

Introduction
Protéines résistance Myxovirus (Mx) sont un élément important de la défense immunitaire innée contre les pathogènes viraux. Ces protéines sont de grandes GTPases dynamine-like qui sont induits par les interférons de type III I et de type. Les gènes Mx correspondantes sont présentes dans pratiquement tous les vertébrés en une ou plusieurs copies et leurs produits géniques inhibent une large gamme de virus, y compris les Orthomyxoviridae (par ex., Virus de la grippe), des Rhabdoviridae (par ex., Virus de la stomatite vésiculaire), Bunyaviridae (par ex. , le virus de la crosse) et Retroviridae (par exemple, virus d'immunodéficience humaine-1) 1-4. On ne sait pas comment ces protéines reconnaissent une telle large éventail de virus, sans aucune séquence primaire partagé apparente motifs de ces virus. Analyser l'interaction des protéines Mx avec leurs cibles virales, impliquant potentiellement complexes d'ordre supérieur avec d'autres facteurs de la cellule hôte, aidera à comprendre les mécanismes moléculaires tHat ont évolué dans la course aux armements entre les virus et leurs hôtes.
L'interaction entre les protéines de mammifères Mx et cibles virales a été le plus étudié pour MxA humaine. MxA humaine peut inhiber la replication de nombreux virus, y compris l'orthomyxovirus virus influenza A et Thogoto. MxA lie les complexes de ribonucléoprotéine (virus Thogoto vRNPs), empêchant ainsi leur entrée nucléaire, ce qui entraîne l'infection de 5 bloc. Cette interaction entre MxA et Thogoto vRNPs de virus a été démontrée avec le co-sédimentation et des expériences de co-immunoprécipitation 6-9. Comment protéines Mx entravent virus de la grippe A est moins clair. Un problème majeur est qu'il ne est pas aisé de démontrer une interaction entre une protéine Mx et un produit du gène de la grippe. Un rapport a montré une interaction entre MxA humaine et la protéine NP de la grippe A cellules infectées par le virus 10. Cette interaction ne peut être représenté par la co-immunoprecipitation si les cellules avaient été traités avec les réticulants réactifs dithiobis (propionate de succinimidyle) avant la lyse, ce qui suggère que l'interaction est transitoire et / ou faible. Des études plus récentes ont montré que l'écart-Mx sensibilité de différentes souches de la grippe A est déterminée par l'origine de la protéine NP 11,12. Dans cette ligne, les virus grippaux A peuvent en partie échapper au contrôle Mx par mutation des résidus spécifiques dans la protéine NP 13. Cela suggère que la principale cible de virus influenza A pour l'hôte Mx est la protéine NP, probablement NP assemblé dans les complexes vRNP. Cependant, aucune de ces études plus récentes ont démontré une interaction entre l'influenza NP ou vRNPs et MxA soit humain ou de souris Mx1.
Récemment, nous avons montré, pour la première fois, une interaction entre la grippe et la protéine NP de souris MX1 avec une co-immunoprécipitation protocole optimisé 14, qui est décrit ici en détail. En général, les co-immunoprecipitation est l'une des approches biochimiques les plus fréquemment utilisés pour étudier les interactions protéine-protéine. Cette technique est souvent préférée à d'autres techniques, par exemple, deux hybrides de levure, car il permet d'étudier les interactions protéine-protéine dans leur environnement naturel. Co-immunoprécipitation peut être effectuée sur des protéines exprimées de manière endogène si des anticorps contre les protéines d'intérêt sont disponibles. En variante, les protéines d'intérêt peuvent être exprimés dans la cellule par transfection ou infection et une étiquette d'affinité peuvent être utilisés. En plus des avantages mentionnés ci-dessus, la co-immunoprécipitation protocole décrit permet la détection des interactions faibles de protéines et / ou transitoires. Le composant principal de ce protocole optimisé est l'addition de N-éthylmaléimide (NEM) dans le tampon de lyse des cellules. NEM est un réactif d'alkylation qui réagit avec les groupes thiol libres, tels que présents dans cysteines, à un pH de 6.5 à 7.5 pour former un thio-ester stable(Figure 1). A pH plus élevé, NEM peut également réagir avec des groupes amino ou 15 subir une hydrolyse. NEM est généralement utilisé pour bloquer les groupes thiol libres, afin d'empêcher la formation de liaisons disulfure ou inhiber l'activité enzymatique. Par exemple, NEM est souvent utilisé pour bloquer les enzymes, qui sont desumoylating proteases cysteine. Dans le protocole décrit co-immunoprécipitation, NEM a été inscrite dans le tampon de lyse car il a été rapporté que la sumoylation de protéines