1. Construction of 150 mm Diameter Shear Rings (Figure 1)
NOTE: Shear rings may be constructed to create many different dimensions by varying the outer and inner Petri dish sizes, resulting in devices with different total surface areas, cell yields and developed ranges of shear forces. This report describes a 150 mm dish combined with an inner 100 mm dish for a total surface area of 98 cm2 (Figure 2).
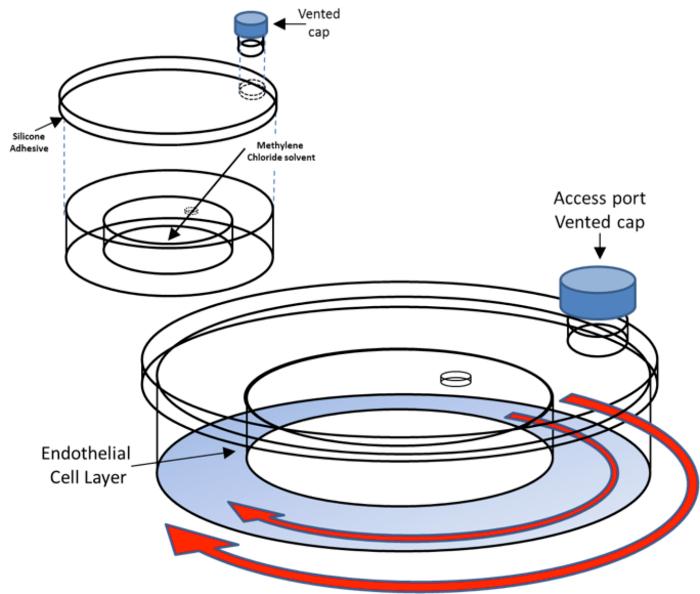
Figure 1. Shear ring assembly. The top portion of the figure shows a partially assembled shear ring. Inject 0.5 ml of methylene chloride through the 3 mm hole with a transfer pipet if a tight seal has not completely formed between the inner and outer dishes. The shear ring is sealed shut by applying a 1 mm thick bead of silicon rubber adhesive around the inner edge of the shear ring top. The bottom portion of the figure shows the assembled shear ring. The blue represents the area of plated endothelial cells. The outer and inner red arrows indicate the orbital motion of the shear ring and media inside the shear ring when placed on an orbital shaker. Please click here to view a larger version of this figure.

Figure 2. Surface measurements for a 150 mm shear ring (not drawn to scale). Top panel shows centrifugal shifting of fluid towards outer ring in response to orbital rotation. Please click here to view a larger version of this figure.
- Create a shear ring construction template by generating a 150 mm outer edge profile in presentation software with a 100 mm internal diameter placed in the center of the 150 mm circle. Print the template on a sheet of A4 white paper.
- Pipet 10 ml of methylene chloride (dichloromethane) into a 150 mm glass Petri dish. It is critical that the dish is glass and not polystyrene (or other vinyl plastic), as methylene chloride solubilizes most plastics and is used here to join plastic components.
CAUTION: Use gloves for all construction with adequate hood ventilation. Methylene chloride is a contact irritant and is hepatotoxic. - Align a 150 mm plastic Petri dish onto the outer shear ring template.
- Drill a 3 mm hole in the center of a 100 mm dish using a rotary tool. Remove any plastic shavings produced from the drilling.
- While holding with a gloved hand, invert and dip the top edge of the 100 mm Petri dish from the previous step into the pool of methylene chloride for approximately 3 sec.
- Transfer the "wetted" 100 mm dish edge-down onto the center of a 150 mm dish, carefully aligning the 100 mm dish onto the marked template. The methylene chloride will fuse the plastic, joining the 100 mm and 150 mm dishes. Gently rotate the 100 mm dish clockwise and counterclockwise a few times to ensure good adhesion to the 150 mm dish.
NOTE: Take great care to ensure proper alignment of the inner dish to the center of the outer dish. Eccentric alignment may result in variations of the shear stress at different locations in the shear ring. Take care to not allow methylene chloride to accidentally drip onto the outer track portion of the 150 mm Petri dish during the placement of the 100 mm dish. This will melt the plastic on the bottom surface where cells will be growing, creating an uneven surface that may cause flow disturbances. - Once the methylene chloride has dried, flip the newly bound Petri dishes over and carefully inspect the points of contact to ensure that a tight seal has been formed between the two Petri dishes.
