

Summary
الاستفادة من Cas9 ستنشئ ribonucleoprotein المعقدة (رنب) وسيلة قوية لتحرير الجينوم الدقيقة، وتتسم بالكفاءة. هنا، نسلط الضوء على فائدتها عبر مجموعة واسعة نطاق من الخلايا والكائنات الحية، بما في ذلك الخلايا البشرية الأساسية والكلاسيكية على حد سواء والناشئة نموذج الكائنات الحية.
Abstract
تحرير الجينوم التوكسينات الخاصة بالموقع مع كريسبر (متفاوت المسافات بانتظام إينتيرسباسيد قصيرة المتناوب يكرر)-نظم (المرتبطة كريسبر) Cas أصبحت بسرعة شائعة بين الباحثين اتباع طائفة واسعة من المسائل البيولوجية. في أغلب الأحيان توظيف المستخدمين البروتين Cas9 المستمدة من العقدية المقيّحة في مجمع مع دليل أعيدت برمجتها بسهولة بالحمض النووي الريبي (جرنة). يتم عرض هذه المكونات في الخلايا، ومن خلال قاعدة الاقتران مع منطقة مكملة لجينوم الحمض النووي (dsDNA) مزدوجة-الذين تقطعت بهم السبل، كليافيس الإنزيم كل خيوط لإنشاء فاصل مزدوج-ستراند (جهاز تسوية المنازعات). يؤدي الإصلاح اللاحقة إلى الإدراج عشوائية أو حذف الأحداث (إينديلس) أو إدماج الحمض النووي المجرب المتوفرة في موقع فاصل.
استخدام المنقي واحد-دليل الحمض النووي الريبي و Cas9 بروتين، ستنشئ لتشكل رنب وتسليمها مباشرة إلى الخلايا، ونهج قوية لتحقيق تحرير الجينات ذات كفاءة عالية. تحرير رنب خاصة ويعزز معدل الجينات الإدراج، نتيجة التي غالباً ما تكون صعبة لتحقيق. بالمقارنة مع التسليم عن طريق بلازميد، استمرار أقصر من رنب Cas9 داخل الخلية يؤدي إلى أقل خارج الهدف أحداث.
على الرغم من مزاياها، العديد من المستخدمين العاديين لتحرير كريسبر الجينات أقل إلماما بهذا الأسلوب. إلى انخفاض الحاجز للدخول، فإننا مخطط بروتوكولات مفصلة لتنفيذ استراتيجية رنب في مجموعة من السياقات، إبراز فوائد متميزة وتطبيقات متنوعة. نحن تغطي التحرير في نوعين من الخلايا البشرية الأولية والخلايا T والخلايا الجذعية المكونة للدم/السلف (هسبكس). ونحن أيضا إظهار كيف رنب Cas9 التحرير يتيح التلاعب الوراثية السطحية بأكمله الكائنات الحية، بما في ذلك النموذج الكلاسيكي الدودة ايليجانس كاينورهابديتيس والأكثر مؤخرا عرض نموذج القشريات، هاواينسيس بارهيالي.
Introduction
fThe كريسبر-Cas9 نظام يسمح للعلماء لتغيير المناطق المستهدفة من أي الجينوم1. ثورة البحوث الأساسية هذه التكنولوجيا سريعة وغير مكلفة ووعود لجعل تأثير عميق على تطوير علاجات المرض شخصية، الزراعة الدقيقة، ويتجاوز2. تحرير كريسبر أداة الديمقراطية وتنفيذ النظام في مختبر جديد يتطلب لا خبرة خاصة في مهارات البيولوجيا الجزيئية الأساسية فقط، وهندسة الجينوم. يمكن الآن أن يدرس الباحثون الكائنات العضوية المستعصية سابقا مع عدد قليل من الوسائل البديلة للمعالجة الجينية3،4. في السنوات الخمس الماضية وحدها، استخدمت كريسبر الجينوم التحرير لمهندس ما يزيد على 200 مختلفة من الفقاريات واللافقاريات والنبات والأنواع الميكروبية.
مقتبس من مسار الدفاع بدائية كريسبر، العناصر الأساسية المطلوبة لتحرير الجينوم الخاصة بالموقع هي البروتين Cas9، عادة من س. المقيّحة والأمثل كودون مع إشارة تعريب نووية المضافة (NLS)، والمتخصصة الجيش الملكي النيبالي دليل5،6. ولو لم تناقش هنا، كما يمكن استخدام أورثولوجويس Cas9 أو اندونوكليسيس كريسبر الأخرى. جرنة التي تحدث بشكل طبيعي ويتكون من قطعتين يدون كل على حدة، و "الجيش الملكي النيبالي كريسبر" (كرنا) وعبر-تفعيل كرنا (تراكرنا)7. يمكن أن تنصهر فيها هذه الكشف في نسخة واحدة، المعروفة باسم الجيش الملكي النيبالي (سجرنا) دليل واحد8. معظم الجينوم المحررين اختيار سجرنا مبسطة9، على الرغم من أن الدليل المزدوج أيضا يستخدم بانتظام10،11. المجربون اختيار 20-النوكليوتيدات (nt) المجينية الحمض النووي هدفا، وضمان أن يقع بجوار قصيرة توقيع تراخيص المطلوبة للاعتراف Cas9، تسمى فكرة مجاورة بروتوسباسير (بام)، وتصميم جرنة يحتوي على تسلسل مكملة12 .
مرة واحدة داخل الخلية، يحدد رنب معقدة هدفها الجينوم وأزواج قاعدة جرنة مع الحمض النووي التكميلية حبلا ثم كليفس الإنزيم كسر كل خيوط الحمض النووي لتوليد مزدوج-حبلا2. آلات إصلاح الخلية بإصلاح جهاز تسوية المنازعات بواحد من اثنين على الأقل من طرق: عن طريق المسار (نج) الانضمام إلى نهاية عرضه للخطأ غير-مثلى أو إصلاح موجه التماثل (HDR)، الذي يشمل بسلاسة الحمض النووي الذي يحتوي على 'الأسلحة' من التماثل إلى أي من الجانبين للكسر. مسار إصلاح السابق عادة يؤدي إلى تشكيل إينديل والجينات ما ترتب عليها من اضطراب، بينما هذا الأخير يسمح المجربون إدراج أو تغيير تسلسل الحمض النووي1.
كفاءة ودقة التحرير تعتمد على الوسائل التي تدخل Cas9 وجرنة في الخلية. قد يتم تسليم هذه العناصر إلى الخلايا المستزرعة، الأجنة، أو الكائنات في النموذج من الأحماض النووية أو أنها ستنشئ رنب معقدة13،،من1415. وتشمل طرق التسليم على أساس الحمض النووي المشترك توصيل الفيروسية، تعداء، أو انهانسر مرناً أو بلازميد الحمض النووي. دليل الحمض النووي الريبي والبروتين Cas9 ثم يتم إنتاجها داخل الخلية وهم المنتسبين لتشكيل مجموعة معقدة.
التسليم المباشر من رنب يتطلب تنقية منفصلة للبروتين Cas9 ودليل الحمض النووي الريبي. يمكن أن يتم ذلك داخل المنظمة، أو يمكن شراؤها بالبروتين وسجرنا من واحدة من العديد من الموردين التجاريين. بمجرد الحصول عليها، Cas9 وجرنة مختلطة بشكل مجمع رنب انزيماتيكالي المختصة وأدخلت إلى الخلايا عن طريق الحقن المباشر في البيض/أجنة مخصبة أو على أساس المادة الدهنية تعداء16انهانسر. التقرير الأول لتحرير رنب تشارك الحقن في الغدد التناسلية ايليجانس جيم- 17. Microinjection لا يزال الوسيلة المفضلة لإدخال رنب في الأجنة والكائنات الحية كلها، لو انهانسر فعالة قد تجلى في الماوس18،19 والفئران20 الأجنة. وصف بروتوكولات لمباشرة ضخ رنب C. ايليجانس الغدد التناسلية والأجنة هاواينسيس P. ونوصي بنوع متخصصة من انهانسر لتسليم رنب عند تحرير الخلايا البشرية الأساسية. هذا الأسلوب، نوكليوفيكشن، ينطوي على برامج انهانسر الأمثل والحلول الخاصة بنوع الخلية ويسمح رنب بالدخول السيتوبلازم و نواة21.
تحرير الجينوم مع رنب يوفر العديد من المزايا المتميزة. لأنه قبل تجميع البروتين وعناصر الجيش الملكي النيبالي، ويمكن ضمان الجودة قبل التسليم، تحرير رنب يتجنب العديد من المخاطر المقترنة بالتسليم على أساس الحمض النووي. هي: ليس هناك أي خطر من اندماج الحمض النووي Cas9-ترميز الجينوم المضيف، مرناً لم تتعرض للتدهور، وأنها تلتف مشاكل في فيفو جرنة أو بروتين التعبير وقابلة للطي، ورابطة،من2223. علاوة على ذلك، باستخدام رنب يؤدي إلى انخفاض السمية والأحداث خارج الهدف أقل بكثير من التعبير بلازميد القائم، نتيجة أقصر فترة نصف العمر رنب داخل الخلية24،25،،من2627.
وأخيراً، تحرير رنب واضح يؤدي إلى ارتفاع معدلات التحرير في مجموعة متنوعة من خطوط الخلايا البشرية، الخلايا الأولية مثل الخلايا الليفية، الخلايا الجذعية الجنينية (بتوليدا)، التي يسببها الخلايا الجذعية pluripotent (إيسبكس)، هسبكس، والخلايا T2416،، 25،،من2627،،من2829؛ في اللافقاريات بما في ذلك هاواينسيس P. C. ايليجانسوذباب الفاكهة3،،من1730؛ في الأنواع الفقارية مثل الزرد، والفئران والجرذان31،32؛ في زراعة الأنواع بما في ذلك نبات، والتبغ، والخس، الأرز والعنب، أبل، الذرة والقمح33،34،،من3536؛ وفي تشلاميدوموناس و Penicillium والمبيضات الأنواع37،،من3839. تواتر تشكيل إينديل يمكن أن تكون أعلى عند استخدام رنب مقارنة بإيصال بلازميد، والإدراج HDR بوساطة الحمض النووي يمكن أن يكون أسهل لتحقيق25،،من2729.
البروتوكول هو موضح هنا يستخدم رنب Cas9، وهو أسلوب فعال، وسهولة التكيف واضحة لتنطبق على مجموعة متنوعة من النظم البيولوجية40،41، لا سيما في الخلايا التي يصعب إلا بالعمل وفي الكائنات الحية دون نظم راسخة للمعالجة الجينية دقيقة. نبدأ بوصف كيفية الحصول على تصميم وتجميع رنب Cas9 قبل تغطي استخدامها عبر نموذج مختلف أنواع الخلايا والكائنات الحية. الخلايا الجذعية المكونة للدم/السلف (هسبكس) والخلايا T يتم تحريرها باستخدام نفس الأسلوب، نوكليوفيكشن، حيث أنها تغطي معا في الخطوتين 2 و 3 من هذا البروتوكول. تحرير إجراءات ل C. ايليجانس موصوفة في الخطوتين 4 و 5، وف. تحرير هاواينسيس مشمول في الخطوتين 6 و 7. أخيرا، حيث يمكن تقييم نجاح تجربة تحرير الجينات في أي كائن حي بالتسلسل الوراثي، يرد سوبستيبس تصف أساليب تحليل ممكن لجميع الخلايا والكائنات الموصوفة في البروتوكول في الخطوة 8.