de la grippe peut influencer l'interaction entre des protéines virales 16. De façon inattendue, l'addition de NEM se est avéré être la clé pour documenter l'interaction entre l'influenza NP et Mx1 souris par co-immunoprécipitation. Il est difficile de comprendre pourquoi l'addition de NEM est crucial pour détecter l'interaction NP-MX1. Peut-être l'interaction est trop transitoire et / ou faible. NEM pourrait stabiliser l'interaction, par exemple, en conservant une conformation spécifique de Mx1, une protéine virale ou même une troisième compo inconnuenent. Un tel effet stabilisant de NEM a été observé précédemment, par exemple, par l'interaction entre la ribonucléotide réductase M1 et son inhibiteur de gemcitabine (F2dC) 17. NP MX1 et deux contiennent de multiples résidus de cysteine qui pourraient être modifiés par NEM. Par exemple, une étude récente de Rennie et al. A démontré qu'un sessiles MxA variante contient trois résidus de solvants cystéine exposés qui peuvent être modifiés par l'iodoacétamide. La mutation de ces résidus serines ne influence pas l'activité enzymatique de MxA, mais empêche l'agrégation à médiation par 18 disulfure. Étant donné que ces cysteines sont conservées dans Mx1, cela suggère que les cysteines dans Mx1 analogues peuvent être modifiées par NEM et en tant que tel ou sa conformation influence la solubilité. En outre, NEM peut également affecter l'activité GTPase de Mx1, ce qui est essentiel pour l'activité anti-influenza de Mx1, et ainsi stabiliser l'interaction entre MX1 et NP. Cependant, un effet direct de NEM sur la GTPase activité de Mx1 est peu probable, car NEM est également nécessaire pour détecter l'interaction entre la grippe NP et GTPase mutants inactifs de la protéine Mx1 14. De toute évidence, plus de recherche est nécessaire pour démêler l'effet de NEM sur l'interaction NP-MX1.
En résumé, la co-immunoprécipitation protocole décrit permet d'étudier l'interaction entre la protéine antivirale MX1 et sa cible virale, la protéine NP grippe. Ce protocole peut également être utilisé pour étudier d'autres interactions faibles ou transitoires qui dépendent de la stabilisation des conformations de protéines spécifiques. Interaction protéine-protéine qui dépendent de conformations spécifiques ont été décrits auparavant, par exemple, pour les protéines liant le calcium, tels que 19 la calmoduline. Enfin, le rôle bénéfique de NEM pourrait également être utilisé dans d'autres procédés qui permettent de détecter les interactions protéine-protéine, telles que les essais de co-sédimentation.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
Remarque: Le protocole de transfection et de co-immunoprécipitation suivante est établie pour un format de boîte de Pétri de 9 cm. D'autres formats sont également possible, après avoir escaladé le protocole.
1. ensemencement des rein embryonnaire humain (HEK) 293T
- Ensemencer les cellules HEK293T un jour avant la transfection à 1,2 x 10 6 cellules par 9 cm boîte de Pétri dans 12 ml de milieu de Eagle modifié par Dulbecco (DMEM) supplémenté avec 10% de sérum de veau foetal, 2 mM de L-glutamine, 0,4 mM de Na-pyruvate, 0,1 mM d'acides aminés non essentiels, 100 U / ml de pénicilline et 0,1 mg / ml de streptomycine.
- Cultiver les cellules 16 h à 37 ° C et 5% de CO 2.
- Inspecter visuellement la morphologie et la viabilité des cellules avec un microscope inversé la lumière avant la transfection. Les cellules ont besoin d'être sous-confluentes de l'efficacité de transfection optimale.
2. Le calcium-phosphate Transfection de cellules HEK293T
Note: Utilisation 0,5-1 pg de pCAXL-NP ou vide pCAXL plasmide en combinaison avec 1-3 ug de pCAXL-MX1 par 9 cm plat. Utiliser une quantité égale d'ADN total de plasmide dans tous les échantillons; ajuster avec le plasmide vide si nécessaire.
- Préparer les tampons de transfection suivantes:
- Préparation de Tris-EDTA (TE) avec des concentrations de 1,0 mM Tris-HCl pH 8,0 et EDTA 0,1 mM pH 8,0.
- Préparer BS / HEPES avec des concentrations de 25 mM de HEPES (5,96 g / l, 4- (2-hydroxyéthyl) -1-piperazineethanesulfonic), 274 mM de NaCl (16 g / L), KCl 10 mM (0,74 g / L), 1,5 mM NaHPO 4 · 12H 2 O (0,5 g / L) et de 11,1 mM de glucose (2 g / L). Ajuster le pH à 7,05.
- Préparer CaCl 2 / HEPES avec des concentrations de 1,25 M CaCl 2 · 2H 2 O (183,8 g / L) et 125 mM de HEPES (29,79 g / L). Ajuster le pH à 7,05 avec du NaOH.
- Réchauffez les tampons de transfection à 37 ° C avant utilisation.
- Préparer les échantillons de plasmide par dilution de l'ADN de plasmide dans 600 ul de TE.Préparer ces mélanges dans les puits d'une plaque à 6 puits.
