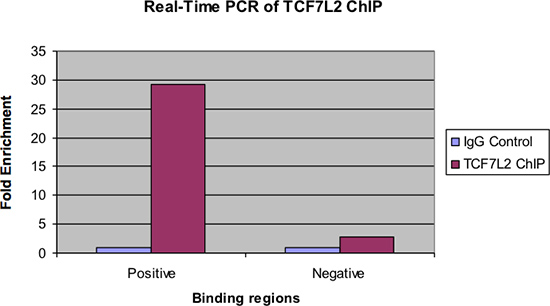
Figur 4.. Real-time PCR af TCF7L2 chip.
| Reagens | Volumen |
| Kuglepelleten | 300 gl |
| BSA (50 mg / ml) | 30 ul |
| 100X proteinaseinhibitor | 10 ul |
| Chip fortyndingsbuffer | 660 pi |
| Total | 1.000 pi |
Tabel 1. Opskrift til at blokere agarose.
| Buffer | Komponenter |
| Lav Salt immunkompleks vaskebuffer | 0,1% SDS 1% Triton X-100 2 mM EDTA 20 mM Tris-HCI pH 8,1 150 mM NaCI |
| High Salt immunkompleks vaskebuffer | 0,1% SDS 1% Triton X-100 2 mM EDTA 20 mM Tris-HCI pH 8,1 500 mM NaCI |
| LiCI immunkompleks vaskebuffer | 0,25 M LiCI 1% NP-40 1% deoxycholat 1 mM EDTA 10 mM Tris-HCI pH 8,1 |
| TE Buffer | 10 mM Tris-HCI pH 8,1 1 mM EDTA, pH 8,0 |
Tabel 2. CHIP vaskebuffere.
| Reagens | Volumen |
| 10 ul | |
| 1 M NaHCO3 | 20 pi |
| H2O | 170 pi |
Tabel 3. Elueringsbufferen for en IP.
| Reagens | 50 ul reaktion | 20 ul reaktion |
| Vand | 27 pi | 10,8 pi |
| 5X PCR reaktionsbuffer | 10 ul | 4 gl |
| MgCl2 | 4 gl | 1,6 gl |
| dNTP (10 mM) | 1 pi | 0,4 gl |
| Primer mix (5 uM hver) | 2 pi | 0,8 gl |
| Taq (Promega Hotstart) | 1 pi </ Td> | 0,4 gl |
| Chip DNA | 5 pi | 2 pi |
Tabel 4. PCR-reaktionsbetingelser mængder.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Det er nu muligt at foretage en genom-dækkende profil af protein-DNA interaktioner forening med chip-seq, som har været meget nylig demonstreret med andre transkriptionsfaktorer 2,3. Nøglen til en vellykket sekventering resultat er dannelsen af en høj kvalitet kromatin immunopræcipitation DNA-skabelon.
Når DNA-skabelonen er blevet genereret, og konstateret at være tilstrækkeligt beriget, kan man derefter tage det i biblioteket forberedelse til efterfølgende sekventering. For eksempel kan man bruge sekventering biblioteket protokol, der af sælger, Illumina. Størrelsen udvælgelsen af dette bibliotek kan udføres ved gelelektroforese og efterfølgende udskæring og oprensning af DNA i ~ 200 - til 700-bp rækkevidde. Reducere størrelsen og indsnævre størrelsesområdet DNA'er indsamlet fra geloprensning skal forbedre positionel opløsning chip-seq. Ved berigelse for mindre stykker indført DNA bundet til faktor intpåløbne, ville man forvente, at webstedet placering vil vinde opløsning. Strammere størrelse udvælgelse forbedrer også størrelsen ensartethed af molekylære kolonier produceres på Illumina platformen. Sådan koloni størrelse ensartethed øger også den effektive læse opnåede tal. Kortere input DNA størrelse producerer også mere robuste kolonier på Illumina platform, og det kan betyde, at kortere DNA stykker inden for et givet input prøve fordeling vil være repræsenteret mere effektivt i den endelige rækkefølge produktion end er længere input stykker fra samme fordeling.
De bioinformatiske tilgange til "næste generation" sekvensanalyse fortsætter med at udvikle sig, med mange leverandører gør deres software open source for yderligere raffinement. Man kan omdanne læser dette kort til unikke genomiske placeringer i et DNA-fragment overlapper profil. Betydende toppe kan identificeres ved threshholding profiler i en højde svarende til en anslået falsk opdagelse sats. Positionen specific frekvens matricer afledt fra dette arbejde kan anvendes til at identificere og lokalisere DNA bindingssteder over den humane genom for en given faktor.
Men man skal være forsigtig med hensyn til, hvilke faktorer man ønsker at studere med chip-seq. Før der iværksættes på en sådan undersøgelse bør man vurdere, om et antistof er til rådighed på markedet, der er anvendelig på chippen indstilling, som en fattig antistof kan have meget skadelige virkninger på ens eksperimentelle resultater. Desuden bør man overveje, om der er splejsningsisoformer af proteinet under undersøgelse, tværtimod er TCF7L2 kendt for at have mange isoformer så vi var særlig forsigtig med at vælge et antistof, som er bundet til aminosyrer konsekvent til stede i alle de vigtigste isoformer af denne transkriptionsfaktor 25..