Protocol
1-رنب الجمعية
-
تصميم التجربة في وقت مبكر، الحصول على جميع عناصر الجيش الملكي النيبالي، والحمض النووي والبروتين قبل الموعد المحدد. كمرور أولى، حاول أحد عناصر إيجابية المدرجة في الجدول 1 واستخدام الكواشف التجارية المبينة في "الجدول للمواد" لضمان تصميم تجريبي يمكن الاعتماد عليها وسلامة المواد. للحصول على نصائح إضافية حول تخطيط تجربة جديدة تحرير الجينوم، انظر الورقات على هذا الموضوع12،،من4243.
ملاحظة: بمجرد تجميعها كما هو موضح في الخطوات اللاحقة، رنبس معدّة مسبقاً قد تكون مخزنة في-80 درجة مئوية.- بعد اختيار الجينات التي تستهدف، استخدم إحدى الأدوات على الإنترنت مجاناً لتصميم أمثل جرنة45،44،46،،من4748. من المؤكد أن استهداف إكسون إذا على أمل أن تولد بالضربة قاضية.
ملاحظة: سوف تساعد هذه الأدوات لتحديد موقع هدف جوار المقيّحة س. بام تسلسل ودرجة عالية الجودة، وانخفاض نقاط خارج الهدف. - تنقية البروتين المقيّحة س. Cas9 من خلال أساليب نشر8، أو الشراء من مورد تجاري.
- إعداد المخزن مؤقت Cas9 نموذجية لإضعاف الجيش الملكي النيبالي، وإعداد رنب، وتخزين البروتين، الذي يحتوي على 20 مم من حبيس الرقم الهيدروجيني 7.5، 150 مم بوكل ووالغليسيرول 10% 1 مم من تسيب. دائماً استخدام مياه خالية من نوكلاس في المخازن المؤقتة التي سيتم استخدامها ريسوسبيند أو تخفف من الحمض النووي الريبي للحيلولة دون تدهور.
- إنتاج دليل الحمض النووي الريبي (تراكرنا وكرنا أو سجرنا) من خلال نسخ في المختبر استخدام أساليب المنشورة، أو شرائه من حمض النووي توليف شركة17،،من2149، 50 , 51.
- إذا كان إدخال جين، توليف أو شراء إحدى الجهات مانحة قالب الحمض النووي.
- تخزين البروتين والحمض النووي الريبي مختبرين في-80 درجة مئوية وذوبان الجليد على الجليد مباشرة قبل الاستخدام.
ملاحظة: كل تجميد أذاب قليلاً يخفض الكفاءة. بروتوكولات مفصلة، والوصول المفتوح ل تنقية Cas952 والنسخ في المختبر من سجرناس53 متوفرة في أماكن أخرى.
- بعد اختيار الجينات التي تستهدف، استخدم إحدى الأدوات على الإنترنت مجاناً لتصميم أمثل جرنة45،44،46،،من4748. من المؤكد أن استهداف إكسون إذا على أمل أن تولد بالضربة قاضية.
- إذا كان يعمل مع C. ايليجانس، تخطي إلى الخطوة 1.5. بروتوكول هاواينسيس P. ، انتقل إلى الخطوة 1، 6. في حالة استخدام سجرنا، انتقل إلى الخطوة 1، 4. تابع إلى الخطوة 1.3 لتجميع جرنا لتحرير الخلية الأولية.
-
تجميع جرنة بخلط مقادير اكويمولار تراكرنا وكرنا. جعل 100 ميليلتر من الأسهم جرنا 80 ميكرومتر، لتجارب التحرير الجينوم حوالي 50.
- احتضان جرنة في 37 درجة مئوية لمدة 30 دقيقة وثم السماح لها لتبرد ببطء إلى درجة حرارة الغرفة.
-
رنب الإعدادية لتحرير الخلية هسبك وترينيداد: تجميع رنب معقدة عن طريق خلط كمية مولى س 1-2 من جرنة إلى بمول 200 من البروتين Cas9 في إجمالي حجم 10 ميليلتر. ببطء شديد، إضافة Cas9 مركزة جرنة (تضعف مسبقاً في المخزن المؤقت Cas9) لحوالي 30 ثانية ، مما يجعل الدوائر السريعة مع الماصة، يصل تركيز Cas9 النهائي إلى 20 ميكرومتر.
- إعداد الترعة انهانسر.
ملاحظة: هذا البروتوكول محددة للنظام التجاري المشار إليها في الجدول للمواد، ولكن تحرير رنب يمكن أيضا أن يتحقق مع غيرها من الأجهزة انهانسر. - إضافة 5 ميليلتر (100 بمولس، والخلايا T) أو 10 ميليلتر (200 بمول، هسبكس) من رنب لكل ومبومو.
- إذا كان إدخال الحمض النووي الجديد بدلاً من صنع بالضربة قاضية، إضافة 1 ميليلتر من 100 ميكرومتر (بمول 100) اليغنوكليوتيد واحد-الذين تقطعت بهم السبل المانحة الحمض النووي (سودن)25،54،55 إلى الترعة أو آبار اللوحة.
- تخطي إلى الخطوة 2 للإرشادات التالية في تحرير الخلية الابتدائية البروتوكول.
- إعداد الترعة انهانسر.
-
رنب الإعدادية لتحرير C. ايليجانس : تجميع رنب المعقدة عن طريق إضافة الكواشف التالية بغية تهيئة وحدة تخزين نهائي من 20 ميليلتر (تتم الإشارة إلى تركيزات نهائية في أقواس): Cas9 (2 ميكرومتر)، حبيس الرقم الهيدروجيني 7.5 (10 ميكرومتر)، بوكل (115 ميكرومتر)، كرنا (12 ميكرومتر) ، تراكرنا (40 ميكرومتر)، وإصلاح قوالب إذا لزم الأمر (0.5 ميكرومتر ssDNA أو ما يصل إلى 350 نانوغرام/ميليلتر دسدنا).
ملاحظة: كفاءة إصلاح Cas9 بوساطة جهاز تسوية المنازعات-قالب يتناسب مع تركيز بناء إصلاح دسدنا؛ وهكذا، أعلى تركيز لإصلاح القالب، أكثر كفاءة إصلاح القوالب. ومع ذلك، ثبت حقن أمزجة المحتوية على أكثر من 350 نانوغرام/ميليلتر من دسدنا للحد من إمكانية حقن الديدان. وهكذا، فمن الأفضل استخدام ما يصل إلى، ولكن لا يزيد عن 350 نانوغرام/ميليلتر من دسدنا في مزيج لتعظيم كفاءة إصلاح مع التقليل من فتكه.- إضافة كرناس متعددة لاستهداف مواقع متعددة في وقت واحد، حسب الحاجة، لفحص co-كريسبر/المشارك-conversion النهج المذكور في الخطوة 5، 4. عند إضافة أكثر من كرنا، إضافة كل تسلسلياً إلى مزيج الرئيسي.
ملاحظة: مقدار كل كرنا لا تحتاج إلى أن يكون هو نفسه، وحتى مضاعفة التركيز الكلي كرناس في مزيج الرئيسي دون تغيير تركيز Cas9 لا يبدو أن تتداخل مع تواتر الطفرات في موضع محدد. وترد أمثلة بالتفصيل في Paix et al. 56. - مزيج من بيبيتينج وتدور الحل رنب في 16,000 ز خ لل 5 s للتأكد من أن الحل هو تجمع في الجزء السفلي من الأنبوب.
- احتضان الحل عند 37 درجة مئوية عن 15 مترا.
- الطرد المركزي العينة في 16,000 س ز لمدة 1 دقيقة بيليه أي الجسيمات التي يمكن أن تسد إبرة microinjection رقيقة--بالملل. استخدام المادة طافية في الخطوات اللاحقة.
- انتقل إلى الخطوة 4 للفترة المتبقية من بروتوكول C. ايليجانس .
- إضافة كرناس متعددة لاستهداف مواقع متعددة في وقت واحد، حسب الحاجة، لفحص co-كريسبر/المشارك-conversion النهج المذكور في الخطوة 5، 4. عند إضافة أكثر من كرنا، إضافة كل تسلسلياً إلى مزيج الرئيسي.
-
رنب الإعدادية لتحرير هاواينسيس P. : إعداد مختبرين Cas9 تستخدم مرة واحدة عن طريق إضعاف لهم مع المياه خالية من نوكلياسي والفينول الحمراء (لتصور الحقن) إلى نهائي تركز ميكرومتر 6.25 الأحمر الفينول Cas9 و 0.15 في المائة.
- تجميع رنب المعقدة عن طريق خلط 2-5 x الزائدة المولى من جرنة للبروتين Cas9 في إجمالي حجم 6 ميليلتر. إضافة 12 بمول من Cas9 إلى جرنة، يصل تركيز Cas9 النهائي إلى 2 ميكرومتر، وتركيز جرنة على 4-8 ميكرومتر، وتركيز الفينول الحمراء إلى 0.05%.
- احتضان هذا الخليط في درجة حرارة الغرفة لمدة 10 دقيقة لمجمع في رنب.
- تخطي إلى الخطوة 6 للإرشادات التالية في التحرير هاواينسيس P. البروتوكول.
2-الخلية الثقافة وإعداد
ملاحظة: إجراء الخطوات 2.1.1 إلى 3.3.3 في سلامة بيولوجية مجلس الوزراء.
-
شراء cryopreserved الإنسان تعبئة الدم المحيطي CD34+ هسبكس من مورد.
- ذوبان الجليد ~ 1 x106 هسبكس في مياه 37 درجة مئوية حمام لمدة 3 دقائق وتحويلها إلى أنبوب مخروطي 15 مل. إضافة 10 مل وسيلة توسيع المصل خالية من مصدر تجاري وتدور المخلوط في ز 100 x دقيقة 10 إزالة المادة طافية وريسوسبيند الخلايا في 2 مل سفيم المكملة. لوحة الخلايا في لوحات 6-جيدا والثقافة لهم في حاضنة 37 درجة مئوية أجل ح 24-48 قبل انهانسر رنب.
- عد الخلايا مع هيموسيتوميتير ونقل العدد الإجمالي من هسبكس (هسبكس 150,000 200,000 الواحد ومبومو أن تكون اليكتروبوراتيد) يلزم أنبوب الطرد مركزي.
- تدور الأنبوب في 100 غ س لمدة 10 دقيقة بيليه الخلايا.
-
شراء البشرية CD4 الأولية+ تي الخلايا من بائع أو عزلها من دم كل البشر ب الطرد المركزي التدرج كثافة29.
- قبل تنشيط خلية T، قبل معطف لوحات الثقافة 48-جيدا مع αCD3 (UCHT1) و αCD28 (CD28.2). معطف اللوحات مع 500 ميليلتر من 10 ميكروغرام/مل αCD3 و 10 ميكروغرام/مل αCD28 في برنامج تلفزيوني على الأقل 2 ح في 37 درجة مئوية.
ملاحظة: لبعض المكاني، نج يمكن أن يتحقق دون التحفيز المسبق، ولكن بما في ذلك هذه الخطوة يزيد من فعاليتها. - خلايا الثقافة T عن 48 ساعة عند 37 درجة مئوية على ألواح جسم زمنياً αCD3/αCD28 في متوسط كاملة ربمي [RPMI 1640 تستكمل مع 5 مم حبيس، 2 مم بديل تجاري للأم الجلوتامين، 50 ميكروغرام/مل من البنسلين/ستربتوميسين، 50 ميكرون من 2-mercaptoethanol، 5 ملم من غير الأحماض الأمينية الأساسية، 5 ملم بيروفات صوديوم، و 10% (المجلد/المجلد) FBS]. الثقافة تي الخلايا في كثافة الخلايا T 2,000,000 في 500 ميليلتر لوسائل الإعلام كل من لوحة 48-جيدا جيدا.
- حساب تي الخلايا باستخدام هيموسيتوميتير ونقل تجربة العدد الإجمالي لخلايا تي ضرورية انهانسر (100,000 1,000,000 تي الخلايا الواحدة ومبومو أن اليكتروبوراتيد) إلى أنبوب الطرد مركزي.