- Ajouter 150 pi de CaCl2 / HEPES d'une manière sage de déposer pour les échantillons de plasmides et mélanger par pipetage 3 fois de haut en bas.
- Préparer la solution de transfection à goutte ajoutant sage la solution de plasmide (TE + DNA + CaCl2 / HEPES; 750 pi) à 750 pi de tampon BS / HEPES prévues dans une plaque à 6 puits frais. Distribuer la solution de plasmide uniformément sur le tampon complète puits contenant de BS / HEPES.
- Agiter la solution de transfection sur un agitateur de plaque pendant 90 secondes à 1000 RPM.
- Incuber le mélange pendant 5 min à température ambiante.
- Ajouter la solution de transfection (1,5 ml) goutte à goutte aux cellules. Utiliser une micropipette P1000 à goutte la solution de transfection sur les cellules. Disperser le mélange sur le 9 cm plat complet Petri et secouer la plaque très doucement.
- Incuber les cellules à 37 ° C et 5% de CO2 pendant 6 heures. Puis retirez le support par aspiration et remplacer immédiatement wie 12 ml du milieu frais, préchauffé. Ajouter doucement le milieu frais aux cellules pour empêcher le détachement des cellules. Pour cela, maintenez la pointe de la pipette contre le côté du puits et poussez doucement sur le milieu.
- Incuber les cellules pendant une 16 heure supplémentaire à 37 ° C et 5% de CO 2.
3. Co-immunoprécipitation
Remarque: Effectuez la co-immunoprécipitation 24 heures après transfection.
- Préparation du tampon de lyse et de faible teneur en sel du tampon de lavage élevée en sel.
- Préparer une solution mère de 2 M N-éthylmaléimide (NEM) en pesant la quantité de NEM et le dissolvant dans de l'éthanol absolu. Préparer la solution mère NEM frais avant utilisation.
ATTENTION: NEM est très toxique, préparer et utiliser cette solution mère dans une hotte. - Préparer du tampon de lyse faible de sel à des concentrations de 50 mM Tris-HCl pH 8, NaCl 150 mM, acide éthylènediaminetétraacétique 5 mM (EDTA), 1% de NP40 et un cocktail d'inhibiteurs de protease (tableau 1 dissoudret dans un tampon de lyse de 50 ml). Ajouter NEM jusqu'à une concentration finale de 25 mM (ce est-à diluer 1:80). Garder sur la glace après avoir ajouté les inhibiteurs de protéase et NEM.
Note: Toujours ajouter les inhibiteurs de protéase et NEM fraîchement avant l'utilisation. - Préparer une solution de lavage riche en sel à des concentrations de 50 mM Tris-HCl pH 8, NaCl 500 mM, EDTA 5 mM et 1% de NP40. Notez que le tampon de lavage en sel élevée ne contient pas de NEM.
- Préparer une solution mère de 2 M N-éthylmaléimide (NEM) en pesant la quantité de NEM et le dissolvant dans de l'éthanol absolu. Préparer la solution mère NEM frais avant utilisation.
- Préparation de lysats cellulaires.
- Retirez le moyen et laver les cellules avec 2 ml de glace froide saline tamponnée phosphate (PBS). Ajouter très doucement le tampon de lavage, les cellules HEK293T se détachent facilement.
- Retirez le PBS et ajouter 600 ul de tampon de lyse de sel de la glace froide bas par 9 cm boîte de Pétri.
- Incuber les plaques pendant 20 minutes sur la glace. Assurez-vous que les plaques sont maintenues horizontal, pour assurer une couverture complète de la surface de la plaque avec du tampon de lyse. Secouer doucement les plaques toutes les 5 min.
- Recueillir le lysat cellulaire dans une microcentrifugeuse de 1,5 mlet centrifuger le tube pendant 3 min à 4 ° C et 16 000 x g pour sédimenter la fraction insoluble.
- Transférer la fraction soluble, ce est à dire le lysat cellulaire à un nouveau tube de 1,5 ml à centrifuger et garder sur la glace. Poursuivra immédiatement le protocole de co-immunoprécipitation, pour empêcher la dissociation des protéines qui interagissent. Effectuez toutes les étapes suivantes autant que possible sur la glace ou à 4 ° C à limiter l'activité protéolytique dans les lysats.
- Génération de complexes immuns.
Remarque: Dans cette étape, la protéine d'intérêt est liée par l'anticorps approprié. Pour étudier l'interaction NP-MX1, utiliser un anticorps monoclonal anti-NP souris.- Pour chaque échantillon, mélanger 135 pi de lysat avec 2 pi d'anticorps monoclonal anti-NP et 113 ul de tampon de lyse faible de sel (volume total de 250 ul). Rangez le lysat restant à -20 ° C pour une analyse ultérieure tels que Western blot, de documenter les niveaux de l'intera supposé d'expressionpartenaires ction dans les cellules transfectées.
