

Summary
Vi presenterer en metodikk for å kvantifisere stivelsen innhold i eggstokken primordia i søte kirsebær (Prunus avium L.) under Vinter dormancy ved hjelp av et bilde analysesystem kombinert med histochemical teknikker.
Abstract
Endringer i stivelse i små strukturer knyttes til viktige hendelser under flere anlegg-utviklingsprosesser, inkludert den reproduktive fasen fra pollinering befruktning og utbruddet av fruiting. Imidlertid er variasjoner i stivelse under blomst differensiering ikke helt kjent, hovedsakelig på grunn av vanskelighetene med å kvantifisere stivelsen innhold i spesielt små strukturer av blomst primordia. Her beskriver vi en metode for kvantifisering stivelse eggstokk primordia av søte kirsebær (Prunus avium L.) ved hjelp av et bilde analysesystem knyttet til mikroskopet, hvilke innrømmer knyttet endringene i stivelsen innhold med de ulike fasene uvirksom fra høsten til våren. For dette formålet bestemmes dormancy status blomsterknopper ved å evaluere bud veksten av skudd overført til kontrollerte forhold på ulike øyeblikk vinteren. For kvantifisering stivelse eggstokk primordia, blomsterknopper sekvensielt samlet, fast, innebygd i parafinvoks, delt og farget med jeg2Kl (kalium iodide-jod). Forberedelser er observert under mikroskopet og analyseres av et bilde analysator som skiller stivelse fra bakgrunnen. Stivelse innhold verdiene hentes ved å måle den optiske densitet for bildet som tilsvarer farget stivelse, vurderer summen av den optiske densitet for hver piksel som en vurdering av stivelsen innhold rammen studerte.
Introduction
Temperert woody stauder tilpasse seg årstidene ved modulerende deres vekst og utvikling. Mens de utvikle om våren og sommeren, slutte de å vokse i løpet av høsten å gå sovende i vinter1. Selv om dormancy tillater dem å overleve på lave vintertemperaturer, er kjøling en forutsetning for en riktig budburst i vår2. De viktige implikasjonene uvirksom i tempererte frukt produksjon og skogbruk har ført til varierte innsats for å finne og forutsi dormancy perioden3. Frukt treslag, empirisk eksperimenter overføre skudd å tvinge forhold og statistiske spådommer basert på data på blomstrende er gjeldende tilnærminger til å bestemme datoen for bryte uvirksom, som tillater forskere å anslå den Chilling krav for hver plante. Hvordan kontrollere dormancy basert på biologiske prosesser er imidlertid uklart3.
Blomstrende i tempererte frukttrær, som søte kirsebær (Prunus avium L.), oppstår en gang i året og varer fjorten dager. Men begynner blomster å skille og utvikle ca 10 måneder tidligere, under forrige sommer4. Blomst primordia slutte å vokse i løpet av høsten forbli sovende inne knoppene vinteren. I denne perioden må hver sorten samle en spesielt skremmende krav for riktig blomstrende4. Til tross for mangel på phenological endringer i knoppene vinteren, blomst primordia er fysiologisk aktive under dormancy, og akkumulering av chilling temperaturer er nylig tilknyttet dynamiske av stivelse opphopning eller redusere i cellene i eggstokken primordium, varer en ny adgang for dormancy besluttsomhet5. Men krever den lille størrelsen og plasseringen av eggstokken primordium en spesiell metode.
Stivelse er større lagringsplass karbohydrater i woody plante arter6. Dermed har endringene i stivelse vært knyttet til fysiologiske aktiviteten blomst vev, som trenger karbohydrater for å støtte deres utvikling7,8. Ulike viktige hendelser under reproduktive prosessen er også knyttet til variasjoner i stivelsen innhold i ulike floral strukturer, for eksempel anther meiose9, veksten av pollen rør gjennom stil eller ovule befruktning10. Histochemical teknikker at påvisning av stivelse i hver bestemt vev av blomst primordia under dormancy. Vanskeligheten imidlertid i kvantifisere at stivelse tillate etter mønsteret av opphopning/redusere over tid eller sammenligner stivelse innhold blant vev, kultivarer eller år. Dette skyldes den lille mengden vev tilgjengelig for analytiske teknikker11. Alternativt kan bildeanalyser knyttet til mikroskopi12 kvantifisering av stivelse i svært små utvalg anlegget vev13.
