

Summary
Nous présentons une méthode pour quantifier la teneur en amidon dans les primordiums ovaire dans sweet cherry (Prunus avium L.) au cours de la dormance hivernale à l’aide d’un système d’analyse image combiné avec des techniques histochimiques.
Abstract
Changements dans l’amidon dans des petites structures sont associées à des événements clés au cours de plusieurs processus de développement de la plante, y compris la phase reproductive de la pollinisation à la fécondation et le début de la fructification. Toutefois, des variations dans l’amidon pendant la différenciation de la fleur ne sont pas totalement connues, principalement en raison de la difficulté de quantifier la teneur en amidon dans les structures en particulier les petits des primordia fleur. Nous décrivons ici une méthode pour la quantification de l’amidon dans les primordiums ovaire de sweet cherry (Prunus avium L.) en utilisant un système d’analyse image attaché à la loupe, qui permet concernant les changements de teneur en amidon avec les différentes phases de la dormance de l’automne au printemps. À cette fin, l’état de dormance des bourgeons floraux est déterminée en évaluant la croissance des bourgeons des tiges transférée dans des conditions contrôlées à des moments différents de l’heure d’hiver. Pour la quantification de l’amidon dans les primordiums ovaire, boutons floraux sont séquentiellement recueillies, fixe, incorporé dans la paraffine, sectionnées et colorées avec j’ai2Kl (iodure de potassium iodée). Préparations sont observées au microscope et analysées par un analyseur d’image qui la distingue clairement l’amidon de l’arrière-plan. Amidon contenus valeurs sont obtenues en mesurant la densité optique de l’image qui correspond à l’amidon coloré, considérant la somme de la densité optique de chaque pixel comme une estimation de la teneur en amidon du cadre étudié.
Introduction
Tempérées ligneuses vivaces s’adaptent aux saisons en modulant leur croissance et leur développement. Alors qu’ils développent au printemps et en été, ils cessent de croître au cours de l’automne pour aller en dormance en hiver1. Bien que la dormance leur permet de survivre aux basses températures hivernales, réfrigération est une condition sine qua non pour un bon débourrement au printemps2. Les répercussions importantes de dormance en production fruitière tempérées et des forêts ont conduit à divers efforts visant à déterminer et à prévoir la période de dormance3. Espèces d’arbres fruitiers, expériences empiriques transférant des pousses au forçage des conditions et des prévisions statistiques basées sur les données de la floraison sont les approches actuelles de déterminer la date de la rupture de la dormance, ce qui permet aux chercheurs d’estimer la exigences pour chaque cultivar de refroidissement. Cependant, comment déterminer l’état de dormance basé sur des processus biologiques reste peu clair3.
Floraison en fruitiers tempérés, comme sweet cherry (Prunus avium L.), se produit une fois par an et dure environ une quinzaine de jours. Cependant, les fleurs commencent à se différencier et de développer une dizaine de mois plus tôt, au cours de l' été précédent4. Les primordiums fleur cesse de croître au cours de l’automne à rester dormant à l’intérieur des bourgeons en hiver. Au cours de cette période, chaque cultivar doit accumuler une exigence particulière paralysant pour bonne floraison4. Malgré l’absence de changements phénologiques dans les bourgeons en hiver, fleur primordiums sont physiologiquement actives en période de dormance, et l’accumulation des températures de réfrigération a été récemment associée à la dynamique de l’accumulation de l’amidon ou diminuer dans les cellules du primordium ovaire, offrant une nouvelle approche pour la détermination de dormance5. Toutefois, la petite taille et l’emplacement du primordium ovaire nécessitent une méthodologie particulière.
L’amidon est le glucide de stockage importants en espèces de plantes ligneuses6. Ainsi, les changements dans l’amidon ont été associés à l’activité physiologique des tissus fleur, qui ont besoin de glucides pour soutenir leur développement7,8. Différents événements clés durant le processus de reproduction sont également liés aux variations de la teneur en amidon dans les différentes structures florales, tels que les anthères méiose9, la croissance des tubes polliniques dans le style ou ovule fécondation10. Des techniques histochimiques permettent la détection de l’amidon dans chaque tissu particulier des primordia fleur en période de dormance. Toutefois, la difficulté reste à quantifier cette fécule pour permettre à la suite de son modèle d’accumulation/diminution avec le temps, ou en comparant l’amidon contenu entre les tissus, les cultivars ou des années. Cela est dû à la petite quantité de tissu disponible pour les techniques d’analyse11. Comme alternative, analyse d’image liée à la microscopie12 permet la quantification de l’amidon dans très petits échantillons de plantes tissus13.