Remarque: vous pouvez mesurer la concentration de protéines du lysat (par exemple avec le réactif Bradford) et utiliser une quantité fixe de protéines totales, par exemple, 400 mg, pour chaque lysat. - Incuber le mélange anticorps lysat pendant 3 heures sur une roue tournant à 4 ° C. Cette étape peut être étendue à une incubation pendant une nuit.
- Pour chaque échantillon, mélanger 135 pi de lysat avec 2 pi d'anticorps monoclonal anti-NP et 113 ul de tampon de lyse faible de sel (volume total de 250 ul). Rangez le lysat restant à -20 ° C pour une analyse ultérieure tels que Western blot, de documenter les niveaux de l'intera supposé d'expressionpartenaires ction dans les cellules transfectées.
- Préparation de la protéine G perles.
Remarque: Les billes de protéine G sont expédiées et stockées dans 20% d'éthanol pour la conservation. La suspension de perles est normalement constitué de 50% de perles et les perles ont besoin d'être lavé avant d'être utilisés pour immunoprécipiter les complexes immuns.- Utilisez 50 ul de billes, soit 100 pi de cordon-boue, pour chaque échantillon. Laver la quantité de billes pour tous les échantillons nécessaires à l'analyse de co-immunoprécipitation dans un tube. Couper l'extrémité d'une pointe de pipette de 1 ml pour faciliter l'ajout de la suspension de perles.
- Centrifuger la protéine G perles suspension à 8000 xg et 4 ° C pendant 30 sec. Retirer la solution d'éthanol et d'ajouter un volume égal de tampon de lyse faible de sel. Centrifugeuse la protéine G perles suspension à 8000 xg et 4 ° C pendant 30 secondes et retirez délicatement le surnageant. Répétez cette étape de lavage 3 fois.
Remarque: Le tampon de lyse faible de sel utilisée pour laver les billes ne ont pas besoin de contenir des inhibiteurs de la protéase ou de NEM. - Estimer le volume des billes de protéine G et ajouter un volume égal de tampon de lyse faible de sel pour faire une nouvelle suspension de billes à 50% dans du tampon de lyse faible de sel.
- Pour chaque échantillon, transférer 100 pi de perles suspension dans un nouveau tube de 1,5 ml à centrifuger et stocker sur la glace jusqu'à utilisation. Faire attention à remettre en suspension la suspension de billes avant de se diviser, comme ces billes sédimentent rapidement au fond du tube.
- Immunoprécipitation des complexes immuns par la protéine G perles et leur élution.
- Avant d'utiliser les billes de protéine G pour l'immunoprécipitation, centrifuger les tubes 30 secondes à 8000 x g et 4 ° C etvérifier par examen visuel qu'il existe un nombre égal de billes présentes dans tous les échantillons. Si nécessaire, ajuster la quantité de perles dans certains des échantillons et centrifuger à nouveau. Jeter les surnageants. Veillez à ne pas déranger les protéines G perle granulés.
- Centrifuger brièvement les complexes immuns (ie., Lysats avec un anticorps, 250 pi) pendant 30 secondes à 8000 xg et 4 ° C pour recueillir l'échantillon complet au fond du tube. Transférez les complexes immuns aux perles de protéines G (50 pi).
- Incuber 60 min sur une roue tournant à 4 ° C. Ne pas incuber ces complexes immuns plus de 75 min avec les billes pour réduire la liaison non spécifique des protéines à la protéine G perles.
- Centrifuger les perles de protéines G (avec complexes immuns liés) pendant 30 secondes à 8000 xg et 4 ° C et enlever les surnageants. Veillez à ne pas déranger les protéines G perle granulés. Facultatif: stocker ces surnageants à 4 ° C ou -20 ° C pour analyse ultérieure,par exemple, pour estimer la quantité de protéine non liée.
- Laver les billes de protéine G pendant environ 5 min avec 900 ul de tampon de lyse élevée en sel. Assurez-vous que les perles sont complètement remises en suspension dans le tampon de lavage pour le lavage optimal. Centrifuger les protéines G perle pour 30 secondes à 8000 xg et 4 ° C et jeter le surnageant. Répétez cette étape de lavage 4 fois. Veillez à ne pas déranger les perles culot de protéine G pour éviter la perte de matériel immunoprécipité.
- Après la dernière étape de lavage, ajouter 50 ul de tampon d'échantillon Laemmli 2x aux billes et chauffer la suspension pendant 10 min à 95 ° C pour éluer les (co) protéines immunoprécipitées.
- Préparer 10 ml de tampon Laemmli 6x avec 1 g de sulfate de dodécyle de sodium, 3,5 ml de glycerol, 3,5 ml de 1 M Tris-HCl pH 6,8 et 420 ul de β-mercaptoéthanol. Ajuster à un volume total de 10 ml par addition d'eau distillée. Diluer trois fois à l'eau distillée pour obtenir un tampon de Laemmli 2x.
