To illustrate typical outcomes of the described methods, we first used the Hemocyte Bleed/Scrape Assay to outline the progression of larval hemocyte numbers and their residence over the course of larval development (Figure 4). Resident and circulating larval hemocyte populations were isolated from single larvae (HmlΔ-GAL4, UAS-GFP; He-GAL4 to label the vast majority of larval hemocytes) and quantified using ImageJ. Cohorts of larvae sized 1.2 mm (~48 hr AEL or 1st instar), 2.5 mm (~80 hr AEL or late 2nd instar), and 3.5 mm (~96 hr AEL or 3rd instar) were examined (Figure 4). Hemocyte numbers expanded over the course of larval development, correlating with and exceeding previous estimates based on light microscopy of dye stained larvae7 and live counting of fluorescent protein labeled hemocytes through the larval cuticle6. In 1st instar larvae almost all hemocytes were resident, while the fraction of circulating hemocytes progressively increased over the course of larval development (Figure 4B,C), consistent with previous publications6,7.
Next we examined whether the method faithfully monitors the transition of hemocytes between the resident and circulating populations. Taking advantage of the phenomenon that resident hemocytes can be transiently detached by mechanical disturbance and they re-adhere to their resident sites spontaneously6, we dispersed resident hemocytes by vortexing with glass beads as described in the Hemocyte Disturbance Assay. Indeed, mechanical disturbance of larvae led to a dramatic increase in the population of circulating hemocytes at the expense of resident hemocytes (Figure 5). After a recovery period of 45 min, hemocytes had largely returned to their adherent state, both by visual inspection and by the assessed percentage of circulating cells (Figure 5D,E). As expected, total hemocyte numbers remained stable over time, despite the shift of hemocytes between the circulating and resident populations.