Microscopie et analyse d’images de la combinaison des approches ont été utilisées pour quantifier le contenu des différents composants dans les tissus végétaux, tels que de la callose14, microtubes15, ou16, l’amidon en mesurant la taille de la zone teintée par le particulier les taches. Pour l’amidon, il peut être facilement détecté à l’aide de l’iodure de potassium iodée (j’ai2KI) réaction17. Cette méthode est très spécifique ; J’ai2KI s’intercale au sein de la structure laminaire des grains d’amidon et forme une couleur bleue ou rouge-brun foncé, selon la teneur en amylose de l' amidon18. Sections colorées avec I tache KI2montrent un contraste suffisant entre l’amidon et le tissu de fond, ce qui permet une détection de l’amidon sans équivoque et la quantification ultérieure par l’image analyse système19. Bien que ce colorant n’est pas stoechiométrique, l’accumulation d’iode est proportionnelle à la longueur de la molécule d’amidon, qui peut varier fortement de17. Ainsi, la taille de la zone tachée, exprimée en nombre de pixels peut-être ne pas refléter avec précision la teneur en amidon, étant donné les différences de hautes teneur en amidon a pu être trouvées entre les champs avec des zones colorées de la même taille. Comme alternative, la teneur en amidon peut être évaluée en mesurant la densité optique des granules colorés sur des images noir et blanc obtenus à partir du microscope, il a été signalé dans différents tissus abricot8,13 , 19, avocat10,20et olive21.
Nous décrivons ici une méthodologie qui allie la détermination expérimentale de l’état de dormance de la quantification de la teneur en amidon dans le tissu ovarien de primordium de l’automne au printemps en cerisier, offrant un nouvel outil pour la compréhension et la prévision de dormance basée sur l’étude des mécanismes biologiques liés à la dormance.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. Collection matière dormance détermination et plante
- Les boutons floraux dans le domaine de l’échantillon. Études de dormance sont des expériences à long terme et adulte arbres assez grand pour recueillir les bourgeons et les pousses tout l’hiver sans compromettre le développement des arbres au printemps prochain. Gestion du verger spécial pourrait être requise selon le système de formation ; ainsi, l’élagage peut être moins sévère que pour fins de production de fruits.
- Chaque semaine, dès le début de l’automne jusqu’au début du débourrement, collecter et peser 10 boutons floraux.
- Fixer les boutons floraux en les plaçant dans un tube de verre de 10 mL avec un bouchon et tremper les échantillons dans une solution de fixation de l’acide acétique/éthanol (3:1) pendant au moins 24 h à 4 ° C. Ensuite, jeter le fixateur et ajouter 75 % éthanol, généreusement s’assurer qu’il couvre les échantillons. Les échantillons peuvent être conservés dans cette solution à 4 ° C jusqu'à l’utilisation.
Remarque : L’infiltration sous vide peut être utilisée pour enlever les bulles d’air à l’intérieur de le œuf et les empêcher de flotter. Cela facilite la pénétration du fixateur dans les tissus mais pourrait endommager la structure des tissus. Essayer de l’éviter si pas nécessaire.
- Les pousses dans le domaine de l’échantillon.
- Chaque semaine, dès le début de l’automne jusqu’au début du débourrement, prendre trois tiges de 15 à 30 cm de longueur et 5 mm de diamètre, contenant au moins 10 boutons de fleurs. Placez-les sur la mousse de fleuriste imbibées d’eau dans une chambre de croissance à 22 ± 1 ° C, avec une photopériode de lumière de 12 h.
- Après 10 jours dans la chambre de croissance, pick et peser 10 boutons floraux de pousses.
- Évaluer la croissance des bourgeons et de déterminer l’état de dormance. La période d’échantillonnage doit être adaptée aux conditions de l’emplacement. Dans les conditions du verger (Saragosse, Espagne, 41 ° 44'30 "N-0 ° 47'00" W et 220 m d’altitude), l’échantillonnage des pousses a été réalisée du 30 novembre jusqu'à la fin de février ou début mars.
- Toutes les semaines évaluer la réponse des bourgeons floraux pour les conditions de croissance approprié de la chambre, dès le début de l’automne jusqu’au débourrement au printemps, en comparant le poids des 10 bourgeons cueillies sur le terrain.
- Si il n’y a pas de différences ou ces différences sont moins de 30 %, sachez que les bourgeons ne ne sont pas acquittés de leurs exigences de refroidissement et endodormant22. Si les différences sont plus de 30 %, considèrent que les bourgeons sont acquittés de leurs exigences de refroidissement et ecodormant22.
2. plante préparation du matériel pour la Quantification de l’amidon
- Certains bourgeons fixes environ six de chaque échantillonnage datent (voir l’étape 1.1). Selon les cultivars, chaque bourgeon de fleur de cerise contient jusqu'à cinq primordiums de fleur.
- Retirez les écailles externes et le bourgeon sur verre d’un horloger avec 75 % d’éthanol pour l’empêcher de se dessécher.
- Disséquer le bourgeon et extraire au moins un primordium de fleur par bourgeon avec l’aide de pinces de précision et un bistouri ophtalmologique sous un microscope stéréoscopique. Le primordium de fleur peuvent être conservés dans un tube de verre de 10 mL avec 75 % d’éthanol à 4 ° C, ou procéder immédiatement à l’étape suivante.