ATTENTION: &# 946; mercaptoéthanol est toxique, préparer et utiliser un tampon Laemmli dans une hotte.
- Préparer 10 ml de tampon Laemmli 6x avec 1 g de sulfate de dodécyle de sodium, 3,5 ml de glycerol, 3,5 ml de 1 M Tris-HCl pH 6,8 et 420 ul de β-mercaptoéthanol. Ajuster à un volume total de 10 ml par addition d'eau distillée. Diluer trois fois à l'eau distillée pour obtenir un tampon de Laemmli 2x.
- Après chauffage, centrifuger les protéines G perle pour 30 secondes à 8000 xg et stocker les échantillons à 4 ° C (à court terme) ou -20 ° C (à long terme).
4. Analyser le (co-) immunoprécipités Protéines
- Visualiser les protéines présentes dans le lysat cellulaire et l'éluat de la co-immunoprécipitation par SDS-PAGE et transfert de Western 20 21,22. Charger généralement la moitié de l'éluat Laemmli sur gel. Veillez à ne pas déranger les billes de protéine G granulés lors de la prise d'échantillons pour le chargement du gel. MX1 et expression NP ont été révélés avec un anticorps anti-anticorps MX1 et NP-anti, 14 respectivement. Les bandes ont été détectées avec une chimioluminescence à base de HRP et un révélateur de film radiographique.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
N-éthylmaléimide est un composé organique qui peut être utilisé pour modifier de manière irréversible des groupes thiol libres, par exemple pour inhiber les proteases cysteine (figure 1).
La protéine antiviral inhibe la grippe A Mx1 la replication du virus en interagissant avec la nucléoprotéine virale. Le protocole de co-immunoprécipitation optimisé décrit ici permet d'étudier cette interaction NP-MX1. des cellules HEK293T ont été transfectées avec des vecteurs d'expression pour la protéine antivirale Mx1 en l'absence ou en présence de la protéine NP grippe. Ensuite, la protéine NP a été tiré vers le bas à partir de lysats cellulaires totaux avec un anticorps monoclonal NP-spécifique. La figure 2 montre que la protéine ne est Mx1 co-immunoprécipitées en présence de co-exprimé NP. Possible non spécifique co-immunoprécipitation de Mx1 en l'absence de NP est causée soit par déroulant non spécifique de la protéine Mx1 par l'anticorps anti-NP ou par liaison non spécifique de la Mx1protéines G perles. Par conséquent, toujours inclure un contrôle négatif pour évaluer cette co-immunoprécipitation non spécifique. La figure 3 montre que l'interaction NP-MX1 ne peut être détectée en présence de NEM. Dans cette expérience, la liaison non spécifique de Mx1 aux billes de protéine G a été évaluée avec une réaction de co-immunoprécipitation de contrôle en l'absence d'anticorps anti-NP.
Ce protocole peut également être utilisé pour étudier l'interaction entre MX1 et la protéine NP grippe isolé à partir de cellules infectées ou à partir de virions. Pour cette application, le protocole ci-dessus a été légèrement adaptée en combinant des lysats de cellules exprimant Mx1 avec des lysats contenant la protéine NP du virus avant de démarrer le protocole de co-immunoprécipitation. La figure 4 montre la co-immunoprécipitation de Mx1 avec NP isolé à partir de cellules transfectées, les cellules infectées ou virions.
En conclusion, ces résultats montrent que cette co-immunoprécipitation protCLO peut être utilisée pour étudier l'interaction entre une protéine antivirale et sa cible virale.
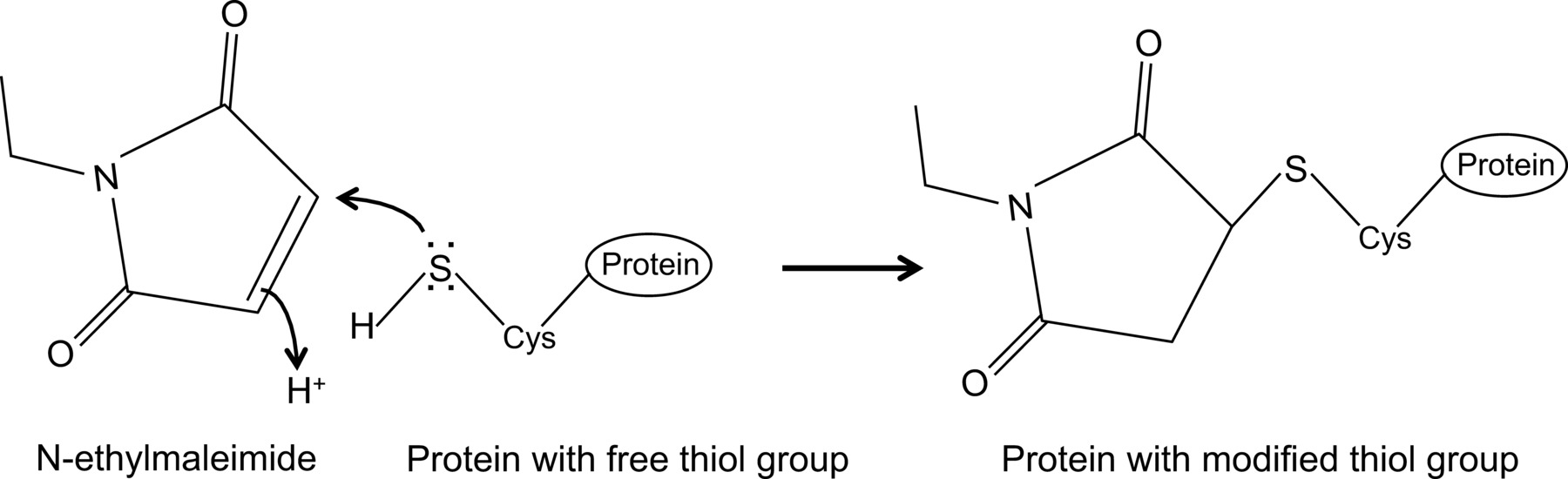
Figure 1:. Modification irréversible des groupes thiols libres par N-éthylmaléimide Se il vous plaît cliquer ici pour voir une version plus grande de cette figure.