- If a tight seal has not completely formed, inject 0.5 ml of methylene chloride through the 3 mm hole with a transfer pipet. Pick up the dish and gently rotate it to allow the methylene chloride to reach the edge.
NOTE: If rotated too quickly, the methylene chloride can spill into the outer portion of the 150 mm dish, deforming the surface where cells will grow and ruining the shear ring. Leaking shear rings should be discarded. - Seal off the hole in the 100 mm dish by applying a 3 mm bead of silicon rubber sealant.
- Using a rotary tool fitted with a flat cutting head, cut off the top 3 cm portion of a 15 ml conical polystyrene tissue culture tube, leaving at least 1 cm of the tube below the cap. Polish the edge of the cut tube until smooth and flat using the flat side of the cutting head. Remove any plastic shavings produced by grinding.
- Trace the cut off 15 ml conical tube onto the lid of the 150 mm dish with a marker, approximately 0.5 cm away from the edge of the dish. Drill a hole inside the drawn circle, leaving a margin approximately 1-2 mm from the edge of the circle.
- Place the cut off conical tube top over the drilled hole. Using a transfer pipet, apply approximately 250 µl of methylene chloride to the cut off edges of the conical tube to bind the conical tube to the 150 mm lid.
- Seal the 150 mm lid onto the 150 mm dish by applying a 1 mm thick bead of silicon rubber sealant around the inner edge of the top Petri dish.
2. Sterilization of Shear Rings
- Pipet approximately 10 ml of phosphate buffered saline into the newly formed shear ring through the 15 ml conical tube port. Swirl around to remove any debris inside the shear ring. Remove the phosphate buffered saline with a Pasteur pipet/vacuum aspirator.
- Repeat the previous step until debris is removed.
- To sterilize the shear ring, use a combination of a 70% ethanol rinse with UV irradiation.
- Unscrew the vented cap, pipet approximately 10 ml of 70% ethanol through the access port, and screw the cap back on. Rotate and flip over the shear ring multiple times, ensuring that the inside of the shear ring is thoroughly washed.
- Under a fume hood, aspirate out excess 70% ethanol. With a spray bottle, thoroughly spray the cap and access port with 70% ethanol.
- Place the shear ring and cap under UV radiation within the tissue culture hood until completely dry.
- Once dry, screw the cap back on and store sterilized shear rings at room temperature until used in cell culture plating.
3. Shear Stress Studies
- Plate endothelial cells in sterilized shear rings following the standard cell culture protocol typically using a 1:4 split ratio for transformed endothelial cell lines.
NOTE: Rat retinal microvascular endothelial cells were obtained commercially. Human brain endothelial cells (hCMEC/D3) cells were provided as a generous gift from Dr. Pierre-Oliver Couraud (Inserm, France) and were cultured in complete endothelial basal medium (EBM). - Allow cultures to reach confluence prior to initiation of flow studies. Use 30 ml of tissue culture media (10% fetal calf serum, Dulbecco's modified Eagle's medium with 1% penicillin/streptomycin/amphotericin). Change cell culture medium every 3 days and maintain cells at 37 °C with 7.5% CO2 and 92.5% room air.
- Place the orbital shaker in the incubator.
Note: Orbital shakers are typically heavy (>10 kg). Place the orbital shaker on the bottom shelf of the incubator to minimize shelf stress and vibration. - Place the experimental shear rings on the orbital shaker. Place static control group shear rings inside the same incubator.
- Estimate the maximum shear stress within the shear ring with the equation
 where
where  is the radius of rotation for the orbital shaker (in cm),
is the radius of rotation for the orbital shaker (in cm),  is the viscosity of the medium (in poise),
is the viscosity of the medium (in poise),  is the density of the medium (in g/ml), and
is the density of the medium (in g/ml), and  is the frequency of rotation (in rotation/sec)24.
is the frequency of rotation (in rotation/sec)24. - Initiate the rotation setting on the orbital shaker to the desired rpm (e.g., 90 rpm), leaving the shear rings on the shaker for the desired duration of shear stress application (e.g., 72 hr).