- Déshydrater les échantillons en une série d’alcool butylique tertiaire.
- Remplacer la solution d’éthanol 75 % avec 10 mL d’alcool butylique tertiaire de 75 % (TBA) couvrir généreusement les échantillons et les incuber pendant 1,5 h. Une pipette Pasteur peut être utile de se défaire de la solution.
- Répétez l’étape 2.2.1, ajouter TBA avec augmentation des concentrations (85 %, 95 % et 100 %, v/v ; et 3 x avec pure TBA) dans un poêle séchage à 30 ° C, avec extraction d’air, depuis TBA pur crystalizes à température ambiante (< 20 ° C) et est très volatile et toxique. Si TBA crystalizes avec l’échantillon, il endommagerait les tissus.
- Incorporer les échantillons en paraffine18.
- Faire fondre de la paraffine en introduisant les perles de la paraffine dans le poêle séchage à 60 ° C, avec extraction d’air le jour précédent. Les perles de paraffine peuvent être également fondus sur une plaque chauffante, mais faire en sorte que la paraffine liquide est à 60 ° C et non à une température plus élevée, afin d’éviter des lésions tissulaires.
- Remplacer le TBA avec un mélange d’huile de l’ATB et paraffine (1:1) et incuber pendant 24 h à l’intérieur d’un four de séchage à 60 ° C. Ensuite, remplacez le mélange de TBA et huile de paraffine avec pur faire fondre la paraffine et incuber pendant au moins 6 h, à l’intérieur d’un four de séchage à 60 ° C. Répéter 2 x et incuber la dernière modification pour au moins 4 à 6 jours.
- Placer chaque échantillon sur un petit moule métallique sur une surface de chaleur, incorporé dans la cire de paraffine et rythme la cassette d’encastrement. Placez-le sur une surface froide et retirez le bloc une fois que la cire est solidifiée.
- L’article et réhydrater les préparatifs.
- Préparer la colle de Haupt : dissoudre 1 g de gélatine ordinaire de Knox dans 100 mL d’eau distillée à 30 ° C ; Ensuite, ajouter 2 g de cristaux lâche de phénol (C6H5OH) et 15 mL de glycérol. Étaler l’adhésif de la Haupt sur une lame de verre avec un pinceau et ajouter une goutte d’une solution de formaldéhyde de 1 %.
- Section chaque paraffine bloquer à 10 µm dans un microtome rotatoire et déposer les éléments sur la lame de verre recouvert d’adhésif de Haupt.
- Placez les lames de verre avec les sections au-dessus d’une surface de chaleur à 35-40 ° C à sec ; puis, déplacez les lames de verre pour une grille coulissante.
- Préparer les solutions dewax et réhydratation. Placer 200 mL de Histoclear II dans trois plats de verre colorées, un autre avec Histoclear II:ethanol (1:1, v/v), une série de l’éthanol (100 %, 70 %, 40 %, v/v) et un lavage final de l’eau distillée.
- Déparaffinage les sections avec trois lavages, chaque 5 min, dans II Histoclear et Histoclear II:ethanol (1:1, v/v). Placer la grille coulissante à l’intérieur de la vaisselle de verre-coloration, veiller à ce que la solution couvre entièrement les diapositives et, ensuite, de déplacer la grille coulissante d’une solution à l’autre.
- Réhydrater les sections de la série suivante de lave-2 min : une série de l’éthanol (100 %, 70 %, 40 %, v/v) et un lavage final de l’eau distillée. Enfin, faire sécher les lames de verre à température ambiante.
- Tacher les sections18.
- J’ai préparer tache KI2: dissoudre 2 g d’iodure de potassium (KI) et 0,2 g d’iode (j’ai2) dans 100 mL d’eau distillée. Appliquez une goutte de frais j’ai2KI sur l’ou les sections pendant 5 min et, ensuite, jetez l’excédent de la tache par absorber avec un papier buvard. Rapidement passer à l’étape suivante.
- Appliquer une petite goutte d’un support de montage synthétique, placer un petit couvercle en verre sur le dessus et appuyez fort. Une fois que le support de montage sèche, observer au microscope lumineux-zone pour une évaluation préliminaire de l’amidon de l’ovaire.
Remarque : Cette étape n’est pas obligatoire, mais un meilleur contraste entre l’amidon et l’arrière-plan est obtenu. Si l’article doit être réutilisé avec autres taches après quantification de l’amidon, placez le couvercle en verre sur la goutte de colorant et jetez l’excédent par absorber avec un papier buvard (voir 2.5.1). Puis, après quantification de l’amidon, laver la je2KI tache avec l’eau distillée et place la préparation sur un feu de surface à 35-40 ° C jusqu’au sec.
3. quantification de la teneur en amidon
- Calibrer les conditions optiques.