Several additional considerations were taken into account. To confirm that vortexing did not cause major tissue damage, vortexing with glass beads was performed in the presence of trypan blue (Sigma) for various time periods (1, 5, 20 min). Both 1 and 5 min vortexing did not cause any obvious tissue disruption, while 20 min vortexing resulted in small areas of damage, resembling damage caused by needle stitches used as positive control (Supplemental Figure 1). While internal damage of epidermis or other tissues without cuticle damage cannot be excluded, this scenario seems rather unlikely as hemocytes of 1 min and 5 min-treated larvae re-adhered in the expected pattern and time frame, suggesting larval integrity was not compromised (Supplemental Figure 1). In contrast, larvae vortexed for 20 min suffered from a lack of re-adhesion, and did not even show attachment of circulating hemocytes to epidermal wound sites, as has been described previously14.
Lastly, to demonstrate reproducibility of the method, we compared biological replicates of 2.5 mm larvae from the above two experiments, which were conducted by distinct experimenters. As illustrated in Supplemental Figure 2, both cohorts showed comparable total numbers of hemocytes per larva, and the percentage of circulating hemocytes. Student’s t testing showed no statistically significant differences, suggesting that the method is reproducible and broadly applicable.
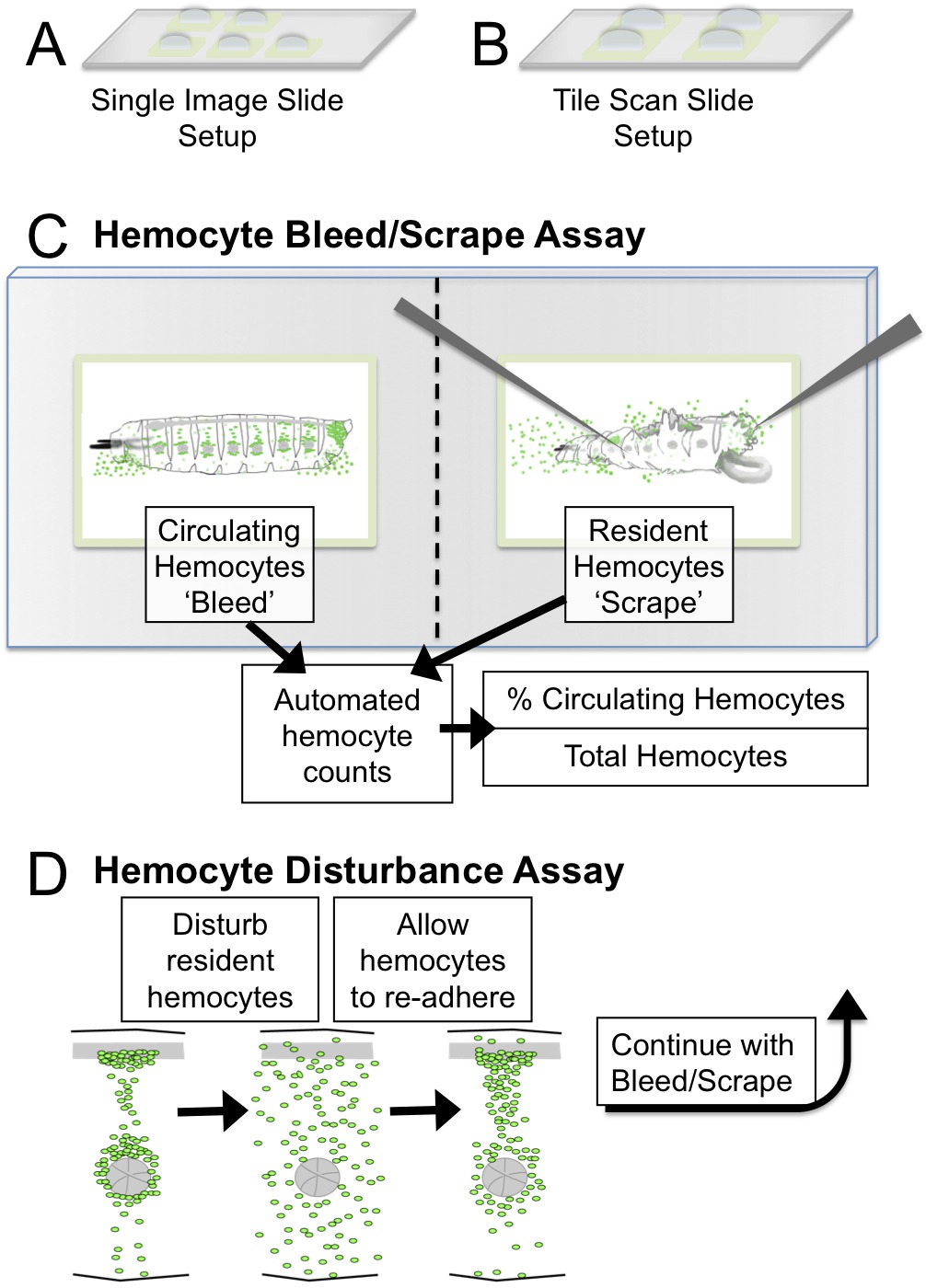
Figure 1. Hemocyte Bleed/Scrape and Disturbance Assay setup and schematic. (A) Single Image Slide Setup: five 2mm squares for imaging with a 5X objective. (B) Tile Scan Slide Setup: four 3 mm squares for imaging bleed/scrapes of ≤2.5 mm larvae with a tile scan microscope. Recommended objectives for imaging are 5X or 10X. (C) Bleed/Scrape Assay schematic and resulting quantifications using ImageJ. (D) In the Disturbance Assay, the hemocyte pattern is mechanically disrupted by vortexing larvae with glass beads. Larvae are allowed to recover over a period of 45 min during which hemocytes re-adhere to the Hematopoietic Pockets. The adhesive properties of hemocytes can be assessed by this method, quantifying the percentage of hemocytes in circulation after disturbance. Please click here to view a larger version of this figure.