Figure 2:. Mx1 interagit avec NP analyse par Western blot d'une expérience de co-immunoprécipitation avec deux échantillons: l'un dans lequel les deux MX1 et NP sont présents et une configuration de commande dans lequel la protéine NP est absent. NP a été immunoprécipité avec des anticorps anti-NP et NP et Mx1 ont été visualisées par western blot. Ce chiffre a été modifié from 14.

Figure 3: N-éthylmaléimide est important de détecter la NP - interaction Mx1 analyse par Western blot d'une expérience de co-immunoprécipitation réalisée en présence ou en l'absence de N-éthylmaléimide.. NP a été immunoprécipité avec des anticorps anti-NP et MX1 et NP ont été visualisées par western blot.

Figure 4:. Mx1 interagit avec NP isolé à partir de cellules infectées ou de virions lysats contenant Mx1 ont été combinés avec des lysats contenant du NP à partir de différentes sources: un lysat témoin (pas vRNPs), transfectées vRNPs, vRNPs partir de cellules infectées ou vRNPs isolé à partir de virions de la grippe A . Après le mélange des lysats, co-immunopreprécipitation avec des anticorps anti-NP a été effectuée et MX1 et NP ont été visualisée par western blot.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
L'étude de l'interaction entre les protéines antivirales et leurs cibles virales est très important de comprendre les détails du mécanisme antiviral de ces protéines. Cela peut donner de nouvelles perspectives sur la façon dont les virus et leurs hôtes co-évolué et être la base pour le développement de nouvelles stratégies antivirales. Le protocole de co-immunoprécipitation optimisé décrit ici permet d'étudier l'interaction entre la protéine de la souris MX1 et sa cible virale, la protéine NP grippe. L'aspect le plus important de ce protocole est l'addition de NEM dans le tampon de lyse, comme l'interaction NP-MX1 est indétectable en l'absence de NEM (figure 3). A ce jour, on ne sait pas pourquoi la présence de NEM est essentiel de détecter cette interaction. Cependant, ce protocole peut être utile pour étudier d'autres interactions faibles et / ou transitoires qui dépendent de la stabilisation des conformations spécifiques de protéines, en particulier si cysteines sont impliquées.
Unelimitation importante des tests de co-immunoprécipitation en général est la disponibilité d'anticorps spécifiques et de haute qualité qui reconnaissent des epitopes conformationnels dans l'un des partenaires d'interaction à haute affinité. L'interaction NP-MX1 n'a pas pu être démontré avec notre maison produit antisérum polyclonal anti-MX1. Cet antisérum immunoprécipite également la protéine NP grippe, même en l'absence de Mx1. En outre, l'anticorps monoclonal anti-NP qui a été utilisé, reconnaît la protéine NP de A / Porto Rico / 8/34 souche de la grippe, mais, malheureusement, il ne est pas adapté pour tirer vers le bas NP de souches de virus de la grippe aviaire. Une autre limitation de ce protocole de co-immunoprécipitation est la liaison non spécifique de Mx1 à la protéine G perles. Cette dernière fixation peut être surmonté en augmentant la concentration de sel dans le tampon de lavage et la réduction du temps de contact entre le lysat et la protéine G perle. En outre, en réduisant la quantité de billes utilisé pour chaque immunoprécipitationréaction de 50 ul à 25 ul, peut en outre diminuer la liaison non spécifique de Mx1 à ces billes. En général, le menu déroulant involontaire de protéines contaminantes, provoqué par la liaison non spécifique aux billes de protéine G, peut également être réduite par d'autres stratégies. Par exemple, ces protéines peuvent être enlevés lors d'une étape de pré-clair, dans lequel ces protéines sont éliminées par incubation du lysat avec des billes de protéine G en l'absence d'anticorps. Les protéines contaminantes sont ensuite éliminés avec les perles et le lysat pré-autorisé est utilisé pour co-immunoprécipitation. Cette stratégie ne est avantageux que les protéines contaminantes sont différentes de la protéine (s) à l'étude. Alternativement, les sites de liaison non spécifiques sur la protéine G perles pourraient être bloquée avec de la BSA. Cependant, cette stratégie ne est recommandée si les perles sont d'abord revêtues avec l'anticorps, comme la BSA peut également réduire la précipitation des complexes immuns (par exemple, la liaison anticorps-réduit par les billes revêtues). Prisen même temps, il est très important d'inclure des contrôles appropriés pour exclure la liaison non spécifique des protéines d'intérêt à la protéine G ou à des billes les anticorps utilisés.