Tilnærminger kombinere mikroskopi og bildet har blitt brukt å kvantifisere innholdet i ulike komponenter i anlegget vev, som callose14, microtubes15, eller stivelse16, ved å måle størrelsen på området farget av bestemte flekker. For stivelse, det kan enkelt oppdages ved hjelp av kalium iodide-jod (jeg2KI) reaksjon17. Denne metoden er svært spesifikke; Jeg2KI setter tiden i laminær strukturen av stivelse korn og danner en mørk blå eller rødbrun farge, avhengig av stivelse18amylose innhold. Deler farget med jeg2KI flekken Vis tilstrekkelig kontrast mellom stivelse og bakgrunn vevet, slik at en utvetydig stivelse gjenkjenning og den påfølgende kvantifiseringen av bildet analyse system19. Selv om dette fargestoffet ikke er stoichiometric, er akkumulering av jod proporsjonale med lengden av stivelse molekyl, som kan variere svært17. Derfor kanskje størrelsen på stained området uttrykt som antall piksler gjenspeiler ikke nøyaktig innholdet i stivelse, siden høy forskjeller i stivelsen innhold finner mellom felt med farget områder av samme størrelse. Alternativt kan stivelsen innhold vurderes ved å måle den optiske densitet for farget granulater på svart-hvitt bilder fra mikroskopet, som det har blitt rapportert i ulike vev i aprikos8,13 , 19, avocado10,20og oliven21.
Her beskriver vi en metode som kombinerer eksperimentell bestemmelse av dormancy status med kvantifiseringen stivelse innhold i eggstokken primordium vev fra høsten til våren i søte kirsebær, tilbyr et nytt verktøy for forståelse og prediksjon uvirksom basert på studier av de biologiske mekanismene knyttet til dormancy.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. dormancy besluttsomhet og plante materiale samling
- Prøve blomsterknopper i feltet. Dormancy studier er langsiktige eksperimenter og krever voksen trær stort nok for å samle knopper og skyter hele vinteren uten at trærne utvikling i løpet av neste våren. Spesielle orchard ledelse kan være nødvendig avhengig av opplæring systemet; beskjæring kan dermed være mindre alvorlig enn til frukt produksjonsformål.
- Hver uke, fra begynnelsen av høsten før utbruddet av bud pause, samle og veier 10 blomsterknopper.
- Fastsette blomsterknopper ved å plassere dem i 10-mL røret med en cap og suge prøvene i en etappe, den stabiliserende løsning av etanol/eddiksyre (3:1) for minst 24 timer på 4 ° C. Deretter forkaste bindemiddel og legge til 75% etanol, sjenerøst å sikre at det dekker prøvene. Eksemplene kan være bevart i denne løsningen på 4 ° C før bruk.
Merk: Vakuum infiltrasjon kan brukes å fjerne luftbobler i knopp og forhindre dem fra flytende. Dette Letter gjennomtrenging av bindemiddel i vev, men kan skade strukturen i vev. Prøv å unngå det hvis ikke nødvendig.
- Smake skyter i feltet.
- Hver uke, fra begynnelsen av høsten før utbruddet av bud pause, ta tre skudd på 15-30 cm i lengde og 5 mm i diameter, som inneholder minst 10 blomsterknopper som hver. Plass dem på vann-gjennomvåt blomsterhandler skum i en vekst kammer på 22 ± 1 ° C med en 12 h lys fotoperiode.
- Etter 10 dager i vekst kammer, plukke og veier 10 blomsterknopper fra skuddene.
- Evaluere bud veksten og kontrollere dormancy. Samplingsperiode har justeres vilkårene for plasseringen. Orchard forhold (Zaragoza, Spania, 41 ° 44'30 "N, 0 ° 47'00" W og 220 moh), prøvetaking skudd ble gjennomført fra 30 November til slutten av februar eller begynnelsen av mars.
- Ukentlig evaluere responsen av blomsterknopper egnet vekst vilkårene for kammeret, fra begynnelsen av høsten før budburst om våren, ved å sammenligne vekten av 10 knopper plukket fra feltet.
- Hvis det er ingen forskjeller eller disse forskjellene er mindre enn 30%, vurdere at knoppene ikke har oppfylt sine skremmende krav og endodormant22. Hvis forskjellene er mer enn 30%, vurdere at knoppene har oppfylt sine skremmende krav og ecodormant22.
2. plante materiale forberedelse til stivelse kvantifisering
- Velg om seks fast knopper fra hver prøvetaking dato (se trinn 1.1). Avhengig av Sorter inneholder hver søte kirsebær blomst knopp opptil fem blomst primordia.
- Fjern eksterne bud skalaer og plassere bud på en urmaker glass med 75% etanol å forhindre det tørker ut.
- Dissekere bud og ekstra minst en blomst primordium per bud med hjelp av presisjon tang og en ophthalmologic skalpell under stereoskopisk mikroskop. Blomst primordium kan bevares i 10-mL røret med 75% etanol 4 ° c, eller fortsette umiddelbart til neste trinn.