- تدور الأنبوب في ز x 90 دقيقة 8 بيليه الخلايا. إذا كانت الخلايا كثافة مفصولة بالتدرج خلال يومين، تدور لهم في 200 غ س ل 8 دقيقة.
- قبل تنشيط خلية T، قبل معطف لوحات الثقافة 48-جيدا مع αCD3 (UCHT1) و αCD28 (CD28.2). معطف اللوحات مع 500 ميليلتر من 10 ميكروغرام/مل αCD3 و 10 ميكروغرام/مل αCD28 في برنامج تلفزيوني على الأقل 2 ح في 37 درجة مئوية.
-
لكل أنواع الخلايا، نضح المادة طافية مع ماصة/فراغ، إزالة أي فقاعات.
- ريسوسبيند الخلايا مع 20 ميليلتر من المخزن المؤقت انهانسر بلطف كل ومبومو.
- إضافة 20 ميليلتر من الخلايا (هسبكس 150,000 200,000 أو خلايا تي 100,000-1,000,000) لكل ومبومو، الذي يحتوي بالفعل على 10 ميليلتر رنب، ومزيج جيد من قبل بيبيتينج صعودا وهبوطاً دون خلق الفقاعات.
3-رنب انهانسر
- اليكتروبوراتي الترعة بعد وضعها في نوكليوفيكتور. هسبكس، استخدم التعليمات البرمجية نبض ER100. للخلايا T، استخدم التعليمات البرمجية نبض EH-115.
-
هسبكس فقط: إضافة 100 ميليلتر من متوسطة سفيم المكملة (درجة حرارة إلى 37 درجة مئوية) لكل ومبومو فور انهانسر وترك الخلايا استرداد ل 10-15 دقيقة
- نقل الخلايا إلى ثقافة لهم في جولة 96-جيدا-قاع لوحة وإضافة ميليلتر 100 إضافية من متوسطة سفيم المكملة لمدة 24 ساعة.
- تغييرها إلى وسيلة سفيم تستكمل طازجة واحتضانها لهم ح 24-72 إضافية.
- إزالة الخلايا للتنميط لهم ح 48-96 وظيفة-انهانسر. تدور الخلايا في 300 غرام x لمدة 5 دقائق وإزالة المادة طافية قبل البدء في استخراج الحمض النووي (الخطوة 8، 2).
-
خلايا T فقط: إضافة 80 ميليلتر من ربمي إكمال ثقافة وسائل الإعلام قبل تسخينها إلى 37 درجة مئوية من الخزان إلى كل ومبومو أو جيدا، باستخدام ماصة متعدد القنوات (إذا لزم الأمر).
- احتضان لهم في 37 درجة مئوية لمدة 15 دقيقة.
- إضافة الوسائط المناسبة، الأجسام المضادة، السيتوكينات، إلخ إلى plate(s) الوجهة والحارة لهم مسبقاً في حاضنة 37 درجة مئوية.
- نقل 107 ميليلتر من الخلايا اليكتروبوراتيد من الآبار إلى لوحة 96-بئر مستديرة القاع استخدام ماصة متعدد القنوات (إذا لزم الأمر).
- للحصول على مزيد من المعلومات حول تقييم نتائج التحرير، انتقل إلى الخطوة 8.
4-إعداد C. ايليجانس
-
قبل microinjection يوم 1: إعداد منصات [اغروس] microinjection.
- جعل حلاً [اغروس] 3% (w/v) في المياه بإضافة [اغروس] إلى المياه وتحقيق الحل الذي يغلي على طبق ساخن أو في فرن ميكروويف.
- ترتيب 24 مم × 50 مم × 1.5 ملم الزجاج غطاء الشرائح على طاولة واستخدام زجاج ماصة باستور لوضع قطره محلول [اغروس] على الشريحة صغيرة (~ 15 ميليلتر). بسرعة تسطيح القطرة [اغروس] بوضع ساترة آخر في أعلى. يسمح [اغروس] ترسيخ ثم قم بإزالة واحدة من كوفيرسليبس.
- ترك الوجه [اغروس] المغلفة ساترة لأعلى على الطاولة بين عشية وضحاها الجاف. بعد 24 ساعة، تخزين منصات [اغروس] في حاوية نظيفة وجافة.
ملاحظة: هذه يمكن استخدامها إلى أجل غير مسمى.
- سحب الإبر microinjection: استخدام زجاج البورسليكات الشعيرات الدموية مع خيوط (القطر الخارجي الداخلية 1.0 مم وقطرها ملم 0.58)، سحب الإبر على أساس ميلو والنار57 وسائر الموارد58. الإبر يمكن استخدامها مباشرة أو يمكن تخزينها في حاوية نظيفة وجافة، استعدت بالطين ويدعم.
- للحفاظ على الديدان، تعد من وسائط نمو ديدان أسطوانية (NGM) أجار تدفقت لوحات بيتري، ورصدت بالبكتيريا OP50 (لبروتوكولات قياسية C. ايليجانس الصيانة ووصفات لوسائط النمو، انظر ستيرناجلي59).
- المرحلة الديدان ل microinjection: ح 12-24 قبل microinjection، اختيار المنحرفين نظموا L4 لصفيحة نغ-أجار جديدة مع البكتيريا OP50 واحتضانها لهم بين عشية وضحاها في 20 درجة مئوية. لكل مزيج المستهدفة/حقن Cas9، بيك الديدان ~ 30 لوحة.
-
يوم microinjection: تحميل microinjection سحبت الإبرة مع الحل رنب طافية إعدادها في الخطوة 1، 5-
- "الماصة؛" المادة طافية من الخطوة 1.5.4 إلى سحبت الماصة الشعرية والردم الحل من الماصة الشعرية في إبرة microinjection استعداد (تحميل عموما أقل من 0.1 ميليلتر).
- جبل الإبرة محملة على جهاز microinjection، تعلق على ميكرومانيبولاتور. تعيين ضغط أجهزة الحقن إلى 250 كيلو باسكال وضغط التوازن إلى 25 الجيش الشعبي الكوري.
-
كسر مرة أخرى نصيحة إبرة المحملة لتوليد على حافة إبرة حادة. مكان 15 ملم × 15 ملم × 1.5 مم ساترة مربعة على الجزء العلوي 24 مم × 50 مم × 1.5 مم ساترة.
- تراكب حافة واحدة من ساترة مربعة مع النفط الهالوكربون 700.
- ضع الإبرة في مجال النفط، على حافة ساترة مربع 15 ملم.
- باستخدام يد لتوجيه المرحلة مجهر وساترة، وفرشاة الشريحة أعلى وعلى طول حافة الإبرة بينما الاكتئاب الدواسة أو الزر الحقن. كسر إبرة التلميح مرة أخرى، زيادة تدفق السائل من الإبرة. تحقيق معدل تدفق أمثل عن طريق جعل حقن خلط تدفق على طول حافة الإبرة، تشكيل فقاعة ~ 1/s.
- تؤكد أن الديدان L4 اختار ح 12-24 قبل microinjection تنمويا نظموا صغار البالغين في يوم الحقن. اختيار الديدان الكبار الصغار لصفيحة نغ-أجار يفتقر للبكتيريا OP50 والسماح لهم بالزحف حول لمدة 5 دقائق. وهذا يقلل من كمية البكتيريا التي تنقل إلى منصة الحقن، التقليل من قباقيب إبرة.
- وضع [اغروس] حقن لوحة/ساترة على نطاق تشريح. استخدام اختيار دودة، إرساء مسار صغير للنفط الهالوكربون على طول حافة واحدة من لوحة المفاتيح.
-
استخدام انتقاء دودة المغلفة في النفط، رفع الديدان عدة قبالة لوحة أجار نغ وإلى المسار من النفط. مع غرامة شعر يلحق ماصة، مثل الخط الطولي جفن أو القط، ضع الديدان في دفع موازية، بلطف الديدان إلى لوحة المفاتيح [اغروس]. حتى مريحة مع الإجراء microinjection، جبل فقط وحقن دودة واحدة في وقت واحد.
ملاحظة: سوف الفتيل [اغروس] الجافة الرطوبة من الديدان، مما يسبب لهم التمسك بلوحة المفاتيح. ونتيجة لذلك، واحد يجب أن تعمل بسرعة كما يمكن ديسيككاتي الديدان.- مرة واحدة في موقف وتعلق على لوحة المفاتيح، تراكب الديدان مع آخر قطرات النفط الهالوكربون (~ 20 ميليلتر) من طرف الدودة بيك.
5- C. ايليجانس الغدد التناسلية Microinjection مع رنبس والرعاية ما بعد الحقن
ملاحظة: بروتوكول microinjection هو مقتبس من ميلو والنار57ووصف بالتفصيل في مكان آخر60،61.
-
ضع ساترة مع الديدان المحملة على الحقن المجهر. تحت تكبير منخفضة (الهدف X 5، 10 X العين)، ضع الديدان إبرة حقن عمودي.
- التبديل إلى تكبير عالية (40 X الهدف، 10 X العين)، قم بتغيير موضع الإبرة المتاخمة الغدد التناسلية الذراع المقابلة للمنطقة قرب النوى في منتصف-للتأخر-باتشيتيني.
- تستخدم في ميكرومانيبولاتور، نقل الإبرة ضد الدودة، كئيبة بشرة قليلاً. ثم، مع يد واحدة، اضغط على الجانب من مرحلة مجهر هزة الإبرة عن طريق بشرة. كساد الدواسة أو الزر حقن ببطء ملء ذراع الغدد التناسلية مع مزيج حقن وإزالة الإبرة.
- كرر هذه الخطوة مع الذراع الغدد التناسلية الأخرى.
-
حالما يتم حقن الديدان، إزالة لوحة ساترة/[اغروس] ووضعه تحت مجهر تشريح.
- استخدام ماصة شعرية سحبت، تحل محل النفط من الديدان التي بيبيتينج مؤقت M9 أكثر منهم. تؤدي هذه المعاملة الإفراج عن الديدان من أجار.
- بعد 10 دقيقة، عندما يتم سحق الديدان حولها في المخزن المؤقت، نقلها إلى صفيحة نغ-أجار مع البكتيريا OP50 استخدام الماصة الشعرية سحبت. ضع اللوحة في 20 درجة مئوية ح 2-3 وحتى الديدان قد شُفي ويتحرك.
- بعد استردادها، فردياً نقل الديدان إلى لوحات نغ-أجار مع OP50 ونقل اللوحات حاضنة 25 درجة مئوية.
-
تسمح ف0-حقن الديدان تنمو وإرساء ذرية لمدة 3 أيام. شاشة ذرية1 و.
- في حالة استخدام كريسبر أول أكسيد الكربون أو التحويل المشاركين62،63،64،65، ثم حدد الديدان المرشح للفرز استناداً إلى عما إذا كان لديهم النمط الظاهري متحولة من الجينات مرجع. نقل هذه الديدان ملحوظ إلى لوحات نغ-أجار جديدة مع OP50 منفردة والسماح لهم بوضع و2 ذرية عند 20 درجة مئوية.
ملاحظة: النمط الظاهري المستخدمة للفرز كريسبر co أو التحديد ينبغي تقديم تقدير مبكر لنجاح تحرير Cas9. - في حالة عدم وجود النمط الظاهري كريسبر co، ميكروينجيكت بلازميد مراقبة إيجابية للمساعدة في تحسين كفاءة microinjection.
ملاحظة: على سبيل المثال، بما في ذلك بلازميد في مزيج الحقن الذي يشفر معلم مشري ميو-2 سوف يساعد تقييم كفاءة الحقن. سوف تكون الديدان حقن بنجاح مع pCFJ90 بعض ذرية مع فارينكسيس نيون.