Remarque : Les niveaux de détection utilisés par l’analyseur d’image pour détecter l’amidon coloré sont dépend directement des conditions de luminosité et grossissement du microscope ; ainsi, fixer ces conditions pour toutes les préparations évaluées. Adapter le rajustement proposé ici au microscope disponible et des conditions de lumière.- Ajuster le diaphragme d’ouverture à la 20 X grossissement et le réglage de la luminosité ou l’intensité lumineuse.
- Assurez-vous qu’il n’y a pas de filtres sur le porte-filtre, puis sélectionnez une condition de lumineux-zone dans le microscope. Sélectionnez un grossissement approprié (par exemple, 40 X pour primordium ovaire cerise).
- Contrôler les conditions d’acquisition d’image. Ajuster les réglages de l’appareil avec une préparation teinté sans tissu via Image | Acquisition | Vue avant.
- Fixer la luminosité à 50 %, le gain à 1,0 x et les indicateurs d’histogramme le gamma de l’évaluer à 1,00 et le contraste à 0 - 100, alignant avec les limites de l’histogramme de distribution de luminosité.
- Activer la fonction de surexposition/sous-exposition et ajuster la durée d’exposition à la limite de surexposition.
- Appliquer la fonction de balance des blancs à l’image complète pour afficher tous les composants de couleur neutre de l’image sans n’importe quel ton de la couleur et la correction de l’ombrage à la totalité de l’image pour créer une image homogène et corrigée.
- Étalonner le système de l’analyse d’Image pour obtenir les valeurs de contrôle le niveau de gris (0, noir ; 255, blanc) de différentes densités optiques (OD) qui sont obtenues par les valeurs de transmission (T).
- Acquérir une image d’une préparation teintée sans tissu, a examiné le contrôle blanc et mesurer le niveau de gris de l’image en noir et blanc par mesure | Gris de mesure | Calibrer le gris | Valeur de référence = 0 | Mesure | Étalonner | OK. Ceci correspond à un facteur de transmission de 100 % ; ainsi, une densité optique de 0, selon OD = 2 - log T.
- Acquérir une image de la même préparation avec un filtre de 4N, qui réduit la quantité de lumière 4 x et mesure le niveau de gris de l’image en noir et blanc par mesure | Gris de mesure | Calibrer le gris | Valeur de référence = 0,6 | Mesure | Étalonner | OK. Ceci correspond à un facteur de transmission de 25 % ; ainsi, une densité optique de 0,6, selon OD = 2 - log T.
- Acquérir une image de la même préparation sans noir lumière, réfléchie, et mesurer le niveau de gris de l’image en noir et blanc par mesure | Gris de mesure | Calibrer le gris | Valeur de référence = 1 | Mesure | Étalonner | OK. Ceci correspond à un facteur de transmission de 0 % ; ainsi, une densité optique de 1, selon OD = 2 - log T.
- Détecter l’amidon.
- Acquérir une image de couleur du champ à mesurer au format TIFF avec une résolution d’au moins 300 points par pouce (PPP).
- Créer une image binaire correspondant à la zone tachée. Définir les seuils de trois couleurs (valeurs comprises entre 0 - 255 pour chacun) jusqu'à ce que l’image binaire reflète exactement les granules d’amidon colorés observés, via Image | Détecter | Sélectionnez les seuils de rouge, de bleu et de vert | OK. Faire des comparaisons visuelles à plusieurs reprises dans différentes préparations et tissus pour régler les niveaux de détection finale. Stocker et utiliser ces niveaux pour toutes les préparations.
- Quantifier l’amidon. Convertir l’image originale de la couleur dans une image noir et blanc avec le système d’analyse des images. Utiliser l’image binaire comme un masque superposé sur l’image en noir et blanc par Image | Edit binaire. Mesurer la somme de la densité optique de chaque pixel sous le masque via mesure | Niveau de gris | OK et considérer cette valeur comme la teneur en amidon dans le champ mesuré.
- Répétez les étapes 3.4.1 - 3.4.3 dans quatre endroits des primordia ovaire pour obtenir une valeur représentative de l’amidon dans l’ovaire de primordia de fleur.
- Répétez les étapes 3.4.1 - 3.4.3 et étape 3.4.5 dans différentes fleurs de chaque date de collecte.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Études de dormance exigent la détermination du moment où les exigences de refroidissement sont réunies. Malgré l’absence de changements phénologiques pendant l’hiver dans des conditions naturelles (Figure 1 a), les cerisiers ne récupèrent pas la capacité de croissance dans des conditions appropriées jusqu'à ce qu’ils passent un certain temps sous des températures basses. Le transfert régulier des pousses à une chambre de conditions contrôlées (Figure 1 b) au cours de l’heure d’hiver a permis l’évaluation de l’état de dormance des bourgeons floraux. L’évaluation de la croissance des bourgeons de fleur a été faite par la mesure de l’augmentation du poids du bourgeon. Alors que, en période de dormance, aucun changement pouvait être observé après 10 jours de conditions favorables (Figure 1), une fois surmontée la dormance, les bourgeons se gonflaient et fait irruption dans la chambre de croissance (Figure 1). Les résultats de cette analyse a permis à l’état de dormance des bourgeons à établir. En raison des températures différentes au cours de l’hiver, la dormance a été surmontée à des dates différentes, selon les années. Alors que, pendant la première année de l’étude, la rupture de la dormance a eu lieu en janvier, la deuxième année a présenté un hiver plus doux ; ainsi, l’accomplissement de refroidissement s’est produit environ trois semaines plus tard, en février.