- Tørke prøvene i en tertiær butyl alkohol serie.
- Erstatt 75% etanol løsningen med 10 mL 75% tertiær butyl alkohol (TBA) å dekke sjenerøst prøvene og ruge dem for 1,5 t. En Pasteur pipette kan være nyttig å forkaste løsningen.
- Gjenta trinn 2.2.1, legge TBA med økende konsentrasjoner (85%, 95% og 100% v/v, og 3 x med ren TBA) i en tørking ovn på 30 ° C med luft ekstraksjon, siden ren TBA crystalizes ved romtemperatur (< 20 ° C) og er svært ustabil og giftige. Hvis TBA crystalizes med prøven, vil det skade vev.
- Bygge inn prøvene i parafin18.
- Smelt parafinen ved å innføre parafin perler i tørking ovnen på 60 ° C med luft ekstraksjon gårsdagen. Parafin perler kan også smeltet på en kokeplate, men sikre at den flytende parafinen er på 60 ° C og ikke på en høyere temperatur, å unngå skade på vev.
- Erstatt TBA med en blanding av TBA og parafin olje (1:1) og ruge 24 h inne en tørking ovn på 60 ° C. Deretter erstatte blanding av TBA og parafinolje med ren smelte parafin og ruge minst 6 h inne en tørking ovn på 60 ° C. Gjenta det 2 x og Inkuber siste endringen for minst 4-6 d.
- Plasser hver prøve på en liten metall base mold over en varme overflate, innebygd i parafinvoks, og tempo innebygging kassetten. Plasserer den over en kald overflate og fjerne blokken når voksen er styrket.
- Delen og rehydrate preparater.
- Forberede Haupts lim: oppløse 1 g vanlig Knox gelatin i 100 mL destillert vann på 30 ° C; Legg deretter til 2 g fenol løs krystaller (C6H5OH) og 15 mL glyserol. Den Haupt lim fordelt på et glass lysbilde med pensel og legge en dråpe av en 1% formaldehyd løsning.
- Hver parafin blokk 10 µm i en rotatory mikrotomen og plasserer delene på av objektglass dekket med Haupts limet.
- Plass glass lysbilder med delene over en varme overflate på 35-40 ° C til tørr; deretter Flytt glass lysbilder til en objektglasstativet.
- Forbered den dewax og rehydrering løsninger. Plasser 200 mL Histoclear II i tre glass flekker retter, en med Histoclear II:ethanol (1:1, v/v), en etanol-serien (100%, 70%, 40%, v/v), og en siste destillert vann vask.
- Dewax avsnittene med tre vasker, hver 5 min, i Histoclear II og Histoclear II:ethanol (1:1, v/v). Plasser objektglasstativet inne glass flekker retter, sikre at løsningen dekker helt lysbildene, og deretter flytter objektglasstativet fra en løsning til neste.
- Rehydrate delene av følgende serien av 2-min vasker: en etanol-serien (100%, 70%, 40%, v/v) og en siste destillert vann vask. Til slutt, tørr glass lysbilder ved romtemperatur.
- Stain deler18.
- Forberede jeg2KI flekken: oppløse 2 g av kalium iodide (KI) og 0.2 g jod (jeg2) i 100 mL destillert vann. Påfør en dråpe frisk jeg2KI over section(s) i 5 minutter og deretter kaste overskytende av flekker ved å absorbere det med blotting papir. Raskt fortsette til neste trinn.
- Bruke en liten dråpe en syntetisk monterer medier, Plasser et lite cover glass på og presse hardt. Når du monterer medier tørker, observere under mikroskop lyse-feltet for en foreløpig vurdering av eggstokk stivelse.
Merk: Dette trinnet er ikke obligatorisk, men en bedre kontrast mellom stivelse og bakgrunnen er oppnådd. Hvis delen må brukes med andre flekker etter stivelse kvantifisering, Legg dekket glasset over dråpe flekken og kaste overflødig ved å absorbere det med blotting papir (se 2.5.1). Deretter stivelse kvantifisering, vaskes ut I2KI flekker med destillert vann og Legg forberedelsene over varme overflaten på 35-40 ° C til tørr.
3. kvantifisering av stivelsen innhold
- Kalibrere optisk betingelsene.
Merk: Søking nivåene brukes av bildet analyseringen for å oppdage farget stivelse er direkte avhengig av lysforholdene og forstørrelsen av mikroskopet; dermed fikse disse forholdene for alle forberedelsene evalueres. Tilpasse justeringen foreslått her tilgjengelig mikroskop og lys.- Justere blenderåpning membranen på 20 X forstørrelse og lysstyrkekontroll eller lysintensiteten.