NOTE: Orbital shakers can produce heat, so the incubator temperature should be monitored initially to ensure 37 °C is maintained throughout the duration of the experiment. Alternatively, shear rings can be transferred into environmental chambers (e.g., modular incubator chamber) and then placed within a rotating incubator. - After cell layers have been exposed to shear for the desired length of time, remove shear rings from the incubator. Pull off the shear ring lid and retrieve cells and/or media for examining desired endpoint analyses (e.g., Western blotting, fluorescence activated cell sorting, etc.)25.
Here we present representative results from both hCMEC/D3 brain endothelial cell and rat retinal microvascular endothelial cell monolayers, cultured in shear rings.
After allowing hCMEC/D3 brain endothelial cell monolayers to grow to confluence in complete EBM, the shear rings were placed on an orbital shaker for 72 hours. Using the equation from step 3.5, the calculated maximal shear stress  was approximately 2.8 dynes/cm2 (with parameters = 0.95 cm, = 0.0101 poise, = 0.9973 g/ml24, = 1.5 rotations/second). We have found that these endothelial cell monolayers sometimes exhibit alignment in parallel with the direction of the periodic flow (Figure 3), although this is not uniformly observed because cell layers usually have excellent adhesion to the shear ring surface throughout the study.
was approximately 2.8 dynes/cm2 (with parameters = 0.95 cm, = 0.0101 poise, = 0.9973 g/ml24, = 1.5 rotations/second). We have found that these endothelial cell monolayers sometimes exhibit alignment in parallel with the direction of the periodic flow (Figure 3), although this is not uniformly observed because cell layers usually have excellent adhesion to the shear ring surface throughout the study.
After allowing the rat retinal microvascular endothelial cell monolayers to grow to confluence in complete rat endothelial cell medium, including EGF/VEGF growth factor supplements, the shear rings were placed on an orbital shaker for 72 hours. Using the equation from step 3.5, the calculated maximal shear stress was approximately 12 dynes/cm2 (with parameters = 0.95 cm, = 0.0101 poise, = 0.9973 g/ml24, = 4 rotations/second). Figure 4 shows that compared to the loading control of β-actin, there was a large and significant loss of platelet endothelial cell adhesion molecule (PECAM-1/CD31) from the endothelial cell surface.
PECAM-1 is an integral membrane protein which is a member of the immunoglobulin (Ig)-superfamily that contains an immunoreceptor tyrosine-dependent inhibitory motif or 'ITIM'26. PECAM-1 is not only expressed on endothelial cells but is also found on hematopoietic cells. PECAM-1 plays significant roles in endothelial cell-cell adhesion, leukocyte junctional transmigration, cell signaling, and importantly, mechano-transduction of shear stress. PECAM-1's role in sensing shear stress is critical for the functions of vascular endothelial cells and homeostasis17. When endothelial monolayers are exposed to shear stress, PECAM-1 responds directly to the mechanical force being exerted on it by altering its tyrosine phosphorylation, and subsequent activation of the ERK1/2 signaling cascade27. Furthermore, the PECAM-1-eNOS complex association is interrupted by disturbances in shear stress28. Therefore, PECAM-1 enables vascular endothelial cells to sense changes in fluid shear stress forces which can lead to reactive dilation of the vessel wall29.
These data support this model showing that endothelial cells respond to exposure to periodic, unidirectional fluid shear by down-regulating an important junctional and adhesive determinant, which mediates intercellular contact as well as transvascular cell exchange.

Figure 3. Brain endothelial cell morphology in a shear ring. The appearance of hCMEC/D3endothelial cell monolayers in shear rings following 48 hours of periodic fluid shear or static exposure. Alignment of cultures is not always observed parallel to flow direction (shown by arrow). Please click here to view a larger version of this figure.

Figure 4. Cell responses to periodic shear. Rat retinal microvascular endothelial cell monolayers cultured in shear rings showed a reduction in PECAM1/CD31 relative to β-actin (n=3 each, students unpaired t-test, *-p<0.05, error bars refer to standard deviation), following 72 hours of periodic fluid shear in shear rings.