- في حالة استخدام كريسبر أول أكسيد الكربون أو التحويل المشاركين62،63،64،65، ثم حدد الديدان المرشح للفرز استناداً إلى عما إذا كان لديهم النمط الظاهري متحولة من الجينات مرجع. نقل هذه الديدان ملحوظ إلى لوحات نغ-أجار جديدة مع OP50 منفردة والسماح لهم بوضع و2 ذرية عند 20 درجة مئوية.
- دراسة الديدان و1 لوجود عمليات التحرير المنشود. اختيار الأم1 و إلى بئر فردية من صفيحة 96-جيدا، ولها، وفحص الحمض النووي لها قبل التضخيم PCR الخاصة بإدراج أو تحليل تسلسل الحمض النووي، أو مساح نوكلاس المقايسة (CEL-1)66.
ملاحظة: يمكن إجراء هذه الاختبارات عند استخدام الفحص co-كريسبر/المشارك-conversion أو غيرها أو اختيار نظم65،66،،من6768. - للحصول على مزيد من المعلومات حول تقييم نتائج التحرير، انتقل إلى الخطوة 8.
6-إعداد هاواينسيس ص
- يوم واحد قبل microinjection، إثراء للأجنة في وقت مبكر بإنشاء خزان زوج في الليلة السابقة؛ الإناث المنفصلين حديثا سوف تحتوي على أجنة مخصبة حديثا. انظر ريم et al. 69 للحصول على التفاصيل.
- في يوم microinjection، جمع الأجنة باريل خلية واحدة (0-4 التسميد بعد ح) أنيسثيتيزينج الإناث جرابيد مع زيت القرنفل 0.02 في المائة في مياه البحر والقشط الأجنة خارج بلدها الحقيبة أمهات البطني باستخدام اللهب--سحبت برفق و ماصة زجاجية مدورة وزوج مملة من الملقط #3.
7- هاواينسيس P. الجنين Microinjection مع رنبس والرعاية ما بعد الحقن
- الردم أنبوب شعري سحبت مع حوالي 1 ميليلتر من مزيج حقن رنب الموصوفة أعلاه.
-
استخدام النتروجين المضغوط إلى ميكروينجيكت كل الجنين كما هو موضح في ريهم et al. 69.
- حقن الأجنة باريل تحت مجهر تشريح استخدام ميكروينجيكتور وميكرومانيبولاتور. تحميل 1.5 ميليلتر من مزيج الحقن في الجزء الخلفي من أنبوب شعري سحبت (4 بوصات-1.0 مم مع خيوط، سحبها باستخدام ميكروبيبيتي سحب الجهاز) باستخدام تلميح ماصة ميكرولوادير.
- إعداد الإبرة في جهاز الحقن، وكسر غيض الإبرة (كمية صغيرة جداً) باستخدام زوج من الملقط تحت نطاق تشريح. معايرة حجم ألقاها ضخ النفط الهالوكربون 700 وقياس قطر الفقاعة.
- قص 'الحوض الصغير' خارج الوكيل المعالجة باستخدام شفرة حلاقة. ملئه في منتصف الطريق مع مياه البحر تصفية تعقيم، وحتى خط الأجنة باريل في الحوض الصغير لتحقيق الاستقرار.
- حقن الأجنة باستخدام الإعداد microinjection، استقرار كل جنين مع زوج من الملقط أثناء الحقن. بعد الحقن، استخدم ماصة نقل زجاج نقل الأجنة أكثر إلى طبق ثقافة جديدة 60 ملم مليئة مياه البحر تصفية تعقيم في منتصف الطريق.
-
إذا كان قد حدث بالفعل الشعبة الأولى للنموذج جنينا الخلية 2 (4-6 ح بعد الإخصاب)، تولد الحيوانات تماما المسخ عن طريق حقن كلا بلاستوميريس. للتأكد من انشقاق مجموع المرحلة 2-خلية، شارك حقن في blastomeres ديكستران فيتك أو تريتك ونلاحظ أن الإشارة مقصور على بلاستوميري واحد تحت فلوري تشريح نطاق بعد الحقن.
- بدلاً من ذلك، إنشاء 'النصف-المسخ' الحيوانات بالحقن مجرد واحدة من بلاستوميريس اثنين في المرحلة 2-خلية (يسار يمين تقريبا مقسمة تبعاً للأنسجة والموقف على طول المحور A-P).
- حقن خلية واحدة في 8 خلايا جنين (7.5-9 ح بعد الإخصاب) لتقييد التحرير إلى طبقة جرثومة واحدة. انظر جربردينج et al. 70 لخارطة المبكر blastomere الأنساب.
-
تبني الأجنة في 60 ملم ثقافة الأطباق (لا يزيد عن 25 للطبق الواحد)، شغل في منتصف الطريق مع تصفية تعقيم مياه البحر، 'قبل اﻷوكسيجين' استخدام bubbler حوض أو عن طريق تهتز بقوة.
- ضع أطباق الأجنة في بلاستيكواري مختومة فضفاضة مبطنة بمناشف الورق الرطب للحفاظ على رطوبة ووضعها في حاضنة 26 درجة مئوية مع دورة ضوء الظلام ح 12.
- نقل الأجنة الباقين على قيد الحياة لتنظيف الأطباق مياه البحر كل بضعة أيام.
ملاحظة: قد يكون مثقف الأجنة في درجة حرارة الغرفة، على الرغم من أنها سوف تضع ببطء أكثر بكثير.
-
تشريح وإصلاح الأجنة في مراحل مختلفة لتحليل تعبير بها في الموقع التهجين أو جسم المصبوغة (انظر براون et al. 71 لدليل تجهيز، ومراجع إضافية ل التشريح وتثبيت72و في الموقع التهجين73وجسم تلطيخ74).
- جعل الإبر التشريح بخيوط قطعة سلك التنغستن بنت حوالي 0.5 في طولها إلى نهاية إبرة الأنسولين. شحذ الإبرة في هيدروكسيد الصوديوم تحت حالية. استخدم حقنه 1 مل كمؤشر إبرة تشريح.
- سد بئر واحدة من طبق زجاج 3-جيدا في منتصف الطريق مع حل الطازجة 9 أجزاء "بيم المخزن المؤقت" (0.1 M من أنابيب الأس الهيدروجيني 6.95، 2 مم عطا، 1 مم MgSO4)، وبرنامج تلفزيوني 1-الجزء العاشر 10 وجزء 1 32% منهاج عمل بيجين. ضع 3-5 أجنة في الطبق وكزه ثقب صغير في كل جنين، باستخدام إبرة حادة تنغستن كزة وواحدة المتبلدة قليلاً لتحقيق الاستقرار، السماح صفار البيض تتدفق ومثبت لتشغيل.
- برفق باستخدام زوج من التنغستن شحذ الإبر، ندف بعيداً الخارجي هما الأغشية المحيطة بالجنين باريل . تشريح لهم في مثبت جعل الأجنة أكثر قوة ولكن العمل بسرعة للحفاظ على الغشاء من أن تصبح ثابتة للجنين، مما يزيد من صعوبة إزالة الغشاء. السماح للأجنة لإصلاح لمدة 15-20 دقيقة لتلطيخ جسم أو 40-50 دقيقة التهجين في الموقع .
- صورة حية من hatchlings وتحليلها لتعمل الخصائص المورفولوجية والسلوكية أو إصلاح ووصمة عار لهم لإجراء تحليلات أكثر تفصيلاً. رفع hatchlings إلى مرحلة النضج الجنسي في 2-3 أشهر إنشاء خطوط محوره وراثيا (انظر كونتاراكيس وبافلوبولوس75 للرعاية فقس البيض وتفاصيل أخرى مفيدة) وخروج المغلوب.
8-تقييم نتائج التحرير
- إذا كان ذلك ممكناً، ابحث عن من النمط الظاهري البصرية أو الفنية في تحرير الخلايا أو الكائنات الحية.
ملاحظة: هذه العملية سوف تختلف على نطاق واسع بالتطبيق، ويرد بعض أمثلة في نهاية تلك الخطوات البروتوكول ذات الصلة المذكورة أعلاه. بعد تصحيح تحور الخلايا المنجلية في هسبكس، تحليل إنتاج الهيموغلوبين ب erythroblasts المتباينة باستخدام [هبلك] (الشكل 1أ). بالضربة قاضية لايل-2 مستقبلات الجينات في خلايا تي يمكن تأكيدها تلطيخ السطحية والتدفق الخلوي (الشكل 1ب). تقييم تعمل C. ايليجانس و هاواينسيس P. ، تراعي مورفولوجيا الحيوان والسلوك تحت مجهر الخفيفة أو الفلورسنت (الأرقام 1 ود 1). - لتحديد الكفاءة ونوع الجينوم عمليات التحرير التي تم إنشاؤها، في تجمعات خلايا تم تحريرها واستخراج عينات من الحمض النووي الجينوم باستخدام أدوات استخراج تجاري21.
-
لإجراء تقدير سريع لتشكيل إينديل، بكر تضخيم مالا يقل عن 200 زوج قاعدي حول القطع الموقع وتؤدي endonuclease1 T7 الاعتداء (T7E1)76 أو مساح (CEL-1 نوكلاس)77.
- إذا تشكيل إينديل في موقع Cas9-قص أو HDR ناجحة سوف إنشاء أو إزالة موقع قيود معروفة، النظر في استخدام هضم إنزيم التقييد لتقدير كفاءة التحرير6. المقايسة تعدد الأشكال (RFLP) طول يفتت قيود يمكن أن يكون وسيلة مريحة للتحقق من الكفاءة إذا كان يحدث أن تكون متاحة.
- الكمي دقيق لكفاءة التحرير وتحديد نتائج التحرير السائد، إرسال amplicon بكر سانجر قياسية التسلسل مع الإشعال على حد سواء إلى الأمام أو عكس.
ملاحظة: إذا كان تحليل لاستنساخ واحدة أو الكائن الحي، تحليل النتائج سانجر بسيط، كما هو موضح في الشكل 2أ. إذا كان تحليل مجموعة خلايا، ثم تحليل تشروماتوجرامس مع أداة على الإنترنت78، كما هو مبين في الشكل 2ب. - لإجراء تقييم كمي كامل وتسلسل تحرير النتائج، إجراء تسلسل عميق27،54، كما هو مبين في الشكل 2ج.
- تقييم مجموعة معينة من التغييرات خارج الهدف، بكر تضخيم مواقع خارج الهدف المتوقع وإرسالها لفئة الخدمات العامة الوطنية. لتمكين الكشف عن الكروموسومات المولدة، نفذ seq دليل79 أو الفائق، إزفاء على نطاق الجينوم تسلسل (هتجتس)80. للحصول على صورة كاملة عن عمليات التحرير خارج الهدف في عدد سكان الاستنساخ، إجراء تسلسل الجينوم الجامع (WGS)81،،من8283.
ملاحظة: هناك مجموعة متنوعة من أساليب لقياس الهدف ومن إيقاف تحريرات الجينوم، أوضح كذلك في استعراض مختلف المواد84،،من8586.
Representative Results
هذه التجارب يمكن استخدامها إظهار كيف قبل تجميع Cas9 رنب للتلاعب بمورثات الخلايا الأولية والكائنات كلها. الباحثون تنقية أو شراء بروتين Cas9 وسجرنا، الجمع بين العنصرين قبل تشكيل المجمع، ويعرض رنب في الخلايا أو الكائن الحي للفائدة. بعد السماح بوقت كاف لتحرير تحدث وتحقق لأبناء الجيل القادم أن يولد (إذا كان قابلاً للتطبيق)، لتعمل و/أو جمع الخلايا للتنميط. ويمكن ملاحظة تعمل عن طريق الاختبارات الوظيفية، وفحوصات التعبير، والتصور (بالعين المجردة أو بالمجهر)، أو أساليب أخرى، اعتماداً على التجربة.