- Kontroller at det er ingen filtre på filterholderen, og velg en lys-feltbetingelse i mikroskopet. Velg en passende forstørrelse (f.eks40 X for søte kirsebær eggstokk primordium).
- Kontrollere bilde oppkjøpet betingelsene. Justere kameraet med en farget forberedelse uten vev via bilde | Kjøp | Pre-view.
- Fastsette lysstyrken på 50%, gevinsten til 1,0 x og histogrammet indikatorene gamma verdi på 1.00 og kontrasten på 0 - 100, kø med grensene for lysstyrke distribusjon histogrammet.
- Aktivere funksjonen overeksponering/undereksponering og justere eksponeringstiden på grensen av overeksponering.
- Hvitbalanse funksjonen gjelde hele bildet viser alle nøytral farge fargekomponentene i bildet uten alle fargetone og skyggelegging korreksjon hele bildet å opprette en korrigert og homogen image.
- Kalibrere bildet analyse systemet for å få kontrollverdiene fra den grå nivået (0, svart, 255, hvit) av forskjellig optisk tetthet (OD) er oppnådd av verdiene for transmisjon (T).
- Få et bilde av en farget forberedelse uten vev, vurdert kontroll hvit og måle den grå nivået av den svarte / hvitt bilde via mål | Grå mål | Kalibrere grå | Referere til verdien = 0 | Mål | Kalibrere | OK. Dette samsvarer med en 100% transmisjon; Således, en optisk densitet 0, ifølge OD = 2 - logge T.
- Kjøpe et bilde av samme utarbeidelse med et 4N filter som reduserer mengden av lys 4 x, og måle den grå nivået av den svarte / hvitt bilde via mål | Grå mål | Kalibrere grå | Referere til verdien = 0,6 | Mål | Kalibrere | OK. Dette samsvarer med en 25% transmisjon; Således, en optisk tetthet på 0,6, ifølge OD = 2 - logge T.
- Få et bilde av samme preparat uten lys, som svart og måle den grå nivået av den svarte / hvitt bilde via mål | Grå mål | Kalibrere grå | Referere til verdien = 1 | Mål | Kalibrere | OK. Dette samsvarer med en 0% transmisjon; Således, en optisk densitet 1, ifølge OD = 2 - logge T.
- Oppdage stivelse.
- Få et fargebilde av feltet måle i TIFF-format med en oppløsning på minst 300 punkt per tomme (PPT).
- Opprette en binær image tilsvarende området farget. Angi tre farger grensene (verdier mellom 0 - 255 for hver) til binære bildet nøyaktig gjenspeiler farget stivelse granulater observert, via bilde | Oppdage | Velg grensene av rødt, blått og grønt | OK. Gjør gjentatte ganger visuelle sammenlikninger i ulike forberedelser og vev å tune opp siste oppdagelsen nivåer. Lagre og bruke disse nivåene for alle forberedelser.
- Kvantifisere stivelse. Konvertere den opprinnelige fargebilde til et svart-hvitt bilde med bildet analysesystem. Bruke binær image som en lagt maske på svart-hvitt bilde via bilde | Binær redigere. Måle summen av den optiske densitet for hver piksel under den maske via mål | Grå nivå | OK og vurdere denne verdien som det stjernen innholdet i feltet målt.
- Gjenta trinn 3.4.1 - 3.4.3 av fire steder av eggstokk primordia å få representativt verdien stivelsen innhold i eggstokken av blomst primordia.
- Gjenta trinn 3.4.1 - 3.4.3 og trinn 3.4.5 i ulike blomster på hver samle dato.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Dormancy studier krever fastsettelse av øyeblikket når skremmende kravene er oppfylt. Til tross for mangel på phenological endringer vinteren under feltforhold (figur 1A) gjenopprette kirsebærtrær ikke kapasiteten til vekst i riktig forhold til de passerer en viss under lave temperaturer. Vanlig overføring av skudd til en kontrollerte forhold kammer (figur 1B) om vinteren lov Evaluering av dormancy status blomsterknopper. Evalueringen av blomst knopp veksten ble gjort ved å måle økningen i knopp vekt. Mens under dormancy, ingen endringer kan være observert etter 10 dager med riktig forhold (figur 1 c), når dormancy ble overveldet, knoppene svulmet og brast i vekst kammer (figur 1 d). Resultatene av denne analysen tillatt dormancy status av knopper skal etableres. På grunn av de forskjellige temperaturene skjer om vinteren, var dormancy overveldet på ulike tidspunkter, avhengig av året. Mens det første året av studien, bryte uvirksom skjedde i januar, andre år presentert en mildere vinter; Dermed oppstod skremmende oppfyllelsen ca tre uker etter, i februar.
