

Summary
Resirkulering endosomer er en del av endosomal rørformet nettverk. Her presenterer vi en metode for å kvantifisere dynamikken i resirkulering endosomer ved hjelp av GFP-STX13 som organellemarkør.
Abstract
Resirkulering endosomer (REs) er rørformede-vesikulære organeller generert fra tidlig / sortering endosomer i alle celletyper. Disse organellene spiller en nøkkelrolle i biogenesen av melanosomer, en lysosomerelatert organell produsert av melanocytter. REs leverer melanocyttspesifikk last til for tidlige melanosomer under dannelsen. Blokkering i generering av REs, observert i flere mutanter av Hermansky-Pudlak syndrom, resulterer i hypopigmentering av hud, hår og øye. Derfor er det nyttig å studere dynamikken (refererer til antall og lengde) av REs for å forstå funksjonen til disse organellene i normale og sykdomstilstander. I denne studien tar vi sikte på å måle RE-dynamikken ved hjelp av en resident SNARE STX13.
Introduction
Biosyntese av melaninpigmenter forekommer i melanosomer, en melanocyttspesifikk lysosomerelatert organelle (LRO) som eksisterer sammen med konvensjonelle lysosomer. Det endokytiske systemet spiller en nøkkelrolle i biogenesen av melanosomer, som kreves for hudfarge og fotobeskyttelse mot ioniserende stråling1,2,3. I løpet av denne prosessen sorteres melaninsyntetiserende enzymer på tidlig / sortering av endosomer og transporteres deretter til for tidlige melanosomer gjennom rørformede eller vesikulære endosomer kalt resirkulering endosomes (REs) 4,5,6,7,8,9,10. Målrettingen og sammensmeltingen av disse organellene regulerer modningen av fullt funksjonelle pigmenterte melanosomer7,11,12,13,14. Defekter i dannelsen av disse organellene eller lastsortering til disse organellene forårsaker oculocutaneous albinisme og andre kliniske fenotyper, observert i Hermansky-Pudlak syndrom15,16.
Her beskriver vi en enkel mikroskopibasert teknikk for å studere og analysere RIT-ene. I denne metoden har vi benyttet oss av et transmembranprotein, Qa-SNARE Syntaxin (STX)13 som ligger på resirkulering endosomes17 og sykluser mellom sortering av endosomer og melanosomer i melanocytter12,18. Videre tillater sletting av N-terminal ustrukturert regulatorisk domene (nemlig SynN eller STX13Δ129) SNARE å sitte fast i melanosomer, som måler fremoverhandelsveien mot melanosome12. Vi har brukt en kjent gjenvinning endosomal markør Rab GTPase (Rab)11 i våre studier14,19. Fluorescensavbildning av proteinene GFP-STX13WT, GFP-STX13Δ129, mCherry-Rab11 og TYRP1 i ville melanocytter etterfulgt av kvantifisering av deres relative lokalisering vil gi naturen og dynamikken til REer i tillegg til deres målretting mot melanosomer. Dermed er dette en enkel teknikk som kan brukes til å visualisere og måle dynamikken i RE i melanocytter.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
Protokollen innebærer såing av melanocytter etterfulgt av transfeksjon av plasmidene. Ytterligere trinn inkluderer fiksering, immunoppnåelse, avbildning og analyse av cellene for å måle lengde og antall REer. Den detaljerte beskrivelsen av protokollen er gitt nedenfor.
1. Såing av mus melanocytter på forhåndsbehandlede deksler
- Belegge glassdekslene i en Petri-tallerken (dvs. 4 - 5 i en 35 mm tallerken) med kjellermembranmatrisemedium (1:20 i komplett RPMI-medium: RPMI + 10% varmeinaktivert FBS + 1x Glutamin + 1x antibiotikablanding) og tørk den i vevskulturhetten i 15 min. Vask dekslene én gang med 1x PBS før bruk.
- Vedlikehold musen melanocytter av vill type (melan-Ink4a-Arf-1 fra C57BL/6J, a/a, Ink4a-Arf-/- mus, beskrevet i20 og tilgjengelig på The Welcome Trust Functional Genomics Cell Bank) i en Petri-tallerken (dvs. 35 eller 60 mm parabolen) supplert med komplett RPMI-medium.