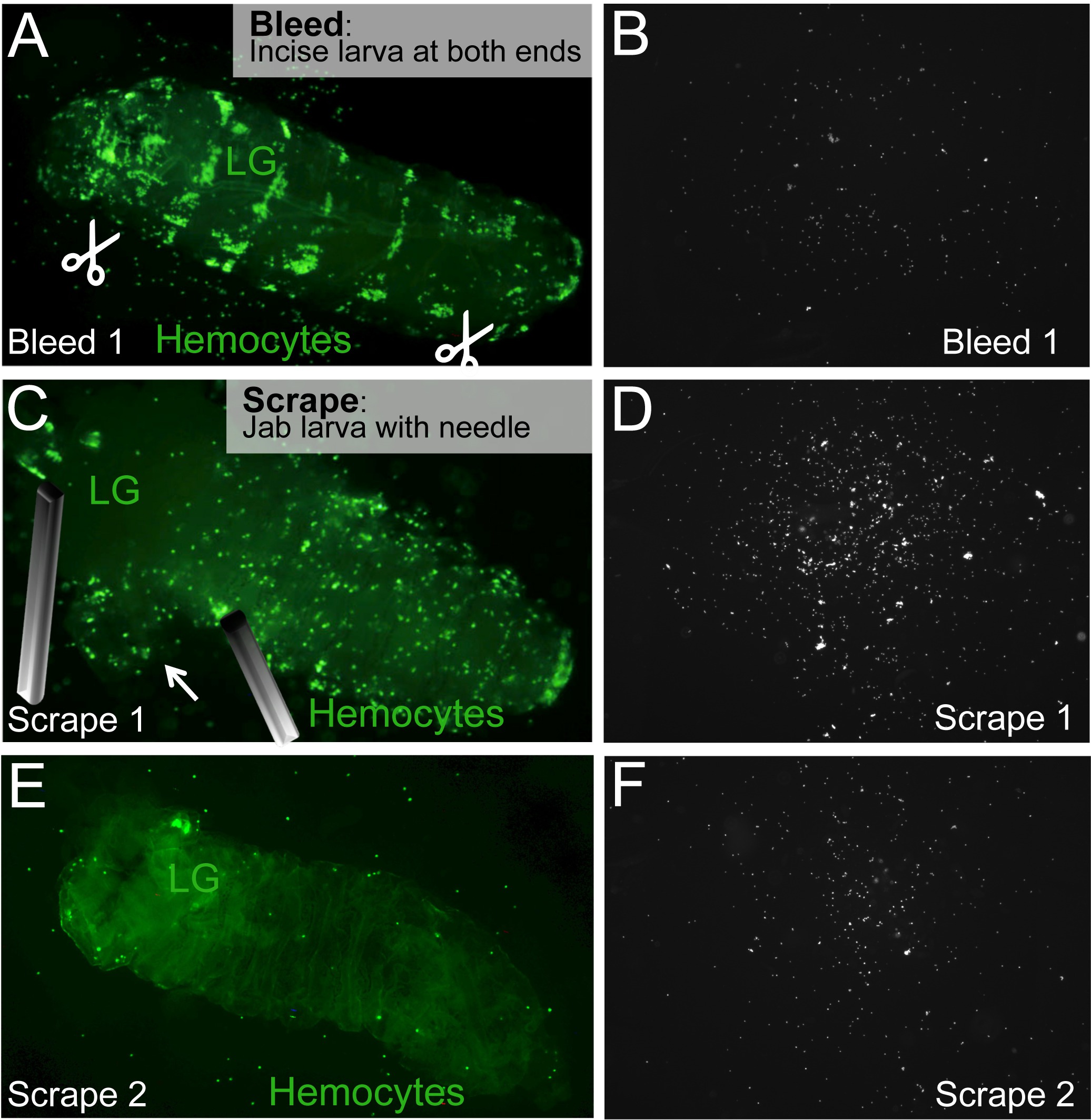
Figure 2. Bleed/Scrape Assay to release circulating and resident hemocytes. (A) To bleed a larva, ventral incisions at the posterior and anterior ends of the larva are made (scissors symbol). (B) Hemocytes in circulation will flow out of the incisions and settle on the surface of the slide. (C) The lymph gland (LG) is located and pinned down, without puncturing it. Resident hemocytes are released by jabbing and/or scraping the larva with a needle. (D) Resident hemocytes on slide. (E,F) The Scrape process is repeated until all resident hemocytes are released. The larval carcass containing the intact lymph gland is left behind. Please click here to view a larger version of this figure.

Figure 3. Automated quantification of hemocytes using ImageJ. (A,B) After opening a hemocyte image file in ImageJ, the Lower Threshold level is adjusted to account for all the cells in the image. (C,D) Analyze Particles requires setting the cell pixel size, circularity, and the result readout format (e.g., Overlay Outlines). (E) Summary window displaying the number of hemocytes. Please click here to view a larger version of this figure.

Figure 4. Representative Results (1). Hemocyte number and resident state over the course of larval development. (A) Overview of the larval stages used; 1st instar (48 hr AEL; ~1.2 mm length); 2nd instar (80 hr AEL; 2.5 mm length); 3rd instar (96 hr AEL; ~3.5 mm length). Genotype is HmlΔ-GAL4, UAS-GFP; He-GAL4. Stages were confirmed by assessing larval mouthhooks. (B) Bar diagram of circulating and resident hemocyte numbers at the respective larval stages. (C) Percentage of circulating hemocytes. Note that the fraction of circulating hemocytes increases disproportionally over the course of larval development. (D) Total hemocytes, resulting from the sum of circulating and resident hemocytes per larva. Hemocytes were quantified using the Bleed/Scrape method; n ≥ 6 larvae/condition, error bars show standard deviation, findings confirmed in 3 independent replicate experiments. Please click here to view a larger version of this figure.