Søte kirsebær Bjørn blomsterknopper i spurs, der apikale bud er en vegetativ bud og lateral knopper er blomsterknopper (figur 1 c og 1 D). Udifferensierte knopper begynte å skille ut blomst eller vegetative knopper på slutten av sommeren, og når de skriver inn dormancy vinteren, flere blomst primordia igjen inne knoppen, beskyttet av tallrike grønne skjell og dekket av ytre vekter ( Figur 1 c og figur 2A). Disseksjon av blomst knopp viste liten blomst primordia inne (figur 2A). Hver blomst knopp inneholdt ett til fem personlige blomst primordia (figur 2B). Til tross for den lille størrelsen på hver blomst primordium, alle deler av en blomst er differensiert og kan skilles: pistil, anthers, kronbladene og kronblader (figur 2C). Bruk av histochemical teknikker (alkohol: eddiksyre [3:1] fiksering, parafinvoks innebygging, mikrotomen snitting og jod-baserte stivelse flekker) lov distribusjon av stivelse i blomsten primordium vev å bli observert (figur 2D ).
Stivelse i eggstokken primordium ble kvantifisert i hver del. Fire tiltak på 1337 µm2, 40 x forstørrelsen, vises det generelle oppsettet stivelse søte kirsebær eggstokk primordium (figur 3A). Stivelse granulater var klart adskilte fra bakgrunnen etter jeg2KI flekker (figur 3B). Stivelse ble identifisert av analysesystem bildet ved å justere fargen terskler av rødt, grønt og blått til alle stivelse granulater observert var dekket av binære bildet opprettet av systemet basert på parameterne definert farge (Figur 3 c). Verdiene for stivelse innhold fra var resultatet av mål på den optiske densitet for hver piksel under masken på svarte og hvite bildet (figur 3D).
Kvantifisering av stivelse avslørte en konsistent mønster av stivelse dynamisk vinteren (Figur 4). Konsekvent, presentert mengden av stivelse ut tidlig på vinteren en optisk densitet verdi på mindre enn 40.0003, mens maksimale nådde en verdi mellom 120 000 og 140 000 i både år. Mens maksimumsverdien er nådd i januar første året (figur 4A), skjedde i februar i det andre året (figur 4B). Kontrast disse resultatene med hudfarge uvirksom, oppstod maksimale stivelse samtidig med skremmende oppfyllelsen i både år.
Denne fremgangsmåten krever fastsettelse av dormancy status (figur 5A) samtidig med stivelse kvantifiseringen på eggstokken vev (figur 5B) for å ramme endringene i det stjernen innholdet i forhold til dormancy.

Figur 1: eksperimentelle set-up for fastsettelse av dormancy status blomsterknopper av søte kirsebær. (A) grener, om vinteren Vis sovende knoppene lukket og dekket av mørkebrune vekter. (B) dette panelet viser skudd overført til vekst kammer. I midten av enkelte Sorter forble sovende med knopper fortsatt stengt (hvit pil), mens andre kunne vokse, viser svulmet knopper (svart pil). (C) dette panelet viser en detalj av et skudd med sovende blomsterknopper ligger lateralt og en enkelt vegetative bud på apikale beliggenhet i anspore. (D) dette panelet viser en detalj av et skudd når dormancy var overveldet etter 10 dager i voksende kammeret, viser bud hevelse. Klikk her for å se en større versjon av dette tallet.