Le protocole de co-immunoprécipitation décrit peut être modifié pour étudier l'interaction de la protéine avec Mx1 NP présent dans un environnement différent, par exemple, dans les cellules infectées ou dans vRNPs purifiés. Mx1 inhibe l'expression de protéines virales, y compris NP, lors infection par le virus influenza 23. Par conséquent, il est techniquement très difficile d'étudier l'interaction NP-MX1 Mx1 transfectées dans des cellules qui sont infectées par la suite. Cependant, ce protocole de co-immunoprécipitation peut également être effectuée après la combinaison des lysats de cellules exprimant Mx1 et les cellules infectées, ce qui permet une détection réussie de l'interaction NP-MX1 (figure 4). Comme l'objectif essentiel de NEM est pas connue, NEM a été ajouté au cours de la lyse des deux populations de cellules. Si on le souhaite, le pH dule tampon de lyse peut être modifié. Les expériences décrites ici sont effectuées à pH 8 (comme dans Turan et al. 10), mais le co-immunoprécipitation NP-MX1 a également été réalisées avec succès avec un tampon de lyse pH 7,2. En fait, un pH de 7,2 est préférée, car à ce pH NEM réagit exclusivement avec les groupes thiol libres et ce pH augmente également le rendement d'extraction de la protéine Mx1. En outre, les billes de protéine G peuvent être remplacés par des billes de protéine A, en fonction des espèces d'hôtes à partir desquels les anticorps utilisés sont dérivés. Cependant, des résultats plus propres sont obtenus avec billes de protéine G. Enfin, ce protocole peut également être utilisé pour étudier l'interaction entre MX1 et d'autres protéines de la grippe, par exemple, 14 PB2. Dans ce cas, un épitope V5 marqué PB2 a été utilisé, ce qui pourrait en principe être combiné avec un gel d'affinité disponible dans le commerce d'agarose anti-V5.
Dans les expériences futures, ce protocole pourrait être utile pour identifier les régions dans MX1 etNP qui sont importants pour l'interaction NP-MX1. Une telle région pourrait être Mx1 boucle L4, comme l'a montré cette boucle pour être critique pour l'interaction entre MxA et le virus Thogoto NP 8,9. Si des anticorps NP-spécifiques appropriés deviennent disponibles, ce protocole permettra également de déterminer si la sensibilité accrue de la grippe aviaire souches A pour Mx1 en corrélation avec une plus forte interaction NP-MX1 ou non, ce qui pourrait aider à comprendre le mécanisme de la sensibilité Mx. En outre, l'interaction entre les composants MX1 et de virus appartenant à la famille des Orthomyxoviridae autres que, peut également être traitée par ce protocole adapté co-immunoprécipitation. Enfin, d'autres études pour démêler l'effet de NEM sur le NP-MX1 sera très précieuse pour mieux comprendre cette interaction spécifique, mais aussi pour avoir un aperçu dans les applications plus larges de l'addition de ce composé au cours d'expériences de lyse cellulaire et co-immunoprécipitation.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Les auteurs déclarent qu'ils ne ont aucun intérêt financier concurrentes.
Acknowledgments
Ce travail a été soutenu par FWO-Vlaanderen, le projet IOF IOF10 / Startt / 027 et l'Université de Gand spécial de recherche Grant BOF12 / GOA / 014.
Materials
| Name | Company | Catalog Number | Comments |
| DMEM high glucose | Gibco | 52100-047 | |
| N-Ethylmaleimide | Sigma | E-3876 | Toxic |
| Igepal CA-630 | Sigma | I-30212 | also known as NP40 |
| Protease inhibitor cocktail | Roche | 11 873 580 001 | |
| anti-NP monoclonal antibody | NIH Biodefense and Emerging Infections Research Resources Repository | NR-4282 | ascites blend of clones A1 and A3 |
| anti-RNP polyclonal serum | NIH Biodefense and Emerging Infections Research Resources Repository | NR-3133 | directed against A/Scotland/840/74 (H3N2) |
| Protein G Sepharose 4FF | GE Healthcare | 17-0618-01 | |
| Hyperfilm ECL 18 x 24 cm | GE Healthcare | 28-9068-36 | |
| ECL western blotting substrate | Pierce | 32106 |
References
- Verhelst, J., Hulpiau, P., Saelens, X. Mx proteins: antiviral gatekeepers that restrain the uninvited. Microbiol Mol Biol Rev. 77 (4), 551-566 (2013).