على سبيل المثال، يمكن متباينة في الكريات الحمراء هسبكس التي تم تحريرها لتصحيح الطفرة بيتا جلوبين الذي يتسبب في مرض الخلية المنجلية وجزيئي لإنتاج صحية أو فقر الدم المنجلي الهيموغلوبين27،87 (الرقم 1 A). تحرير الخلايا T لضرب عالية تقارب إيل-2 مستقبلات المورثة، CD25 (IL2RA)، يمكن تحليلها بواسطة تلطيخ السطحية والتدفق الخلوي88، وحلل وظيفيا للكشف عن استجابة إرسال إشارات لتحفيز إيل-2 (الشكل 1ب ). يمكن أيضا إعادة برمجة خلايا تي في العديد من الطرق الهامة سريرياً التي تتطلب تقييما لتعمل مختلفة، بما في ذلك الفعالية من الإصابة بفيروس نقص المناعة البشرية89 و في فيفو انتيتومور كفاءة السيارة تي الخلايا11.
استخدام نهج فحص co-كريسبر/المشارك-conversion، الديدان C. ايليجانس يتم تحريرها في وقت واحد في اثنين المكاني62. تقرير التنمية البشرية في الجينات مرجع دبي-10 استخدام سودن إصلاح قالب النتائج طفرة مكسب لدالة مهيمنة بسهولة-وسجل دبي-10 . الحيواناتdpy-10(gof) 1و متخالف رولر (رول) والحيوانات متماثل dpy-10(gof) بدين وقصير (دبي). وجود النمط الظاهري يشير إلى أن Cas9 التحرير وقع في هذه الحيوانات ويحسن احتمالات تحديد حدث تحرير في محور ثان في الحيوانات1 رول أو دبي و. ينبغي أن تؤدي تجربة ناجحة لتحرير 33-50 في المائة من حقن الديدان0 ف الغلة ذرية1 و 20 أو أكثر التي هي رول أو دبي90. ثم أن من الممكن اختيار الحيوانات غير رول للعودة دبي-10 إلى wildtype وحدد لتحرير متماثل للفائدة. وكقاعدة عامة، ينبغي أن يكون تركيز كرنا استهداف الجين مرجع كريسبر co نصف من كرنا استهداف الجين للفائدة. إذا لم يتم استرداد عملية تحرير في الجينات للفائدة، يمكن تعديل نسب "الكشف كريسبر" اثنين لزيادة احتمال استعادة الطفرة المنشودة. على سبيل المثال، زيادة مقدار كرنا لجينات الفائدة بالنسبة إلى كرنا الجينات مرجع ستزيد النسبة المئوية للديدان امتلاك عمليات التحرير في جينات الاهتمام داخل سكان الديدان التي تمتلك أيضا تعديلات في موضع الجينات مرجع. تختلف الترددات التحويل المشاركين، ولكن عادة ما تكون المعدلات 20-60%، وكثيراً ما تسفر عن تحريرات متماثل في الجيل1 و (الشكل 1ج).
ف. hatchlings هاواينسيس التي تم تحريرها لضرب الجين البطن-ب (عبد-ب) عرض تشوهات واضحة المورفولوجية3 (الشكل 1د). هذا الجين المطلوب للزخرفة البطن الصحيحة، وتقديم نتائجها اختلال في الصدر من نوع القفز والمشي الساقين استبدال الساقين السباحة والربط التي عادة ما تكون في البطن.
تحديد الجينوم تحرير النتائج على مستوى قد يتطلب التسلسل أو في المختبر والرزن يقوم بالكشف عن التغييرات في التسلسل. هنا، نعرض الممثل تسلسل البيانات من نموذج أنواع الخلايا والكائنات الحية، تسليط الضوء على نهج مختلفة لتحرير التحديد الكمي. لاحظ أن تسميات الشكل هي عامة لجميع الأساليب الموضحة هنا يمكن تطبيقها على أي نظام بيولوجي.
النهج القائم على التسلسل تتفاوت في التعقيد التقني وعمق النتائج. السكان المحررة الاستنساخ أو الكائنات الفردية يمكن فصله بسهولة، يمكن متسلسلة الأفراد تم تحريرها بعد استخراج الحمض النووي الجينوم. وتكشف النتائج التسلسل سانجر القياسية سوف تغيير تسلسل في موقع Cas9-قطع في فرد، مع فراميشيفتس الافتراضية التي سوف تخل بوظيفتها (الشكل 2أ). على شبكة الإنترنت أداة تستخدم لتسلسل نهج آخر هو سانجر على أساس التسلسل التي يمكن تطبيقها على السكان مختلطة بدلاً من طفرات الفردية78. ويتم تحليل تسلسل مع أداة على إنترنت يمكن التقريبي العام التحرير الكفاءة، فضلا عن نتائج التسلسل الغالبة. وترد البيانات الممثلة في الشكل 2ب.
أسلوب تسلسل الأكثر شمولاً الموصوفة هنا تسلسل العميق (يشار إليها في بعض الأحيان التسلسل الفائق أو الجيل القادم). يوفر هذا الأسلوب تسلسل الحمض النووي من مورثات فردية في مجموعة سكانية مختلطة. ويمكن توضيح هذه البيانات في مجموعة متنوعة من الطرق. هنا، وقد صنفنا ما يلي تسلسل الفردية من تحرير الخلايا استناداً إلى نتائج التحرير (الشكل 2-ج). يتم تحرير معظم الخلايا عن طريق المسار نهيج، الذي عادة ما يؤدي إلى تعطيل الجينات. وفي حالات أخرى، تم تبديل الجين المستهدف بها لصيغة بديلة عبر27من تقرير التنمية البشرية.
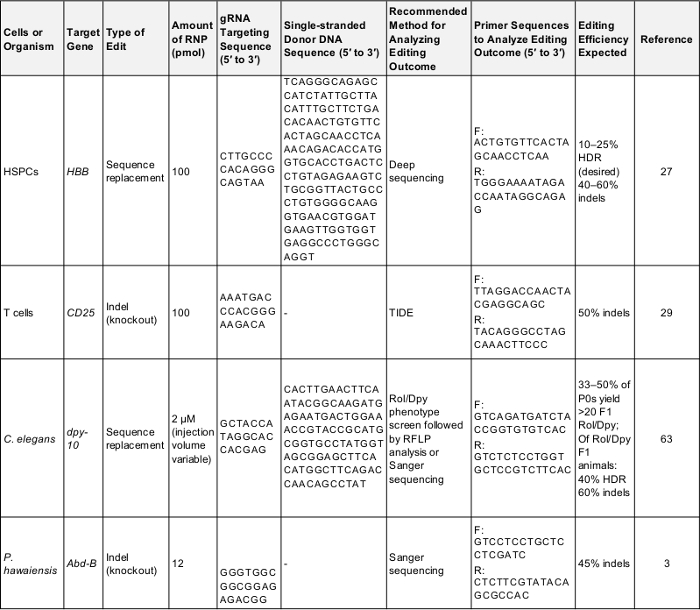
الجدول 1: ضوابط إيجابية للجينوم أولى تجارب التحرير- يظهر هذا الجدول المعلومات الأساسية اللازمة لأداء جينوم أول مرة تحرير التجربة في كل من الخلايا والكائنات الموصوفة في هذا البروتوكول. عقب هذه المعلمات من المرجح أن تؤدي إلى نتيجة ناجحة يمكن استخدامه لاختبار البروتوكول أو كخط أساس للمقارنة مرة المجرب هو استهداف جين لمصلحتهم الخاصة. واو: إلى الأمام، عكس r:، تقرير التنمية البشرية: إصلاح الموجه من التماثل. اضغط هنا لتحميل هذا الجدول.

الشكل 1 : الممثل المظهرية ناتج عن رنب Cas9 تحرير خلايا الإنسان الأولية والكائنات الحية. (أ) هذا هو تتبع [هبلك] تبين أن بعد الجينوم نجاح التحرير، هسبكس أن تكون متباينة في أواخر المرحلة اريثروبلاستس سوف تنتج الهيموغلوبين أكثر وظيفية من المنجل الهيموغلوبين. إنتاج الكريات الحمراء متحولة الهيموغلوبين فقر الدم المنجلي (HbS)، بينما الخلايا المحررة بنجاح سوف تنتج الهيموغلوبين صحية (HbA و HbA2) فضلا عن الهيموغلوبين الجنيني (HbF). هي رسوم بيانية امتصاص في وحدات التعسفي (الاتحاد الأفريقي). نشرت هذا الفريق لأول مرة في دويت هt al. 27-هو طبع بإذن من "الرابطة الأمريكية" "النهوض بالعلم". (ب) على اليسار، لكل حالة، وهذا الفريق يظهر التدفق الخلوي بيانات توضح أن الملطخة بسطح خلايا تي لا تعبر عن CD25 بعد الجين CD25 قد خرج مع رنب. وفرة CD25 المرسومة على المحور السيني مع حجم الخلية على المحور ص. على اليمين، لكل حالة، ويظهر هذا الفريق الكمي والرمات-Stat5 (pStat5) بعد الاستقراء مع إيل-2. مما يشير إلى ينخفض عند مستقبلات إيل-2 غياب (CD25 كو). وفرة pStat5 المرسومة على المحور س، ويتم مقارنة البيانات الناجمة عن ثلاثة مستويات مختلفة من المدخلات إيل-2 عمودياً. (ج) يظهر هذا الفريق ايليجانس كاينورهابديتيس co-كريسبر/مشارك-conversion شاشة استهداف دبي-10 كعلامة التحويل المشاركين. اثنين دليل الكشف المستهدف اثنين المكاني و دبي-10 والجينات المفضلة الخاصة بك (يفج)، في نفس ف0-حقن الحيوان. نتائج تقرير التنمية البشرية في دبي-10 النمط الظاهري رول أو دبي. اختيار الحيوانات1 رول أو دبي و يزيد من فرص تحديد عمليات التحرير في موضع الثاني. (د) تبين هذا الفريق أن wildtype هاواينسيس باريل hatchlings البطن العادي مع السباحة وربط الساقين. Hatchlings المغلوب عبد-B (و0 الأفراد) وضع البطن تحول نحو الصدر. وهكذا، السباحة والساقين المراسي قد ولت والاستعاضة عن الساقين القفز والمشي المقترنة الصدر عادية. الرجاء انقر هنا لمشاهدة نسخة أكبر من هذا الرقم-

الشكل 2 : نتائج نموذجية من تحرير أساليب تحليل نتائج. (أ) هذا الفريق يظهر أمثلة سانجر تسلسل النتائج الفردية و1 هاواينسيس P. الكائنات الحية، بما في ذلك تسلسل wildtype وثلاثة إينديلس المختلفة التي تعطل وظيفة الجينات بتحويل الإطار القراءة المفتوحة. (ب) "المد هذه" النتائج تظهر المجموعة من عمليات الإدراج والحذف الأحداث التي وقعت في موقع هدف Cas9 في بركة خلايا تي متسلسلة. المحور يشير إلى طول معين الإدراج أو الحذف في النيوكليوتيدات. (ج) تبين هذه النتائج تسلسل عميق أي الجينوم التحرير دون نوكليوفيكشن أو جرنة، ونجاح تحرير مع سليمة Cas9 رنب، مجمعة حسب نتائج إصلاح الحمض النووي في هسبكس. الرجاء انقر هنا لمشاهدة نسخة أكبر من هذا الرقم-
Discussion
وضع جينوم قوية تحرير البروتوكول في خلية السطر أو الكائن الحي لمصلحة يتطلب التحسين والتجريبية اختبار العديد من المعلمات الرئيسية، تمت مناقشتها في هذا القسم. محاولة عدد قليل من الاختلافات من النهج العام المقدمة هنا تشجيعا كبيرا. القيد الرئيسي من هذا البروتوكول أن تطبيق هذه الأساليب على الخلايا الأخرى أو الكائنات قد تؤدي إلى نتائج مختلفة تبعاً للأنواع التي شملتها الدراسة، وقد لا تشجع تصميم تجريبي الذي يؤدي إلى دفع جينات ذات الكفاءة العالية الإدراج الحمض النووي. وبالتالي، نوصي بدءاً من الأساليب المعروضة هنا، واستكشاف الأخطاء وإصلاحها كما هو موضح أدناه.