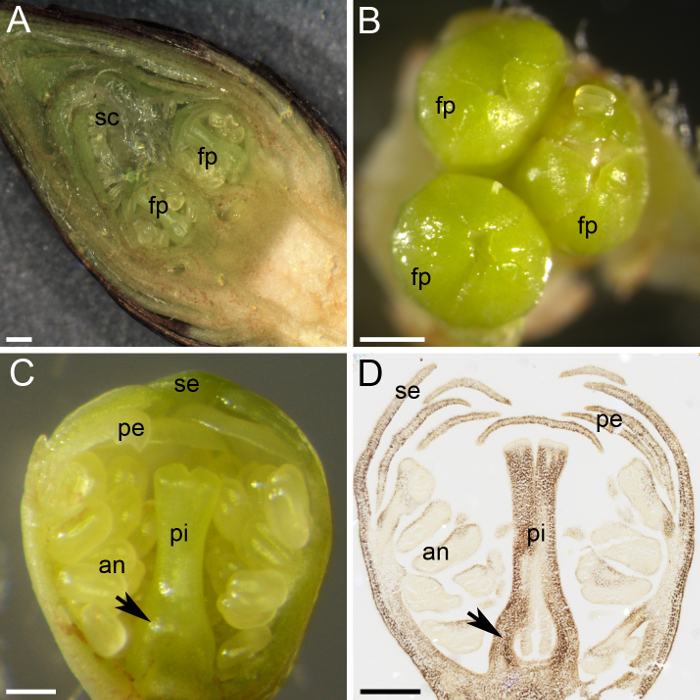
Figur 2: plante materiale forberedelse til stivelse kvantifisering. (A) dette panelet viser en tverrgående del av en blomst knopp, viser to blomst primordia (fp) beskyttet av tallrike skjell (sc). (B) tre blomst primordia (fp) samles i en blomst knopp. (C) dette panelet viser en tverrgående del av en blomst primordium, med alle kranser differensiert: kronblader (se), kronblad (pe), anthers (en) og pistil (pi). Eggstokken primordium kjennetegnes ved foten av pistil (pil). (D) en midten av en blomst primordium var samlet og løst i januar, innebygd i parafinvoks, delt og farget med jeg2KI (mørk brun) for stivelse. Dette panelet viser eggstokk primordium (pil). Skala barer er 500 µm i paneler A og B og 200 μm i paneler C og D. Klikk her for å se en større versjon av dette tallet.

Figur 3: stivelse kvantifisering i søte kirsebær eggstokk primordium. (A) dette panelet viser en midten av en eggstokk primordium med jeg2KI, viser fire rammene i hvilke stivelse innhold ble målt. (B) dette panelet viser en detalj av eggstokk-primordium. Stivelse granulater er farget i mørk brun. (C) dette panelet viser et bilde med pseudofarger i hvilke stivelse tilsvarer ulike nyanser av blått. (D) dette panelet viser en binære bilder maske dekker jeg2KI-farget stivelse (blå) på svart-hvitt originalbildet. Optisk densitet måles i piksler på det opprinnelige bildet dekket av masken. Skala barer er 100 µm i panelet A og 20 µm i paneler B - D. Klikk her for å se en større versjon av dette tallet.

Figur 4: representant resultatene av stivelse kvantifiseringen i søte kirsebær eggstokk primordia samlet månedlig fra høsten til våren i løpet av to år med forskjellige vinter temperaturer. (A) dette panelet viser resultatene fra år 2010-2011, som hadde en kald vinter. Skremmende oppfyllelsen (snøfnugg) oppstod i januar, samtidig med maksimale stivelse. (B) dette panelet viser resultatene fra år 2011-2012, som hadde milde vintre. Skremmende oppfyllelsen (snøfnugg) oppstod i februar, samtidig med maksimale stivelse. Verdiene er de gjennomsnittlig ± standardfeilen til gjennomsnittet. Klikk her for å se en større versjon av dette tallet.
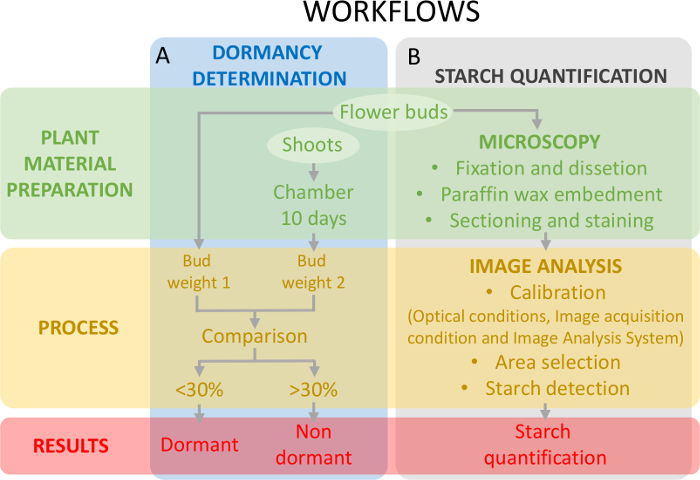
Figur 5: ordningen eksperimentell design å vurdere dormancy status knopper og stivelse kvantifiseringen i eggstokken primordia i søte kirsebær. (A) dette panelet viser arbeidsflytene dormancy status besluttsomhet: plante materiale utarbeidelsen prosessen og resultatene. (B) dette panelet viser arbeidsflytene for stivelse kvantifisering: histochemical forberedelse av knopper til en mikroskopisk observasjon for stivelse, bildet analyse påvisning av stivelse og stivelse kvantifisering. Klikk her for å se en større versjon av dette tallet.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Dormancy i woody stauder presenterer klare konsekvenser i frukt produksjon og Skogsdrift i et klima i endring, selv om biologiske prosessen bak dormancy uklar. Dormancy studier kan være nærmet fra ulike synsvinkler, men forskningen på jakt etter en biologisk markør for Vinter dormancy har intensivert de siste årene. Men har de fleste forsøk på å finne en utvetydig indikator viser når en har brutt dormancy vært mislykket3. Metodikk beskrevet heri, kombinere histochemical teknikker18 med analyse bilde, har vært veldig nyttig å undersøke forholdet mellom karbohydrater reserver av et bestemt vev og dens fysiologisk aktivitet i søte kirsebær knopper i forskjellige faser av dormancy5 og kan brukes også på andre arter og vev10,13,20.