Cerise douce porte boutons floraux dans les dents, où le bourgeon apical est un bourgeon végétatif et les bourgeons latéraux sont les boutons floraux (Figure 1C et 1D). Bourgeons indifférenciées ont commencé à se différencier en fleurs ou bourgeons végétatifs à la fin de l’été, et lorsqu’ils entrent dans la dormance en hiver, plusieurs primordiums fleur restent à l’intérieur de le œuf, protégée par nombreuses écailles vertes et couvertes par des échelles extérieures brun ( Figure 1 et Figure 2 a). La dissection du bourgeon floral a montré les primordiums petite fleur à l’intérieur (Figure 2 a). Chaque bouton floral figurant un à cinq primordiums des fleurs individuelles (Figure 2 b). Malgré la petite taille du primordium de chaque fleur, toutes les parties d’une fleur sont différenciées et peuvent être distingués : le pistil, les anthères, les pétales et les sépales (Figure 2). L’utilisation des techniques histochimiques (alcool : acétique [3:1] fixation, enrobage de paraffine, microtome de sectionnement et de coloration d’amidon à base d’iode) a permis la distribution de l’amidon dans la fleur tissus primordium à observer (Figure 2D ).
Amidon dans le primordium de l’ovaire a été quantifié dans chaque section. Quatre mesures de µm 13372, à un grossissement de 40 x, représentaient la disposition générale de l’amidon dans le primordium d’ovaire sweet cherry (Figure 3 a). Les granules d’amidon sont distingués nettement de l’arrière-plan après j’ai2KI coloration (Figure 3 b). L’amidon a été identifié par le système d’analyse des images, en ajustant la couleur seuils de rouge, vert et bleu jusqu'à ce que toutes les granules d’amidon observés étaient couverts par l’image binaire créé par le système basé sur les paramètres de couleurs définis (Figure 3). Les valeurs de teneur en fécule obtenus étaient le résultat de la mesure de la densité optique de chaque pixel sous le masque sur l’image en noir et blanc (Figure 3D).
La quantification de l’amidon a révélé un modèle cohérent de fécule dynamique au cours de l’hiver (Figure 4). Régulièrement, la quantité d’amidon dans le début de l’hiver a présenté une valeur de densité optique de moins de 40 0003, tandis que le maximum atteint une valeur comprise entre 120 000 et 140 000 dans les deux ans. Alors que la valeur maximale est atteinte en janvier au cours de la première année (Figure 4 a), elle a eu lieu en février à la deuxième année (Figure 4 b). Contrastées de ces résultats avec le teint de la dormance, le maximum d’amidon a eu lieu concomitamment avec la paralysant l’accomplissement dans les deux ans.
Cette approche nécessite la détermination de l’état de dormance (Figure 5 a) concomitamment avec la quantification de l’amidon sur le tissu de l’ovaire (Figure 5 b) afin d’encadrer les changements de la teneur en amidon par rapport à la dormance.

Figure 1 : montage expérimental pour la détermination de l’état de dormance des bourgeons de fleur de cerisier. Branches (A), au cours de l’hiver, montrent les bourgeons dormants fermés et couverts par des écailles bruns foncés. (B), ce panneau montre les pousses, transférés à la chambre de croissance. À la mi-janvier, certains cultivars sont restés dormants avec les bourgeons encore fermé (flèche blanche), tandis que d’autres ont pu se développer, montrant les bourgeons soufflées (flèche noire). (C) ce panneau montre un détail d’un tournage avec boutons floraux dormants situé latéralement et un bourgeon unique situé en position apicale dans l’épi. (D), ce panneau montre un détail d’un tournage, une fois la dormance a été surmontée après 10 jours dans la chambre de croissance, montrant le gonflement des bourgeons. S’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.