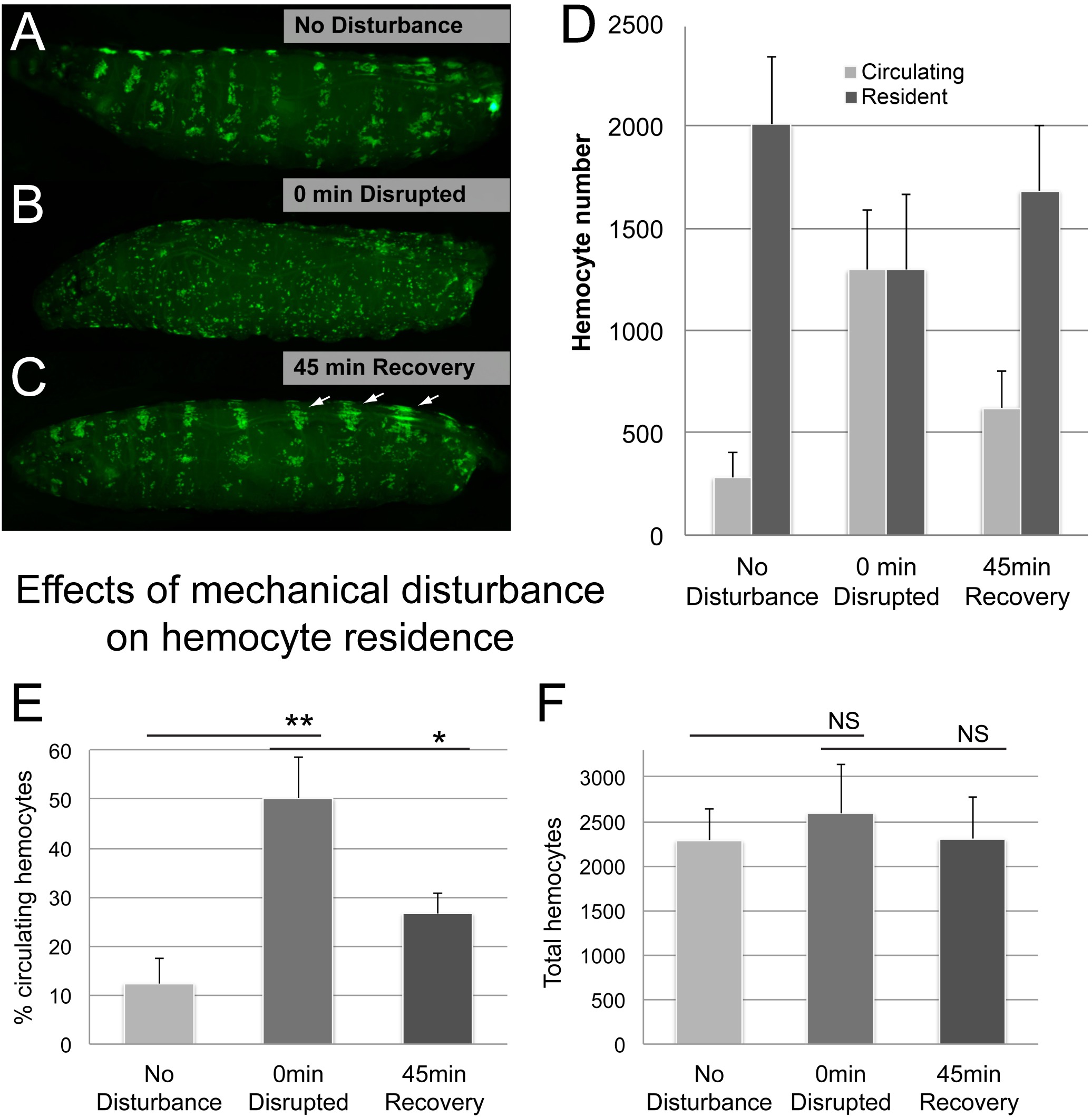
Figure 5. Representative Results (2). Effects of mechanical disturbance on hemocyte residence. (A-C) Example of a larva before and after vortexing with glass beads, followed by 45 min recovery. (A) No disturbance control; hemocytes are localized in Hematopoietic Pockets. (B) Disrupted hemocyte pattern at 0 min after vortexing larvae in a suspension of glass beads and water. (C) Hemocyte pattern at 45 min of recovery post-disturbance; many hemocytes have relocated to the Hematopoietic Pockets; note enlarged dorsal-vessel associated clusters and dorsal stripes which are predominant sites of early post-disturbance accumulation (arrows). Genotype is HmlΔ-GAL4, UAS-GFP; He-GAL4 x yw. (D) Percentage of circulating hemocytes quantified by the Bleed/Scrape method. (E) Total hemocytes, resulting from the sum of circulating and resident hemocytes per larva. n ≥ 4 larvae/condition, error bars show standard deviation, findings confirmed in 3 independent replicate experiments. Student’s t-test to confirm significance, NS (not significant), ** (p ≤ 0.05), ** (p ≤ 0.01). Please click here to view a larger version of this figure.