Karbohydrater reserver i form av stivelse spiller en viktig rolle både flower utvikling og reproduktive prosessen7,10,13,20,21,23 og i sesongvariasjoner av temperert woody stauder6. Forskjellige studier på dormancy har betalt oppmerksomhet til stivelse i knopper24. Men på grunn av sin lille størrelse, flere hele knopper kreves for bruk av kvantitative analytiske metoder, og oppdagelsen av kvantitative variasjoner i bestemt vev eller celler er begrenset av maskering effekten av de omkringliggende cellene. Kombinasjonen av histochemical teknikker med bildet analyse systemet tilbyr en god mulighet til å studere endringer i ulike strukturer inne knoppen stivelsen innhold.
Denne metoden har begrensning for å hindre kvantifisering av det eksakte innholdet i stivelse i vevet men kan relative stivelsen innhold til kvantifiseres å følge kvantitative stivelse endringer over tid25 og sammenligne stivelse av forskjellige vev13,20, kultivarer eller år5. For at riktig sammenligning av verdiene for optisk tetthet mellom felt, vev og knopper, kalibrering av systemet (lys flekker intensitet og forstørrelse) og innstillingen for farge terskler må etableres nøyaktig , lagres og brukes for alle forberedelser.
Stivelse flekker og kvantifisering basert på kalium jod kan påfølgende bruk av annen flekker etter vask av delen. Dermed ulike analyser kan utføres uten ekstra forberedelser og syntetisk montering media ikke er brukt5,20. Likeledes kan morphometric målinger26 gjøres etter samme forberedelsene, slik at mønsteret av stivelse akkumulering til innramming i forhold til veksten av ulike strukturer5,13. Metoden kan tilpasses andre strukturer eller arter med justering av farge nivåene som bruker analyzer til å gjenkjenne stivelse, noe som gjør studiet av andre utviklingsprosesser som innebærer endringer i stivelsen innhold i små grupper av celler.
Forholdet mellom dormancy utgivelsen og stivelse akkumulering i eggstokken primordia avduket ved bruk av denne metoden gir et godt grunnlag for å forstå det biologiske grunnlaget for dormancy og skremmende krav5. Imidlertid kan stivelse kvantifisering av bildeanalyse på parafin-embedded deler vise seg for å være svært upraktisk og tidkrevende å anslå skremmende kravene til mange kultivarer. Arbeidet må være fokusert på å studere pålitelig biologiske indikatorer som kan enkelt angi statusen dormancy av treet. I mellomtiden mønster av stivelse-varianter under dormancy kan brukes til å ramme videre fysiologiske og genetiske studier, og kombinasjonen av histochemical teknikker med bildeanalyser beskrevet her kan brukes i andre woody flerårig avlinger til kvantifisere stivelsen innhold av ulike vev i forhold til dormancy.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Forfatterne ikke avsløre.
Acknowledgments
Forfatterne takke takknemlig Maria Herrero og Eliseo Rivas for nyttig diskusjon og råd. Dette arbeidet ble støttet av Ministerio de Economía y Competitividad-europeisk Regional Development Fund, EU [bevilgning nummer BES-2010-037992 til E. F.]; Instituto Nacional de Investigación y Tecnología Agraria y Alimentaria [grant tall RFP2015-00015-00, RTA2014-00085-00, RTA2017-00003-00]. og Gobierno de Aragón-European Social Fund, EU [Grupo Consolidado A12-17R].
Materials
| Name | Company | Catalog Number | Comments |
| Precision scale | Sartorius | CP225D | |
| Stereoscopic microscope | Leica Microsystems | MZ-16 | |
| Drying-stove | Memmert | U15 | |
| Paraffin Embedding station | Leica Microsystems | EG1140H | |
| Rotatory microtome | Reichert-Jung | 1130/Biocut | |
| Microtome blade | Feather | S35 | Stainless steel |
| Bright field microscope | Leica Microsystems | DM2500 | |
| Digital Camera | Leica Microsystems | DC-300 | |
| Image Analysis System | Leica Microsystems | Quantiment Q550 |
References
- Kurokura, T., Mimida, N., Battey, N. H., Hytönen, T. The regulation of seasonal flowering in the Rosaceae. Journal of Experimental Botany. 64 (14), 4131-4141 (2013).