- Goujon, C., et al. Human MX2 is an interferon-induced post-entry inhibitor of HIV-1 infection. Nature. 502 (7472), 559-562 (2013).
- Kane, M., et al. MX2 is an interferon-induced inhibitor of HIV-1 infection. Nature. 502 (7472), 563-566 (2013).
- Liu, Z., et al. The interferon-inducible MxB protein inhibits HIV-1 infection. Cell Host Microbe. 14 (4), 398-410 (2013).
- Kochs, G., Haller, O. Interferon-induced human MxA GTPase blocks nuclear import of Thogoto virus nucleocapsids. Proc Natl Acad Sci U S A. 96 (5), 2082-2086 (1999).
- Flohr, F., Schneider-Schaulies, S., Haller, O., Kochs, G. The central interactive region of human MxA GTPase is involved in GTPase activation and interaction with viral target structures. FEBS Lett. 463 (1-2), 24-28 (1999).
- Kochs, G., Haller, O. GTP-bound human MxA protein interacts with the nucleocapsids of Thogoto virus (Orthomyxoviridae). J Biol Chem. 274 (7), 4370-4376 (1999).
- Mitchell, P. S., et al. Evolution-guided identification of antiviral specificity determinants in the broadly acting interferon-induced innate immunity factor MxA. Cell Host Microbe. 12 (4), 598-604 (2012).
- Patzina, C., Haller, O., Kochs, G. Structural requirements for the antiviral activity of the human MxA protein against Thogoto and influenza A virus. J Biol Chem. 289 (9), 6020-6027 (2014).
- Turan, K., et al. Nuclear MxA proteins form a complex with influenza virus NP and inhibit the transcription of the engineered influenza virus genome. Nucleic Acids Res. 32 (2), 643-652 (2004).
- Dittmann, J., et al. Influenza A virus strains differ in sensitivity to the antiviral action of Mx-GTPase. J Virol. 82 (7), 3624-3631 (2008).
- Zimmermann, P., Manz, B., Haller, O., Schwemmle, M., Kochs, G. The viral nucleoprotein determines Mx sensitivity of influenza A viruses. J Virol. 85 (16), 8133-8140 (2011).
- Manz, B., et al. Pandemic influenza A viruses escape from restriction by human MxA through adaptive mutations in the nucleoprotein. PLoS Pathog. 9 (3), e1003279 (2013).
- Verhelst, J., Parthoens, E., Schepens, B., Fiers, W., Saelens, X. Interferon-inducible protein Mx1 inhibits influenza virus by interfering with functional viral ribonucleoprotein complex assembly. J Virol. 86 (24), 13445-13455 (2012).
- Brewer, C. F., Riehm, J. P. Evidence for possible nonspecific reactions between N-ethylmaleimide and proteins. Anal Biochem. 18 (2), 248-255 (1967).
- Wu, C. Y., Jeng, K. S., Lai, M. M. The SUMOylation of matrix protein M1 modulates the assembly and morphogenesis of influenza A virus. J Virol. 85 (13), 6618-6628 (2011).
- Chen, Z., Zhou, J., Zhang, Y., Bepler, G. Modulation of the ribonucleotide reductase M1-gemcitabine interaction in vivo by N-ethylmaleimide. Biochem Biophys Res Commun. 413 (2), 383-388 (2011).
- Rennie, M. L., McKelvie, S. A., Bulloch, E. M., Kingston, R. L. Transient dimerization of human MxA promotes GTP hydrolysis, resulting in a mechanical power stroke. Structure. 22 (10), 1433-1445 (2014).
- Gifford, J. L., Walsh, M. P., Vogel, H. J. Structures and metal-ion-binding properties of the Ca2+-binding helix-loop-helix EF-hand motifs. Biochem J. 405 (2), 199-221 (2007).
- Separating Protein with SDS-PAGE. , JoVE. Available from: http://www.jove.com/science-education/5058/separating-protein-with-sds-page (2014).
- Gallagher, S., Chakavarti, D. Immunoblot analysis. J Vis Exp. (16), (2008).
- Eslami, A., Lujan, J. Western blotting: sample preparation to detection. J Vis Exp. (44), (2010).
- Pavlovic, J., Haller, O., Staeheli, P. Human and mouse Mx proteins inhibit different steps of the influenza virus multiplication cycle. J Virol. 66 (4), 2564-2569 (1992).