- Vask cellene to ganger med 1x PBS og tilsett 0,5 eller 1 ml Trypsin-EDTA (0,25%) løsning for trypsinisering av celler. Inkuber cellene ved 37 °C i 2 - 5 min for løsrivelse fra Petri-retten.
- Tilsett 1 - 2 ml komplett RPMI-medium, suspender og overfør deretter cellene til et sentrifugerør.
- Sentrifuger cellefjæringen ved 4 °C, 376 x g i 5 minutter og bruk deretter pelletsen på nytt i 1x PBS.
- Gjenta sentrifugeringstrinnet og resuspender cellene i 1 ml komplett RPMI-medium.
- Frø cellene på kjellermembranen middels belagte deksler med 50-60% samløp (ca. 6 x 105 celler på en 35 mm cellekulturfat som inneholder 4 - 5 deksler). Tilsett alltid 200 nM PMA (tilsett alltid 5 μL arbeidslager 40 μM phorbol 12-myristate 13-acetat) til den belagte cellefjæringen i komplett RPMI-medium.
- Etter sådd, inkuber platen ved 37 °C i 12-24 timer.
2. Transfeksjon av celler med STX13 plasmider
- Bruk følgende reagenser: pEGFP-C1-STX13WT og pEGFP-C1-STX13Δ129 (beskrevet i12). mCherry-Rab11, var en vennlig gave fra Graça Raposo, Institut Curie, Paris (beskrevet i referanse19). Anti-TYRP1 antistoff fra ATCC (TA99).
- Etter 12 - 24 timers såing, transfekt cellene med plasmider ved hjelp av et lipidbasert transfeksjonsreagens. For en 35 mm tallerken, ta 5 μL av transfeksjonsreagenset i 250 μL OPTI-MEM medium i et mikrocentrifugerør og ta omtrent 200 ng av hver plasmid i 250 μL OPTI-MEM.
- Inkuber rørene som inneholder DNA og transfeksjonsreagenset i 5 min. Bland uten gjentatt pipettering (totalt volum vil være ca. 500 μL). Inkuber i 30 min ved romtemperatur. Utfør håndsapping av røret hvert 10.
- Under inkubasjonen, vask cellene to ganger med 1x PBS, en gang med OPTI-MEM og tilsett deretter 1 ml OPTI-MEM i cellene.
- Legg til transfeksjonsreagens-DNA-blandingen til cellene på en dråpevis måte ved å dekke parabolen.
- Inkuber cellene ved 37 °C i 6 timer. Aspirer OPTI-MEM-mediet med transfeksjonsreagens og tilsett komplett RPMI-medium supplert med 200 nM PMA.
- Inkuber cellene ved 37 °C i 48 timer.
3. Fiksering av cellene
MERK: Følgende prosedyre utføres utenfor vevskulturhetten.
- Etter 48 h transfeksjon, vask cellene to ganger med 1x PBS og fest deretter cellene med 3% formaldehyd (fersk tilberedt i 1x PBS) i 30 minutter.
- Etter fiksering, vask cellene to ganger med 1x PBS og oppbevar dekslene i 1x PBS til videre bruk. Alternativt kan celler monteres på glasssklier (se nedenfor) eller lagres ved 4 °C.
4. Immunstaining av cellene
- Forbered et fuktig kammer: Legg parafinfilmskåret stykke på fuktig filterpapir i en Petri-tallerken, dekket med aluminiumsfolie.
- Forbered 25 μL primær antistoffløsning (0,2% saponin i 1x PBS, 0,1% BSA i 1x PBS og 0,02% natriumazid i 1x PBS). Tilsett antistoff ved en fortynning på 1:200. Tilsett denne løsningen som en dråpe på en parafinfilm i det fuktige kammeret.
- Løft forsiktig dekslene med tang, snu det på denne dråpen av primær antistofffargingsløsning og dekk deretter lokket på det fuktige kammeret. Inkuber ved romtemperatur i 30 min.