- Rohde, A., Bhalerao, R. P.
- Fadón, E., Rodrigo, J. Unveiling winter dormancy through empirical experiments. Environmental and Experimental Botany. 152, 28-36 (2018).
- Fadón, E., Rodrigo, J., Herrero, M. Is there a specific stage to rest? Morphological changes in flower primordia in relation to endodormancy in sweet cherry (Prunus avium L.). Trees - Structure and Function. , In Press (2018).
- Fadón, E., Herrero, M., Rodrigo, J. Dormant flower buds actively accumulate starch over winter in sweet cherry. Frontiers in Plant Science. 9 (171), (2018).
- Loescher, W. H., Mccamant, T., Keller, J. D. Carbohydrate reserves, translocation and storage in woody plant roots. HortScience. 25 (3), 274-281 (1990).
- Hedhly, A., et al. Starch turnover and metabolism during flower and early embryo development. Plant Physiology. , (2016).
- Rodrigo, J., Hormaza, J. I., Herrero, M. Ovary starch reserves and flower development in apricot (Prunus armeniaca). Physiologia Plantarum. 108 (1), 35-41 (2000).
- Julian, C., Rodrigo, J., Herrero, M. Stamen development and winter dormancy in apricot (Prunus armeniaca). Annals of Botany. 108 (4), 617-625 (2011).
- Alcaraz, M. L., Hormaza, J. I., Rodrigo, J. Pistil starch reserves at anthesis correlate with final flower fate in avocado (Persea americana). PLoS One. 8 (10), e78467 (2013).
- Smith, A. M., Zeeman, S. C. Quantification of starch in plant tissues. Nature Protocols. 1 (3), 1342-1345 (2006).
- Eliceiri, K. W., et al.
- Rodrigo, J., Herrero, M. Influence of intraovular reserves on ovule fate in apricot (Prunus armeniaca L.). Sexual Plant Reproduction. 11, 86-93 (1998).
- Zhou, J., Spallek, T., Faulkner, C., Robatzek, S. CalloseMeasurer: A novel software solution to measure callose deposition and recognise spreading callose patterns. Plant Methods. 8 (1), (2012).
- Faulkner, C., et al. An automated quantitative image analysis tool for the identification of microtubule patterns in plants. Traffic. 18 (10), 683-693 (2017).
- Kuhn, B. F. Determination of starch in ovules of the sour cherry cv. "Stevnsbaer.". European Journal of Horticultural Science. 71 (3), 120-124 (2006).
- Johansen, D. A. Plant microtechnique. , McGraw-Hill. New York, NY. (1940).
- Ruzin, S. E. Plant microtechnique and microscopy. , Oxford University Press. New York, NY. (1999).
- Rodrigo, J., Rivas, E., Herrero, M. Starch determination in plant tissues using a computerized image analysis system. Physiologia Plantarum. 99 (1), 105-110 (1997).
- Alcaraz, M. L., Hormaza, J. I., Rodrigo, J. Ovary starch reserves and pistil development in avocado (Persea americana). Physiologia Plantarum. 140 (4), 395-404 (2010).
- Suarez, C., Castro, A. J., Rapoport, H. F., Rodriguez-García, M. I. Morphological, histological and ultrastructural changes in the olive pistil during flowering. Sexual Plant Reproduction. 25, 133-146 (2012).
- Lang, G. A., Early, J. D., Martin, G. C., Darnell, R. L. Endodormancy, paradormancy, and ecodormancy - Physiological terminology and classification for dormancy research. HortScience. 22 (3), 371-377 (1987).
- Hedhly, A., Vogler, H., Eichenberger, C., Grossniklaus, U. Whole-mount clearing and staining of arabidopsis flower organs and siliques. Journal of Visualized Experiments. 2018 (134), 1-10 (2018).
- Kaufmann, H., Blanke, M. Changes in carbohydrate levels and relative water content (RWC) to distinguish dormancy phases in sweet cherry. Journal of Plant Physiology. 218 (July), 1-5 (2017).
- Herrero, M., Dickinson, H. G. Pollen-pistil incompatibility in Petunia hybrida: changes in the pistil following compatible and incompatible intraspecific crosses. Journal of Cell Science. 36, 1-18 (1979).
- Carpenter, A. E., et al. CellProfiler: image analysis software for identifying and quantifying cell phenotypes. Genome Biology. 7 (10), R100 (2006).