Sammenfattende kan kombinationen af kromatin immunpræcipitering og ultra-high-throughput sekventering (Chip-seq) identificere og kortlægge protein-DNA interaktioner i en given væv eller cell linje. Vi har skitseret hvordan man kan generere en høj kvalitet chip skabelon til efterfølgende sekventering.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Forfatterne erklærer, at de ikke har nogen konkurrerende finansielle interesser.
Acknowledgments
Arbejdet er støttet af et institut Development Award fra Børnehospitalet i Philadelphia.
Materials
| Name | Company | Catalog Number | Comments |
| QIAquick PCR Purification Kit | Qiagen | 28104 | |
| EZ-ChIP Kit | Millipore | 17-371 | |
| GoTaq Hot Start Polymerase | Promega | M5001 | |
| Misonix Sonicator | Qsonica | XL-2000 | |
| NanoDrop 1000 Spectrophotometer | Thermo-Scientific | ||
| Positive control primer sequences (TCF7L2-1) Forward- 5'-TCGCCCTGTCAATAATCTCC-3' Reverse- 5'-GCTCACCTCCTGTATCTTCG-3' Negative control primer sequences (CTRL-1) Forward-5'-ATGTGGTGTGGCTGTGATGGGAAC-3' Reverse- 5'-CGAGCAATCGGTAAATAGGTCTGG-3' |
|||
References
- Odom, D. T., et al. Control of pancreas and liver gene expression by HNF transcription factors. Science. 303, 1378-1381 (2004).
- Johnson, D. S., Mortazavi, A., Myers, R. M., Wold, B. Genome-wide mapping of in vivo protein-DNA interactions. Science. 316, 1497-1502 (2007).
- Robertson, G., et al. Genome-wide profiles of STAT1 DNA association using chromatin immunoprecipitation and massively parallel sequencing. Nature Methods. 4, 651-657 (2007).
- Reich, N. C., Liu, L.
- Lodige, I., et al. Nuclear export determines the cytokine sensitivity of STAT transcription factors. The Journal of Biological Chemistry. 280, 43087-43099 (2005).
- Schroder, K., Sweet, M. J., Hume, D. A. Signal integration between IFNgamma and TLR signalling pathways in macrophages. Immunobiology. 211, 511-524 (2006).
- Vinkemeier, U. Getting the message across, STAT! Design principles of a molecular signaling circuit. The Journal of Cell Biology. 167, 197-201 (2004).
- Brierley, M. M., Fish, E. N.
- Bentley, D. R.
- Grant, S. F., et al. Variant of transcription factor 7-like 2 (TCF7L2) gene confers risk of type 2 diabetes. Nature Genetics. 38, 320-323 (2006).
- Sladek, R., et al. A genome-wide association study identifies novel risk loci for type 2 diabetes. Nature. 445, 881-885 (2007).
- Wellcome Trust Case Control Consortium. Genome-wide association study of 14,000 cases of seven common diseases and 3,000 shared controls. Nature. 447, 661-678 (2007).
- Saxena, R., et al. Genome-wide association analysis identifies loci for type 2 diabetes and triglyceride levels. Science. 316, 1331-1336 (2007).
- Zeggini, E., et al. Replication of genome-wide association signals in UK samples reveals risk loci for type 2 diabetes. Science. 316, 1336-1341 (2007).
- Scott, L. J., et al. A genome-wide association study of type 2 diabetes in Finns detects multiple susceptibility variants. Science. 316, 1341-1345 (2007).
- Steinthorsdottir, V., et al. A variant in CDKAL1 influences insulin response and risk of type 2 diabetes. Nature Genetics. 39, 770-775 (2007).
- Salonen, J. T., et al. Type 2 Diabetes Whole-Genome Association Study in Four Populations: The DiaGen Consortium. American Journal of Human Genetics. 81, 338-345 (2007).
- Zeggini, E., McCarthy, M. I. TCF7L2: the biggest story in diabetes genetics since HLA. Diabetologia. 50, 1-4 (2007).
- Weedon, M. N.
- Hattersley, A. T. Prime suspect: the TCF7L2 gene and type 2 diabetes risk. The Journal of Clinical Investigation. 117, 2077-2079 (2007).
- Yochum, G. S., et al. Serial analysis of chromatin occupancy identifies beta-catenin target genes in colorectal carcinoma cells. Proceedings of the National Academy of Sciences of the United States of America. 104, 3324-3329 (2007).
- Duval, A., Busson-Leconiat, M., Berger, R., Hamelin, R. Assignment of the TCF-4 gene (TCF7L2) to human chromosome band 10q25.3. Cytogenet. Cell Genet. 88, 264-265 (2000).
- Pomerantz, M. M., et al. The 8q24 cancer risk variant rs6983267 shows long-range interaction with MYC in colorectal cancer. Nature Genetics. 41, 882-884 (2009).
- Tuupanen, S., et al. The common colorectal cancer predisposition SNP rs6983267 at chromosome 8q24 confers potential to enhanced Wnt signaling. Nature Genetics. 41, 885-890 (2009).
- Zhao, J., Schug, J., Li, M., Kaestner, K. H., Grant, S. F. Disease-associated loci are significantly over-represented among genes bound by transcription factor 7-like 2 (TCF7L2) in vivo. Diabetologia. 53, 2340-2346 (2010).
- Benjamini, Y., Yekutieli, D. Quantitative trait Loci analysis using the false discovery rate. Genetics. 171, 783-790 (2005).