- På samme måte, lag den sekundære antistoffløsningen ved en fortynning på 1:500 og legg den på parafinfilm ved siden av dekslene i det fuktige kammeret. For farging av kjernen, tilsett DAPI (1:20,000 til 1:30,000) til løsningen.
- Bruk tang, plukk forsiktig opp dekslene fra den primære antistoffløsningen og dypp den tre ganger i 1x PBS (i et glassbeger).
- Trykk på dekslene på vevspapir for å fjerne overflødig PBS på dekslene. Plasser den på sekundær antistofffargingsløsning i det fuktige kammeret og ikke utsett den for lys på grunn av tilstedeværelsen av fluorescerende merkede antistoffer i løsningen.
- Inkuber dekslene igjen i 30 min ved romtemperatur. Vær alltid oppmerksom på siden av dekslene som har cellene gjennom disse trinnene.
- Etter inkubasjonen, plukk forsiktig opp dekslene fra den sekundære antistoffløsningen og dypp den tre ganger i 1x PBS. Trykk videre på dekslene på vevspapir for å fjerne overflødig PBS på dekslene.
- Plasser 12 μL Fluoromount-G monteringsreagens på en glasssklie og plasser forsiktig den fargede dekslen (vendt mot glasset) på monteringsreagenset. Snu glassskuffen på vevspapir, og trykk deretter forsiktig.
5. Fluorescensmikroskopi av cellene
- Bilde de fargede cellene under Bright-field (BF) og fluorescens (IF) filtre ved hjelp av et invertert fluorescensmikroskop utstyrt med et CCD-kamera ved hjelp av 60x (olje) apochromatisk mål eller et annet mikroskop med en lignende konfigurasjon.
6. Kvantifisering av overlapping mellom RE lokaliserte proteiner og melanosomer:
MERK: Følgende trinn følges for kvantifisering av Manders overlappingskoeffisient mellom proteinene ved hjelp av Fiji-programvare (fritt nedlastbar fra lenken: https://imagej.net/software/fiji/). Bruk TIFF-bildet med flere kanaler.
- Åpne raw-bildet. Gå til Bilde-alternativet, velg Farge | Del kanaler, og bruk de to kanalene til analyse.
- Åpne JACoP-plugin-modulen i Plugin-alternativet.
- Angi terskelen for begge kanalene slik at alle lyspunktene er valgt og bakgrunnen elimineres.
- Gå til Analyse-alternativet, velg M1- og M2-koeffisienter for å få Manders overlappingskoeffisient.
- Trykk på Analyser-alternativet i JACoP-plugin-modulen, og se resultatet som viser Manders overlappingskoeffisient.
7. Kvantifisering av resirkulering endosomer rørformet antall og lengde:
MERK: Følgende trinn følges for kvantifisering av antall og lengde på tubuliene ved hjelp av Fiji-programvare.
- Åpne alternativet Raw image Gå til bilde, velg Farge | Del kanaler og bruk ønsket kanal for analyse.
- Gå til Bilde på nytt, velg Type | Konverter til 8-biters bilde.
- Gå deretter til Plugins, velg Analyser | Rørhet. Angi sigmaverdi til 0,1075 . Trykk OK.
- Gå til Bilde på nytt, velg Type | Konverter til 8-biters bilde.
- Gå til bilde | Juster | Terskelverdi (bruk de samme terskelverdiene for alle bildene. Bilder bør ha omtrent lik intensitet).
- Gå til Prosess | Binær | Konverter til maske.
- Gå til Prosess, velg Binær , og velg deretter Skeletonize.
- Gå til Analyser, velg Skjelett og velg Analyser skjelett. Resultat og utdata | velger (a) Beregn den høyeste korteste banen | (b) viser detaljert informasjon | (c) viser merket skjelett. Trykk OK.
MERK: Resultatet åpnes i tabellformat. Kolonnen med gjennomsnittlig grenlengde viser lengden på alle de forskjellige tubuliene i cellen som er valgt (angi skala for å hente verdier i mikrometer). - Hvis du vil ha tak i antall tubuli i cellen, går du til Analyser og velger Analyser partikkelalternativ . Trykk OK.