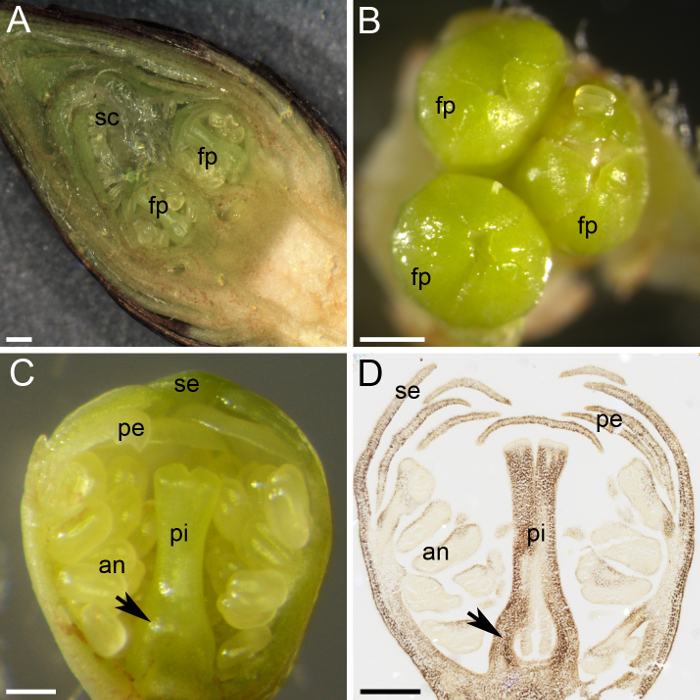
Figure 2 : préparation du matériel pour la quantification de l’amidon des plantes. (A), ce tableau montre une section transversale d’un bourgeon de fleur, montrant deux primordia de fleur (fp) protégée par nombreuses échelles (sc). (B) trois primordiums fleur (fp) se réunissent dans un bourgeon floral. (C) ce panneau montre une section transversale d’un primordium de fleur, avec tous les verticilles différenciées : sépales (se), des pétales (pe), anthères (un) et pistil (pi). Le primordium de l’ovaire se distingue à la base du pistil (flèche). (D), une section centrale d’un primordium de fleur a été recueillie et fixée en janvier, incorporée dans la cire de paraffine, sectionnée et colorée avec j’ai2KI (brun foncé) pour l’amidon. Ce panneau indique le primordium de l’ovaire (flèche). Les barres d’échelle sont 500 µm de panneaux A et B et 200 µm dans les séries C et D. S’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.

Figure 3 : amidon de quantification dans primordium ovaire sweet cherry. (A), ce panneau affiche une section centrale d’un primordium ovaire teinté avec I2KI, montrant les quatre cadres dans lequel amidon contenu a été mesurée. (B), ce tableau montre un détail du primordium ovaire. Les granules d’amidon sont colorées en brun foncé. (C), ce panneau affiche une image en pseudo-couleur dans lequel amidon correspond aux différentes nuances de bleu. (D), ce panneau indique un masque binaire image j’ai couvrant2KI colorés à l’amidon (bleu) sur l’image originale noir et blanc. La densité optique est mesurée dans les pixels de l’image originale, caché derrière le masque. Les barreaux de l’échelle sont de 100 µm en groupe A et 20 µm de panneaux B - D. S’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.

Figure 4 : résultats représentatifs de la quantification de l’amidon dans les primordiums ovaire cerise prélevés mensuellement à partir de l’automne jusqu’au printemps au cours de deux années de conditions de température différentes hiver. (A), ce panneau montre les résultats des années 2010-2011, qui a eu un hiver froid. L’accomplissement paralysant (flocon) a eu lieu en janvier, en même temps avec le maximum d’amidon. (B), ce panneau affiche les résultats des années 2011-2012, qui a eu un hiver doux. L’accomplissement paralysant (flocon) a eu lieu en février, en même temps avec le maximum d’amidon. Les valeurs sont la moyenne ± l’écart-type de la moyenne. S’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.
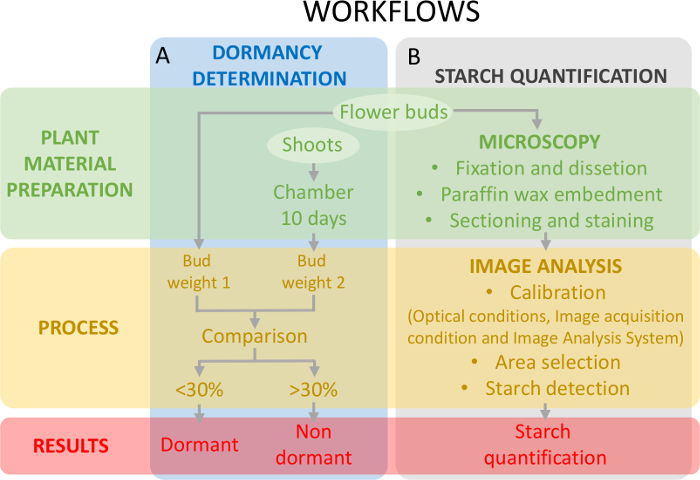
Figure 5 : schéma du plan expérimental pour évaluer l’état de dormance des bourgeons et la quantification de l’amidon dans les primordiums ovaire dans sweet cherry. (A), ce panneau montre les flux de travail de la détermination du statut de dormance : la préparation de matériel de plantes, le processus et les résultats obtenus. (B), ce panneau montre les flux de travail de la quantification de l’amidon : la préparation histochimique des bourgeons d’une observation microscopique de l’amidon, la détection de l’analyse d’image d’amidon et de la quantification de l’amidon. S’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Dormance des plantes ligneuses vivaces présente des implications claires dans la production de fruits et des forêts au changement climatique, même si le processus biologique derrière la dormance reste floue. Études de dormance peuvent être abordés sous différents angles, mais la recherche à la recherche d’un marqueur biologique de dormance hivernale s’est intensifiée ces dernières années. Cependant, la plupart des tentatives pour trouver un indicateur sans équivoque montrant quand un bourgeon a cassé la dormance ont été infructueuses3. La méthodologie décrite dans les présentes, alliant des techniques histochimiques18 image analyse, a été très utile d’examiner la relation entre les réserves de glucides d’un tissu particulier et son activité physiologique dans les bourgeons de cerisier pendant les différentes phases de dormance5 et peut être également appliqué aux autres espèces et les tissus10,13,20.