استكشاف الأخطاء وإصلاحها الجينوم تحرير جودة الكاشف:
توليد أو شراء الكواشف عالية الجودة خطوة حاسمة في الجينوم أي تحرير البروتوكول. يمكن تنقية في المختبر البروتين Cas9 أو شراؤها تجارياً. ملاحظة العديد من البروتوكولات تركيز نهائي ل Cas9 في وصفات رنب، لكن الجينات الأمثل تحرير النشاط سيتوقف على النشاط المحدد لأي إعداد البروتين Cas9 الفردية، التي تختلف تبعاً للمصدر. مرة واحدة وتعمل على البروتوكول المعروضة هنا، النظر في تحسين كمية رنب المستخدمة من قبل المعايرة المستويات Cas9 لإنشاء بتركيز أمثل: واحد يوفر الانقسام مستهدفة محددة للغاية والحمض النووي دون الانقسام خارج الهدف لا لزوم لها والناجمة عن Cas9 المفرطة40.
يمكن أيضا أن تكون دليل الحمض النووي الريبي النقاء والتجانس المحددات للجينوم التحرير النجاح22. سجرناس التي تم شراؤها أو مكونات منفصلة كرنا وتراكرنا الكواشف عامة عالية الجودة ومجموعة متنوعة من التعديلات الكيميائية متاحة لمكافحة مشاكل مع تدهور الجيش الملكي النيبالي أو لإشراب ميزات إضافية إلى رنب91. بينما جرناس معدلة كيميائيا قد لا يكون ضروريا للجينوم القياسية تحرير التجارب، لاحظ بعض المجموعات أعلى بكثير تحرير الكفاءة مع هذه الكواشف، ولذلك قد يكون من المفيد محاولة بعد إتقان العملية و/أو عند تدهور جرنة يبدو أن22،مسألة91. النسخ في المختبر وجل اللاحقة هو تنقية بديل غير مكلفة، التي قد تكون كافية للجينوم الروتينية تحرير تجارب17،21،،من4950. علاوة على ذلك، عدة نهج التي تطبق عادة على إنتاج جرنة متجانسة السكان في الأحياء، بما في ذلك الختان المستندة إلى رايبوزيم والحمض الريبي النووي النقال من أدلة فردية، ويمكن أن تمتد إلى في المختبر إعداد الجيش الملكي النيبالي لتوليد الأنظف 92من المنتجات.
دليل الحمض النووي الريبي والمانحة الحمض النووي تصميم نصائح:
دليل الحمض النووي الريبي التحديد عامل حاسم في تحقيق كفاءة عالية في الهدف التحرير مع التقليل من احتمالات الانقسام خارج الهدف. للمساعدة في تحديد دليل، استخدمت عدة دراسات شاشات الفائق مقترنة بتسلسل الجيل التالي لتجميع ميزات تسلسل من دلائل نجاح47،79،،من9394، ،من 9596. هذه الميزات قد استخدمت لوضع خوارزميات التنبؤية والأدوات على الإنترنت للمساعدة في دليل التحديد44،45،46،،من4748. ترتكز هذه الخوارزميات على شاشات استخدام النظم القائمة على الحمض النووي لدليل التعبير الجيش الملكي النيبالي. يتم التعبير عن أدلة استخدام مروج "الثالث بول"، والتعبير عنها ولذلك فعرضه للقيود المرتبطة بالنسخ "بول الثالث"، مثل الإنهاء السابق لأوانه عندما تواجه المسارات من اليوراسيل97،98، 99. ومع ذلك، باستخدام رنبس مع المختبر-دليل تجميعي الكشف يتجاوز تلك الشواغل ويبسط القيود على تصميم الدليل. سمات مشتركة التي انبثقت من هذه الخوارزميات وأكدت العديد من الدراسات مع تحرير الجينوم فعالة للغاية، هو وجود البيورين، لا سيما جوانين، في نهاية محددة الهدف تسلسل الدليل 3 '. هذه الميزة دليل نجاحا كبيرا بين الكائنات الحية تتراوح بين الثدييات C. ايليجانسوذباب الفاكهة الزرد65،،من100101. وبالإضافة إلى ذلك، C. ايليجانس، تصميم أدلة مع دينوكليوتيدي زز في نهاية 3 ' استهداف المنطقة دليل استراتيجية فعالة للتنبؤ بفعالية عالية دليل الكشف65. ومن الناحية المثالية، اختبار أدلة متعددة في نفس الوقت لتحديد ما هو الأكثر نجاحا لتطبيق محدد.
عند محاولة إدخال تسلسل الحمض النووي في الجينوم، تصميم قالب الحمض النووي أو المانحة مهم أيضا. يتم إدراج اليغنوكليوتيد الذين تقطعت بهم السبل-واحد المانحين (سودنس) أكثر موثوقية من قوالب إصلاح نموذجية أخرى، خطي مزدوج-الذين تقطعت بهم السبل وبلازميد الحمض النووي54،،من55102. في بعض المكاني، ويمكن تحسين كفاءة تقرير التنمية البشرية مع سودنس التي تعتبر مكملة لغير المستهدفة أو المشردين حبلا الحمض النووي وامتلاك الأسلحة التماثل التي غير متماثلة في الطول27،55. حيث يتم إدراج في موقع قطع القالب إصلاح ويشمل تسلسل المستهدفة، يجب اتخاذ خطوات لمنع Cas9 من ناهضة الجهة المانحة الحمض النووي قبل أو بعد الإدراج الجينوم. ويتم ذلك بجعل الطفرات الصامتة إلى بام تسلسل أو بذور المنطقة، تجنب الاعتراف بواسطة Cas9 مع الاحتفاظ بوظيفة ال21،الجينات مدرج103. على الرغم من النوكليوتيدات واحدة حتى يتغير إلى بام من المحتمل أن إلغاء ربط104، في محاولة لتغيير النيوكليوتيدات أربعة على الأقل أن تكون آمنة.
الأهمية والتطبيقات المستقبلية:
جينوم التحرير مع Cas9 كريسبر برز كوسيلة قوية تمكن السطحية التحوير الوراثي لأي كائن حي. تحرير مع رنب Cas9 يأخذ جهد أكثر قليلاً في البداية ولكن واضحة لاستخدامها بمجرد إنشاء الكواشف والبروتوكولات في مختبر. تحرير الخلايا مع رنب قبل تجميع بدلاً من بلازميد الحمض النووي يؤدي إلى ارتفاع الكفاءة التحرير الشامل، بما في ذلك الإدراج الجينات صعوبة تحقيق عن طريق تقرير التنمية البشرية، مع أقل آثار خارج الهدف24،،من2526 , 27 , 29-علاوة على ذلك، المجربون تجنب المشاكل مع التعبير الجيني والحمض النووي الريبي تدهور، طي البروتين، والرابطة بين جرنة و Cas9 الجزيئات توليفها بشكل منفصل داخل الخلية،من2223. تحرير رنب تلتف أيضا شواغل السلامة عن الطفرات insertional والتعبير المطرد التي قد تنشأ عندما تكون طرق التسليم الفيروسية المستخدمة سريرياً14. وبسبب هذه المزايا، العديد من العلماء إجراء ما قبل السريرية، صالح تجارب إثبات مفهوم تحرير رنب للتطبيقات العلاجية البشرية. في فيفو و السابقين فيفو الجينوم رنب-على أساس نهج التحرير في التنمية لعلاج أو حتى علاج مجموعة متنوعة من الظروف، من الأمراض الوراثية مثل دوشين ضمور العضلات105 ومرض الخلية المنجلية27 إلى فيروس نقص المناعة البشرية29 والسرطان11. من المثير للاهتمام، Cas9 رنب متزايدة يعمل كوسيلة إيصال للهندسة الزراعية، لأنها تمكن 'خالية من الحمض النووي' تحرير محطات33،،من3436.
Disclosures
أصحاب مارسون ألكسندر والذرة هاء يعقوب هم المؤسسين "المداواة أضواء كاشفة". يعقوب هاء الذرة مستشار "البعثة المداواة" ومختبرة تلقت دعم البحوث التي ترعاها من استرا زينيكا، وشركة فايزر. ألكسندر مارسون مستشار جونو التداوي والمداواة الميثاق، ومختبرة تلقت دعم البحوث التي ترعاها من المداواة جونو، ابينوميكس، وشركة سانوفي. كما طبق مختبرة لبراءات الاختراع ذات الصلة بالتكنولوجيا رنب Cas9.
Acknowledgments
ونشكر العديد من الأعضاء السابقة مختبراتنا و "منطقة خليج" الجينوم المجتمع التحرير لمساهمتها في تطوير هذه الأساليب. ونحن نشكر روس ويلسون للغاية قراءة هذا المخطوط.
البحث ألكسندر مارسون معتمد من قبل هدية من أرنب جيك ومنح "جمعية التصلب المتعدد وطنية" (CA 1074-A-21). حاصل على "جائزة الوظيفي" "علماء الطب" من صندوق ويلكوم بوروز مارسون ألكسندر وهو "محقق بيوهوب زوكربيرج تشان". البحوث يعقوب هاء الذرة يدعمه لي كا شينغ المؤسسة والمعهد الطبي التراث البحوث الطبية، ومعهد كاليفورنيا "الطب التجديدي". وتمول البحوث فاربود بنوم وباربارا ج. ماير في الجزء NIGMS المنحة R01 GM030702 إلى باربرا ج. ماير، وهو محقق من معهد هوارد هيوز الطبي. البحوث جارفيس إيرين ونيبام H. باتل تموله جزئيا المنحة NSF دائرة الرقابة الداخلية--1257379 وتعترف جارفيس إيرين الدعم من جرفب جبهة الخلاص الوطني وزمالة الدراسات العليا فيلوماثيا.