MERK: I de oppnådde resultatene viser tellekolonnen antall tubuli i den aktuelle cellen. - Lagre dataene og analyser.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Kvantifisering av STX13Δ129 mutant lokalisering til melanosomene
Immunfluorescensmikroskopi av STX13 i mus wild type melanocytter viste GFP-STX13WT lokalisert som vesikulære og rørformede strukturer og GFP-STX13Δ129 lokalisert som ringlignende strukturer i tillegg til celleoverflaten (figur 1A). Videre viste intracellulær ringlignende GFP-STX13Δ129 kolokalisering med melanosomproteinet TYRP1 (figur 1A) og lysfeltbildede melanosomer (data ikke vist)12. Som vist tidligere, observeres en kohort av overekspressert GFP-STX13WT i melanosomer12. For å måle den relative lokaliseringen av GFP-STX13WT og GFP-STX13Δ129 til melanosomer, har vi brukt Fiji og analysert med JACoP-plugin. Den målte manderens overlappingskoeffisient (MOC) mellom GFP-STX13Δ129 med TYRP1 er ca. 1,5 ganger høyere sammenlignet med GFP-STX13WT med TYRP1 (figur 1B). Interessant nok viste TYRP1 2,9 ganger høyere MOC-verdier med GFP-STX13Δ129 sammenlignet med GFP-STX13WT (figur 1B). Disse dataene indikerer at lokaliseringen av GFP-STX13Δ129 til melanosomer er relativt høyere sammenlignet med GFP-STX13WT i steady state.
Kvantifisering av STX13WT lokalisering til resirkulering endosomes
Immunfluorescensmikroskopi av GFP-STX13WT viste kolokalisering med kjent resirkulering endosomalt protein Rab11 (uttrykt som mCherry-Rab11) (figur 2A, B). Den målte MOC mellom GFP-STX13WT og mCherry-Rab11 er ca. 1,4 ganger høyere sammenlignet med mCherry-Rab11 med GFP-STX13WT (figur 2B). For å måle antall og lengde på GFP-STX13WT-positive endosomale tubuli, har vi brukt Fiji-programvare som beskrevet i protokolldelen. mCherry-Rab11 brukes som en positiv kontroll i forsøkene (figur 2). Melanocytter transfektert med GFP-STX13WT viste et høyere antall tubuli per celle sammenlignet med celler som uttrykker mCherry-Rab11 (Figur 2C toppgraf, sammenlign bar A med bar B). Tubule-tallene reduseres imidlertid ved samuttrykk av GFP-STX13WT og mCherry-Rab11 i cellene (figur 2C toppgraf, sammenligne bar A med C og bar B med D). Interessant nok er den gjennomsnittlige tubule lengden (μm) for både GFP-STX13WT og mCherry-Rab11 sammenlignbar med hverandre i celler som uttrykker individuelt eller sammen (Figur 2C bunngraf). Sammen antyder disse dataene at GFP-STX13WT lokaliserer til REer som ligner på Rab11.