Réserves de glucides sous forme d’amidon jouent un rôle important dans le développement des fleurs et le processus de reproduction7,10,13,20,21,23 et la saisonnalité des vivaces ligneuses tempérées6. Différentes études sur la dormance ont prêté attention à l’amidon dans les bourgeons24. Toutefois, en raison de leur petite taille, plusieurs bourgeons ensemble sont requis pour l’utilisation des méthodes d’analyse quantitatives et la détection des variations quantitatives dans des cellules ou des tissus particuliers est limitée par l’effet de masquage des cellules environnantes. La combinaison des techniques histochimiques avec système d’analyse de l’image offre une bonne occasion d’étudier les changements de la teneur en amidon des structures différentes à l’intérieur de le œuf.
Cette méthode a la limitation de la prévention de la quantification de la teneur exacte de l’amidon dans le tissu mais permet à la teneur en amidon relative à quantifier à suivre les changements quantitatifs amidon plus de temps de25 et de comparer l’amidon contenu de différents tissus13,20, cultivars ou ans5. Afin de permettre la comparaison correcte entre les valeurs de densité optique entre les champs, des tissus et des bourgeons, le calibrage du système (luminosité, coloration intensité et grossissement) et la fixation de seuils doivent être établis avec précision de couleur , stocké et utilisé pour toutes les préparations.
Amidon de coloration et de quantification basée sur l’iode de potassium permet l’utilisation ultérieure d’autres taches après le lavage de la section. Ainsi, différentes analyses peuvent être effectuées sans préparations supplémentaires et montage synthétique médias ne sont pas utilisés5,20. De même, de mesures morphométriques26 peut être fait après les mêmes préparations, ce qui permet du patron d’accumulation d’amidon pour être encadré par rapport à la croissance des différentes structures5,13. La méthode peut être adaptée à d’autres structures ou espèces à l’ajustement des niveaux de couleur qui utilise l’analyseur pour détecter l’amidon, qui peut permettre l’étude des autres processus de développement qui impliquent des changements dans la teneur en amidon en petits groupes de cellules.
La relation entre la libération de dormance et amidon accumulation dans les primordiums ovaire dévoilée par l’utilisation de cette méthode fournit une base solide pour comprendre les fondements biologiques de la dormance et paralysant exigences5. Cependant, amidon quantification par analyse d’image sur des sections de paraffine pourrait se pour révéler très lourd et fastidieux à estimer les besoins de refroidissement d’un grand nombre de cultivars. Futurs efforts doivent se concentrer sur l’étude des indicateurs biologiques fiables qui peuvent facilement indiquent l’état de dormance de l’arbre. Pendant ce temps, le patron des variations de l’amidon en période de dormance peut être utilisé pour encadrer davantage les études physiologiques et génétiques, et la combinaison des techniques histochimiques avec analyse d’image décrit ci-après pourrait servir dans d’autres cultures ligneuses vivaces à quantifier la teneur en amidon des tissus différents en ce qui concerne la dormance.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Les auteurs n’ont rien à divulguer.
Acknowledgments
Les auteurs remercient chaleureusement Maria Herrero et Eliseo Rivas pour leur discussion utile et des conseils. Ce travail a été soutenu par le Ministerio de Economía y Competitividad — fonds européen de développement régional, l’Union européenne [numéro de licence BES-2010-037992 E. F.] ; l’Instituto Nacional de Investigación y Tecnología Agraria y Alimentaria [grant nombre 00015-RFP2015-00, RTA2014-00085-00, RTA2017-00003-00] ; et le Gobierno de Aragón — Fonds Social européen, l’Union européenne [Grupo Consolidado A12-17R].