Materials
| Name | Company | Catalog Number | Comments |
| Reagents/Materials | |||
| DNA oligonucleotides | Integrated DNA Technologies | - | IDT will provide custom DNA sequences, including those in Table 1 |
| Guide RNAs | Synthego | - | Synthego will provide high-quality sgRNAs for S. pyogenes Cas9, including custom sgRNAs containing the targeting sequences included in Table 1 |
| Purified Cas9 protein (EnGen Cas9 NLS, S. pyogenes) | New England Biosciences | M0646T | If possible, purifying Cas9 in-house or purchasing from local core facilities is a less expensive option |
| Normal peripheral blood CD34+ stem/progenitor cells | AllCells | PB032-2 | |
| StemSpan SFEM | StemCell Technologies | 09650 | |
| StemSpan CC110 | StemCell Technologies | 02697 | |
| P3 Primary Cell 4D-Nucleofector X Kit | Lonza | V4XP-3032 | |
| RPMI-1640 Medium, With sodium bicarbonate, without L-glutamine, liquid | Sigma | R0883-6X500ML | |
| EasySep™ Human T Cell Isolation Kit | Stemcell | 17951 | |
| cell culture plate, 96 wells, round | Fisher Scientific | 3799 | |
| CTS (Cell Therapy Systems) Dynabeads CD3/CD28 | Life Tech | 40203D | |
| Reombinant Human IL-2 | UCSF Pharmacy | NA | |
| SepMate-50 500-pack IVD | Stemcell Technologies | 85460 | |
| OP50 Escherichia coli | Caenorhabditis Genetics Center | OP-50 | https://cgc.umn.edu/ |
| Nematode Growth Media agar in petri dishes | - | - | See Stiernagle, T (ref. 59) |
| Standard borosilicate glass capillaries with filament: 4 in (100 mm), 1/0.58 OD/ID | World Precision Instruments | 1B100F-4 | |
| Single-barrel standard borosilicate glass capillaries: 6 in (152 mm), 2/1.12 OD/ID | World Precision Instruments | 1B200-6 | |
| Cover glass; 24 × 50 mm | Thermo Fisher Scientific | 12-544E | |
| Cover glass; 22 × 22 mm | Thermo Fisher Scientific | 12-518-105K | |
| Apex LE agarose | Genesee Scientific | 20-102 | |
| Halocarbon oil 700 | Sigma-Aldrich | H8898-100ML | |
| pCFJ90 plasmid | Addgene | 19327 | |
| Compressed nitrogen | - | ||
| 60 mM culture dishes | BD | ||
| Capillary tubes with filament: 4 in (1.0 mm) | World Precision Instruments | T2100F-4 | |
| Sylgard 184 | Dow Corning | ||
| Petri dishes (100 × 15 mm) | - | ||
| Tungsten wire (0.005 in. diameter) | Ted Pella | ||
| Perfluoroalkoxy alkane (PFA) | - | ||
| Marine salt | - | ||
| 9" pasteur pipettes | - | ||
| Phenol red | - | ||
| Nuclease-free water | - | ||
| Equipment | |||
| 4D Nucleofector | Lonza | AAF-1002X | |
| MZ75 Stereomicroscope | Leica | Out-of-production. Current model is the M80 Stereomicroscope | |
| Axio Vert35 inverted phase contrast fluorescent microscope | Zeiss | Out-of-production. Current model is the Axio VertA.1 | |
| Laser-based micropipette puller (for C. elegans protocol) | Sutter Instrument | FG-P2000 | |
| Picoliter Microinjector (for C. elegans protocol) | Warner Instruments | PLI-100A | |
| Three-axis Joystick oil hydraulic micromanipulator | Narishige International | MO-202U | |
| Coarse manipulator | Narishige International | MMN-1 | |
| Micropipette puller (for P. hawaiensis protocol) | Sutter Instrument | P-80/PC | |
| Microinjector (for P. hawaiensis protocol) | Narishige | IM300 | |
| Microloader pipette tips | Eppendorf | 5242956003 | |
| NG-agar |
References
- Komor, A. C., Badran, A. H., Liu, D. R. CRISPR-based technologies for the manipulation of eukaryotic genomes. Cell. , 1-17 (2016).
- Barrangou, R., Horvath, P. A decade of discovery: CRISPR functions and applications. Nature Microbiology. 2, 1-9 (2017).
- Martin, A., Serano, J. M., et al. CRISPR/Cas9 mutagenesis reveals versatile roles of Hox genes in crustacean limb specification and evolution. Current Biology. 26 (1), 14-26 (2016).
- Goldstein, B., King, N. The future of cell biology: emerging model organisms. Trends in Cell Biology. 26 (11), 818-824 (2016).
- Mali, P., Yang, L., et al. RNA-guided human genome engineering via Cas9. Science. 339 (6121), 823-826 (2013).
- Cong, L., Ran, F. A., et al. Multiplex genome engineering using CRISPR/Cas systems. Science. 339 (6121), 819-823 (2013).
- Deltcheva, E., Chylinski, K., et al. CRISPR RNA maturation by trans-encoded small RNA and host factor RNase III. Nature. 471 (7340), 602-607 (2011).
- Jinek, M., Chylinski, K., Fonfara, I., Hauer, M., Doudna, J. A., Charpentier, E. A Programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science. 337 (6096), 816-821 (2012).
- Nowak, C. M., Lawson, S., Zerez, M., Bleris, L. Guide RNA engineering for versatile Cas9 functionality. Nucleic Acids Research. 44 (20), 9555-9564 (2016).
- Jiang, W., Cox, D., Zhang, F., Bikard, D., Marraffini, L. A. RNA-guided editing of bacterial genomes using CRISPR-Cas systems. Nature Biotechnology. , 1-9 (2013).
- Rupp, L. J., Schumann, K., et al. CRISPR/Cas9-mediated PD-1 disruption enhances anti-tumor efficacy of human chimeric antigen receptor T cells. Scientific Reports. 7 (1), 737 (2017).
- Graham, D. B., Root, D. E. Resources for the design of CRISPR gene editing experiments. Genome Biology. 16, 260 (2015).
- Wang, H., La Russa, M., Qi, L. S. CRISPR/Cas9 in genome editing and beyond. Annual Review of Biochemistry. 85, 2270 (2016).
- Nelson, C. E., Gersbach, C. A. Engineering delivery vehicles for genome editing. Annual Review of Chemical and Biomolecular Engineering. 7, 637-662 (2016).
- Yin, H., Kauffman, K. J., Anderson, D. G. Delivery technologies for genome editing. Nature Reviews Drug Discovery. 16 (6), 387-399 (2017).
- Zuris, J. A., Thompson, D. B., et al. Cationic lipid-mediated delivery of proteins enables efficient protein-based genome editing in vitro and in vivo. Nature Biotechnology. 33 (1), 73-80 (2015).
- Cho, S. W., Lee, J., Carroll, D., Kim, J. -S., Lee, J. Heritable gene knockout in Caenorhabditis elegans by direct injection of Cas9-sgRNA ribonucleoproteins. Genetics. 195 (3), 1177-1180 (2013).
- Wang, W., Kutny, P. M., et al. Delivery of Cas9 protein into mouse zygotes through a series of electroporation dramatically increases the efficiency of model creation. Journal of Genetics and Genomics. 43 (5), 319-327 (2016).
- Chen, S., Lee, B., Lee, A. Y. -F., Modzelewski, A. J., He, L. Highly efficient mouse genome editing by CRISPR ribonucleoprotein electroporation of zygotes. Journal of Biological Chemistry. 291 (28), 14457-14467 (2016).
- Remy, S., Chenouard, V., et al. Generation of gene-edited rats by delivery of CRISPR/Cas9 protein and donor DNA into intact zygotes using electroporation. Scientific Reports. 7 (1), 16554 (2017).
- DeWitt, M. A., Corn, J. E., Carroll, D. Genome editing via delivery of Cas9 ribonucleoprotein. Methods. , 1-7 (2017).
- Hendel, A., Bak, R. O., et al. Chemically modified guide RNAs enhance CRISPR-Cas genome editing in human primary cells. Nature Biotechnology. 33 (9), 985-989 (2015).
- Thyme, S. B., Akhmetova, L., Montague, T. G., Valen, E., Schier, A. F. Internal guide RNA interactions interfere with Cas9-mediated cleavage. Nature Communications. 7, 11750 (2016).
- Kim, S., Kim, D., Cho, S. W., Kim, J., Kim, J. -S. Highly efficient RNA-guided genome editing in human cells via delivery of purified Cas9 ribonucleoproteins. Genome Research. 24 (6), 1012-1019 (2014).
- Lin, S., Staahl, B. T., Alla, R. K., Doudna, J. A. Enhanced homology-directed human genome engineering by controlled timing of CRISPR/Cas9 delivery. eLife. 3, 04766 (2014).
- Liang, X., Potter, J., et al. Rapid and highly efficient mammalian cell engineering via Cas9 protein transfection. Journal of Biotechnology. 208, 44-53 (2015).
- DeWitt, M. A., Magis, W., Bray, N. L., Wang, T. Selection-free genome editing of the sickle mutation in human adult hematopoietic stem/progenitor cells. Science Translational Medicine. 8 (360), (2016).
- Ramakrishna, S., Kwaku Dad, A. -B., Beloor, J., Gopalappa, R., Lee, S. -K., Kim, H. Gene disruption by cell-penetrating peptide-mediated delivery of Cas9 protein and guide RNA. Genome Research. 24 (6), 1020-1027 (2014).
- Schumann, K., Lin, S., et al. Generation of knock-in primary human T cells using Cas9 ribonucleoproteins. Proceedings of the National Academy of Sciences of the United States of America. 112 (33), 10437-10442 (2015).
- Lee, J. -S., Kwak, S. -J., et al. RNA-guided genome editing in Drosophila with the purified Cas9 protein. G3: Genes, Genomes, Genetics (Bethesda, MD). 4 (7), 1291-1295 (2014).
- Sung, Y. H., Kim, J. M., et al. Highly efficient gene knockout in mice and zebrafish with RNA-guided endonucleases. Genome Research. 24 (1), 125-131 (2014).
- Menoret, S., De Cian, A., et al. Homology-directed repair in rodent zygotes using Cas9 and TALEN engineered proteins. Scientific Reports. 5, 14410 (2015).
- Woo, J. W., Kim, J., et al. DNA-free genome editing in plants with preassembled CRISPR-Cas9 ribonucleoproteins. Nature Biotechnology. 33 (11), 1162-1164 (2015).
- Malnoy, M., Viola, R., et al. DNA-free genetically edited grapevine and apple protoplast using CRISPR/Cas9 ribonucleoproteins. Frontiers in Plant Science. 7, 1904 (2016).
- Svitashev, S., Schwartz, C., Lenderts, B., Young, J. K., Mark Cigan, A. Genome editing in maize directed by CRISPR-Cas9 ribonucleoprotein complexes. Nature Communications. 7, 13274 (2016).
- Liang, Z., Chen, K., et al. Efficient DNA-free genome editing of bread wheat using CRISPR/Cas9 ribonucleoprotein complexes. Nature Communications. 8, 14261 (2017).
- Shin, S. -E., Lim, J. -M., et al. CRISPR/Cas9-induced knockout and knock-in mutations in Chlamydomonas reinhardtii. Scientific Reports. 6, 27810 (2016).
- Pohl, C., Kiel, J. A. K. W., Driessen, A. J. M., Bovenberg, R. A. L., Nygård, Y. CRISPR/Cas9 based genome editing of Penicillium chrysogenum. ACS Synthetic Biology. 5 (7), 754-764 (2016).
- Grahl, N., Demers, E. G., Crocker, A. W., Hogan, D. A. Use of RNA-protein complexes for genome editing in non-albicans Candida species. mSphere. 2 (3), (2017).
- Rivera-Torres, N., Kmiec, E. B. A standard methodology to examine on-site mutagenicity as a function of point mutation repair catalyzed by CRISPR/Cas9 and ssODN in human cells. Journal of Visualized Experiments. (126), (2017).
- Nandal, A., Mallon, B., Telugu, B. P. Efficient generation and editing of feeder-free IPSCs from human pancreatic cells using the CRISPR-Cas9 system. Journal of Visualized Experiments. (129), (2017).
- Mohr, S. E., Hu, Y., Ewen-Campen, B., Housden, B. E., Viswanatha, R., Perrimon, N. CRISPR guide RNA design for research applications. The FEBS Journal. 283 (17), 3232-3238 (2016).
- Bauer, D. E., Canver, M. C., Orkin, S. H. Generation of genomic deletions in mammalian cell lines via CRISPR/Cas9. Journal of Visualized Experiments. (95), e52118 (2015).
- Hsu, P. D., Scott, D. A., et al. DNA targeting specificity of RNA-guided Cas9 nucleases. Nature Biotechnology. 31 (9), 827-832 (2013).
- Heigwer, F., Kerr, G., Boutros, M. E-CRISP: fast CRISPR target site identification. Nature Methods. 11 (2), 122-123 (2014).
- Moreno-Mateos, M. A., Vejnar, C. E., et al. CRISPRscan: designing highly efficient sgRNAs for CRISPR-Cas9 targeting in vivo. Nature Methods. 12 (10), 982-988 (2015).
- Labun, K., Montague, T. G., Gagnon, J. A., Thyme, S. B., Valen, E. CHOPCHOP v2: a web tool for the next generation of CRISPR genome engineering. Nucleic Acids Research. 44, 272-276 (2016).
- Haeussler, M., Schönig, K., et al. Evaluation of off-target and on-target scoring algorithms and integration into the guide RNA selection tool CRISPOR. Genome Biology. 17 (1), 148 (2016).
- Lo, T. -W., Pickle, C. S., et al. Precise and heritable genome editing in evolutionarily diverse nematodes using TALENs and CRISPR/Cas9 to engineer insertions and deletions. Genetics. 195 (2), 331-348 (2013).
- Bassett, A., Liu, J. -L. CRISPR/Cas9 mediated genome engineering in Drosophila. Methods. 69 (2), 128-136 (2014).
- Prior, H., Jawad, A. K., MacConnachie, L., Beg, A. A. Highly efficient, rapid and co-CRISPR independent genome editing in Caenorhabditis elegans. G3: Genes, Genomes, Genetics. , Bethesda, MD. (2017).
- Hirsh, A. Cas9 expression and purification protocol. protocols.io. , (2017).
- DeWitt, M. A., Wong, J. In vitro transcription of guide RNAs. protocols.io. , (2017).
- Yang, L., Guell, M., et al. Optimization of scarless human stem cell genome editing. Nucleic Acids Research. 41 (19), 9049-9061 (2013).
- Richardson, C. D., Ray, G. J., DeWitt, M. A., Curie, G. L., Corn, J. E. Enhancing homology-directed genome editing by catalytically active and inactive CRISPR-Cas9 using asymmetric donor DNA. Nature Biotechnology. 34 (3), 339-344 (2016).
- Paix, A., Folkmann, A., Seydoux, G. Precision genome editing using CRISPR-Cas9 and linear repair templates in C. elegans. Methods. 121-122, 86-93 (2017).
- Mello, C., Fire, A.
- Sutter Pipette Cookbook. , Available from: https://www.sutter.com/PDFs/pipette_cookbook.pdf (2017).
- Stiernagle, T.
- Evans, T. C. Transformation and microinjection. WormBook: the online review of C. elegans biology. , (2006).
- Berkowitz, L. A., Knight, A. L., Caldwell, G. A., Caldwell, K. A. Generation of stable transgenic C. elegans using microinjection. Journal of Visualized Experiments. (18), (2008).
- Kim, H., Ishidate, T., et al. A co-CRISPR strategy for efficient genome editing in Caenorhabditis elegans. Genetics. 197 (4), 1069-1080 (2014).
- Arribere, J. A., Bell, R. T., Fu, B. X. H., Artiles, K. L., Hartman, P. S., Fire, A. Z. Efficient marker-free recovery of custom genetic modifications with CRISPR/Cas9 in Caenorhabditis elegans. Genetics. 198 (3), 837-846 (2014).
- Ward, J. D. Rapid and precise engineering of the Caenorhabditis elegans genome with lethal mutation co-conversion and inactivation of NHEJ repair. Genetics. 199 (2), 363-377 (2015).
- Farboud, B., Meyer, B. J. Dramatic enhancement of genome editing by CRISPR/Cas9 through improved guide RNA design. Genetics. 199 (4), 959-971 (2015).
- Wood, A. J., Lo, T. -W., et al. Targeted genome editing across species using ZFNs and TALENs. Science. 333 (6040), 307 (2011).
- Friedland, A. E., Tzur, Y. B., Esvelt, K. M., Colaiácovo, M. P., Church, G. M., Calarco, J. A. Heritable genome editing in C. elegans via a CRISPR-Cas9 system. Nature Methods. 10 (8), 741-743 (2013).
- Dickinson, D. J., Ward, J. D., Reiner, D. J., Goldstein, B. Engineering the Caenorhabditis elegans genome using Cas9-triggered homologous recombination. Nature Methods. 10 (10), 1028-1034 (2013).
- Rehm, E. J., Hannibal, R. L., Chaw, R. C., Vargas-Vila, M. A., Patel, N. H. Injection of Parhyale hawaiensis blastomeres with fluorescently labeled tracers. Cold Spring Harbor Protocols. 2009 (1), (2009).
- Gerberding, M., Browne, W. E., Patel, N. H. Cell lineage analysis of the amphipod crustacean Parhyale hawaiensis reveals an early restriction of cell fates. Development (Cambridge, England). 129 (24), 5789-5801 (2002).
- Browne, W. E., Price, A. L., Gerberding, M., Patel, N. H. Stages of embryonic development in the amphipod crustacean, Parhyale hawaiensis. Genesis. 42 (3), New York, NY. 124-149 (2005).
- Rehm, E. J., Hannibal, R. L., Chaw, R. C., Vargas-Vila, M. A., Patel, N. H. Fixation and dissection of Parhyale hawaiensis embryos. Cold Spring Harbor Protocols. 2009 (1), (2009).
- Rehm, E. J., Hannibal, R. L., Chaw, R. C., Vargas-Vila, M. A., Patel, N. H. In situ hybridization of labeled RNA probes to fixed Parhyale hawaiensis embryos. Cold Spring Harbor Protocols. 2009 (1), (2009).
- Rehm, E. J., Hannibal, R. L., Chaw, R. C., Vargas-Vila, M. A., Patel, N. H. Antibody staining of Parhyale hawaiensis embryos. Cold Spring Harbor Protocols. 2009 (1), (2009).
- Kontarakis, Z., Pavlopoulos, A. Transgenesis in non-model organisms: the case of Parhyale. Methods in Molecular Biology. 1196, Clifton, NJ. 145-181 (2014).
- Kim, H. J., Lee, H. J., Kim, H., Cho, S. W., Kim, J. -S. Targeted genome editing in human cells with zinc finger nucleases constructed via modular assembly. Genome Research. 19 (7), 1279-1288 (2009).
- Qiu, P., Shandilya, H., D'Alessio, J. M., O'Connor, K., Durocher, J., Gerard, G. F. Mutation detection using Surveyor nuclease. BioTechniques. 36 (4), 702-707 (2004).
- Brinkman, E. K., Chen, T., Amendola, M., van Steensel, B. Easy quantitative assessment of genome editing by sequence trace decomposition. Nucleic Acids Research. 42 (22), 168 (2014).
- Tsai, S. Q., Zheng, Z., et al. GUIDE-seq enables genome-wide profiling of off-target cleavage by CRISPR-Cas nucleases. Nature Biotechnology. 33 (2), 187-197 (2015).
- Frock, R. L., Hu, J., Meyers, R. M., Ho, Y. -J., Kii, E., Alt, F. W. Genome-wide detection of DNA double-stranded breaks induced by engineered nucleases. Nature Biotechnology. 33 (2), 179-186 (2015).
- Smith, C., Gore, A., et al. Whole-genome sequencing analysis reveals high specificity of CRISPR/Cas9 and TALEN-based genome editing in human iPSCs. Cell Stem Cell. 15 (1), 12-13 (2014).
- Veres, A., Gosis, B. S., et al. Low incidence of off-target mutations in individual CRISPR-Cas9 and TALEN targeted human stem cell clones detected by whole-genome sequencing. Cell Stem Cell. 15 (1), 27-30 (2014).
- Kim, D., Bae, S., et al. Digenome-seq: genome-wide profiling of CRISPR-Cas9 off-target effects in human cells. Nature Methods. 12 (3), 237-243 (2015).
- Hendel, A., Fine, E. J., Bao, G., Porteus, M. H. Quantifying on- and off-target genome editing. Trends in Biotechnology. 33 (2), 132-140 (2015).
- O'Geen, H., Yu, A. S., Segal, D. J. How specific is CRISPR/Cas9 really. Current Opinion in Chemical Biology. 29, 72-78 (2015).
- Tsai, S. Q., Joung, J. K. Defining and improving the genome-wide specificities of CRISPR-Cas9 nucleases. Nature Reviews Genetics. 17 (5), 300-312 (2016).
- Hoban, M. D., Cost, G. J., et al. Correction of the sickle cell disease mutation in human hematopoietic stem/progenitor cells. Blood. 125 (17), 2597-2604 (2015).
- Simeonov, D. R., Gowen, B. G., et al. Discovery of stimulation-responsive immune enhancers with CRISPR activation. Nature. , (2017).
- Hultquist, J. F., Schumann, K., et al. A Cas9 ribonucleoprotein platform for functional genetic studies of HIV-host interactions in primary human T cells. Cell Reports. 17 (5), 1438-1452 (2016).
- Paix, A., Wang, Y., et al. Scalable and versatile genome editing using linear DNAs with microhomology to Cas9 sites in Caenorhabditis elegans. Genetics. 198 (4), 1347-1356 (2014).
- Lee, K., Mackley, V. A., et al. Synthetically modified guide RNA and donor DNA are a versatile platform for CRISPR-Cas9 engineering. eLife. 6, (2017).
- Minkenberg, B., Wheatley, M., Yang, Y. CRISPR/Cas9-enabled multiplex genome editing and its application. Progress in Molecular Biology and Translational Science. 149, 111-132 (2017).
- Doench, J. G., Fusi, N., et al. Optimized sgRNA design to maximize activity and minimize off-target effects of CRISPR-Cas9. Nature Biotechnology. 34 (2), 184-191 (2016).
- Doench, J. G., Hartenian, E., et al. Rational design of highly active sgRNAs for CRISPR-Cas9-mediated gene inactivation. Nature Biotechnology. , 1-8 (2014).
- Liu, H., Wei, Z., Dominguez, A., Li, Y., Wang, X., Qi, L. S. CRISPR-ERA: a comprehensive design tool for CRISPR-mediated gene editing, repression and activation. Bioinformatics (Oxford, England). 31 (22), 3676-3678 (2015).
- Wu, X., Scott, D. A., et al. Genome-wide binding of the CRISPR endonuclease Cas9 in mammalian cells. Nature Biotechnology. 32 (7), 670-676 (2014).
- Bogenhagen, D. F., Brown, D. D. Nucleotide sequences in Xenopus 5S DNA required for transcription termination. Cell. 24 (1), 261-270 (1981).
- Cozzarelli, N. R., Gerrard, S. P., Schlissel, M., Brown, D. D., Bogenhagen, D. F. Purified RNA polymerase III accurately and efficiently terminates transcription of 5S RNA genes. Cell. 34 (3), 829-835 (1983).
- Chen, B., Gilbert, L. A., et al. Dynamic imaging of genomic loci in living human cells by an optimized CRISPR/Cas system. Cell. 155 (7), 1479-1491 (2013).
- Gagnon, J. A., Valen, E., et al. Efficient mutagenesis by Cas9 protein-mediated oligonucleotide insertion and large-scale assessment of single-guide RNAs. PLoS ONE. 9 (5), 98186 (2014).
- Ren, X., Yang, Z., et al. Enhanced specificity and efficiency of the CRISPR/Cas9 system with optimized sgRNA parameters in Drosophila. Cell Reports. 9 (3), 1151-1162 (2014).
- Ran, F. A., Hsu, P. D., Wright, J., Agarwala, V., Scott, D. A., Zhang, F. Genome engineering using the CRISPR-Cas9 system. Nature Protocols. 8 (11), 2281-2308 (2013).
- Serano, J. M., Martin, A., et al. Comprehensive analysis of Hox gene expression in the amphipod crustacean Parhyale hawaiensis. Developmental Biology. 409 (1), 297-309 (2016).
- Sternberg, S. H., Redding, S., Jinek, M., Greene, E. C., Doudna, J. A. DNA interrogation by the CRISPR RNA-guided endonuclease Cas9. Nature. , 1-17 (2014).
- Lee, K., Conboy, M., et al. Nanoparticle delivery of Cas9 ribonucleoprotein and donor DNA in vivo induces homology-directed DNA repair. Nature Biomedical Engineering. 1 (11), 889-901 (2017).









