Materials
| Name | Company | Catalog Number | Comments |
| Precision scale | Sartorius | CP225D | |
| Stereoscopic microscope | Leica Microsystems | MZ-16 | |
| Drying-stove | Memmert | U15 | |
| Paraffin Embedding station | Leica Microsystems | EG1140H | |
| Rotatory microtome | Reichert-Jung | 1130/Biocut | |
| Microtome blade | Feather | S35 | Stainless steel |
| Bright field microscope | Leica Microsystems | DM2500 | |
| Digital Camera | Leica Microsystems | DC-300 | |
| Image Analysis System | Leica Microsystems | Quantiment Q550 |
References
- Kurokura, T., Mimida, N., Battey, N. H., Hytönen, T. The regulation of seasonal flowering in the Rosaceae. Journal of Experimental Botany. 64 (14), 4131-4141 (2013).
- Rohde, A., Bhalerao, R. P.
- Fadón, E., Rodrigo, J. Unveiling winter dormancy through empirical experiments. Environmental and Experimental Botany. 152, 28-36 (2018).
- Fadón, E., Rodrigo, J., Herrero, M. Is there a specific stage to rest? Morphological changes in flower primordia in relation to endodormancy in sweet cherry (Prunus avium L.). Trees - Structure and Function. , In Press (2018).
- Fadón, E., Herrero, M., Rodrigo, J. Dormant flower buds actively accumulate starch over winter in sweet cherry. Frontiers in Plant Science. 9 (171), (2018).
- Loescher, W. H., Mccamant, T., Keller, J. D. Carbohydrate reserves, translocation and storage in woody plant roots. HortScience. 25 (3), 274-281 (1990).
- Hedhly, A., et al. Starch turnover and metabolism during flower and early embryo development. Plant Physiology. , (2016).
- Rodrigo, J., Hormaza, J. I., Herrero, M. Ovary starch reserves and flower development in apricot (Prunus armeniaca). Physiologia Plantarum. 108 (1), 35-41 (2000).
- Julian, C., Rodrigo, J., Herrero, M. Stamen development and winter dormancy in apricot (Prunus armeniaca). Annals of Botany. 108 (4), 617-625 (2011).
- Alcaraz, M. L., Hormaza, J. I., Rodrigo, J. Pistil starch reserves at anthesis correlate with final flower fate in avocado (Persea americana). PLoS One. 8 (10), e78467 (2013).
- Smith, A. M., Zeeman, S. C. Quantification of starch in plant tissues. Nature Protocols. 1 (3), 1342-1345 (2006).
- Eliceiri, K. W., et al.
- Rodrigo, J., Herrero, M. Influence of intraovular reserves on ovule fate in apricot (Prunus armeniaca L.). Sexual Plant Reproduction. 11, 86-93 (1998).
- Zhou, J., Spallek, T., Faulkner, C., Robatzek, S. CalloseMeasurer: A novel software solution to measure callose deposition and recognise spreading callose patterns. Plant Methods. 8 (1), (2012).
- Faulkner, C., et al. An automated quantitative image analysis tool for the identification of microtubule patterns in plants. Traffic. 18 (10), 683-693 (2017).
- Kuhn, B. F. Determination of starch in ovules of the sour cherry cv. "Stevnsbaer.". European Journal of Horticultural Science. 71 (3), 120-124 (2006).
- Johansen, D. A. Plant microtechnique. , McGraw-Hill. New York, NY. (1940).
- Ruzin, S. E. Plant microtechnique and microscopy. , Oxford University Press. New York, NY. (1999).
- Rodrigo, J., Rivas, E., Herrero, M. Starch determination in plant tissues using a computerized image analysis system. Physiologia Plantarum. 99 (1), 105-110 (1997).
- Alcaraz, M. L., Hormaza, J. I., Rodrigo, J. Ovary starch reserves and pistil development in avocado (Persea americana). Physiologia Plantarum. 140 (4), 395-404 (2010).
- Suarez, C., Castro, A. J., Rapoport, H. F., Rodriguez-García, M. I. Morphological, histological and ultrastructural changes in the olive pistil during flowering. Sexual Plant Reproduction. 25, 133-146 (2012).
- Lang, G. A., Early, J. D., Martin, G. C., Darnell, R. L. Endodormancy, paradormancy, and ecodormancy - Physiological terminology and classification for dormancy research. HortScience. 22 (3), 371-377 (1987).
- Hedhly, A., Vogler, H., Eichenberger, C., Grossniklaus, U. Whole-mount clearing and staining of arabidopsis flower organs and siliques. Journal of Visualized Experiments. 2018 (134), 1-10 (2018).
- Kaufmann, H., Blanke, M. Changes in carbohydrate levels and relative water content (RWC) to distinguish dormancy phases in sweet cherry. Journal of Plant Physiology. 218 (July), 1-5 (2017).
- Herrero, M., Dickinson, H. G. Pollen-pistil incompatibility in Petunia hybrida: changes in the pistil following compatible and incompatible intraspecific crosses. Journal of Cell Science. 36, 1-18 (1979).
- Carpenter, A. E., et al. CellProfiler: image analysis software for identifying and quantifying cell phenotypes. Genome Biology. 7 (10), R100 (2006).