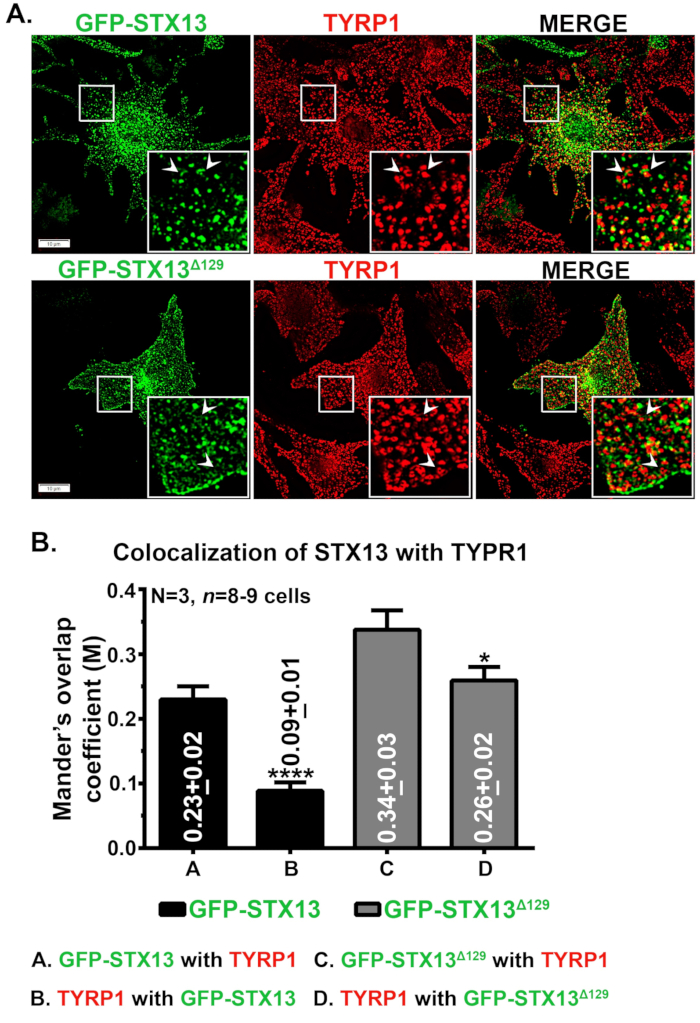
Figur 1: Lokalisering av GFP-STX13WT og GFP-STX13Δ129 til melanosomer i villtype melanocytter. (A) Melan-Ink4a-Arf-1 melanocytter ble transfected med GFP-STX13WT og GFP-STX13Δ129. Celler ble fikset, farget med anti-TYRP1 antistoff og deretter analysert ved fluorescensmikroskopi. Innfelt er forstørret visning av de hvite innrammede områdene. Piler peker på lokalisering av GFP-STX13WTto REs og GFP-STX13Δ129 til melanosomer. Skalastenger, 10 μm. (B) Kvantifisering av colokalisering mellom STX13 og TYRP1. Manders overlappingskoeffisient (M) mellom GFP-STX13WT eller GFP-STX13Δ129 med TYRP1 og omvendt er representert (gjennomsnittlig ± S.E.M.) separat i plottet. N=3. *p ≤0,05 og ****p ≤0.0001. Klikk her for å se en større versjon av denne figuren.

Figur 2: Lokalisering av GFP-STX13WT til resirkulering av endosomer i ville melanocytter. (A) Melan-Ink4a-Arf-1 melanocytter ble transfektert med GFP-STX13WT og mCherry-Rab11. Celler ble fikset og analysert ved fluorescensmikroskopi. Innfelt er forstørret visning av de hvite innrammede områdene. Piler peker på lokaliseringen av GFP-STX13WT til mCherry-Rab11-positive rom. Skalastenger, 10 μm. (B) Kvantifisering av colokalisering mellom STX13 og Rab11. Manders overlappingskoeffisient (M) mellom GFP-STX13WT med mCherry-Rab11 og omvendt er representert (gjennomsnittlig ± S.E.M.) separat i plottet. N=3. p ≤0.001. C. Kvantifisering av antall og lengde (i μm) av STX13- eller Rab11-positive REer. Gjennomsnittlig antall tubuli/celle og gjennomsnittlig tubule lengde på GFP-STX13WT og mCherry-Rab11 er representert (gjennomsnittlig ± S.E.M.) separat i plottet. N=3. Merk at cellene er transfektert med GFP-STX13WT og mCherry-Rab11 i (C) og (D). Klikk her for å se en større versjon av denne figuren.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Resirkulering av endosomer er en kohort av endokytiske organeller, og de formidler resirkulering av last til celleoverflaten i alle celletyper21,22,23,24,25. I spesialiserte celletyper som melanocytter, avleder disse organellene delvis sin trafficking rute mot melanosomene for deres biogenese3,16,26. Videre genereres REer etter lastsortering på tidlig / sortering endosomer og morfologisk vises som rørformede vesikulære strukturer. Deres dynamikk (antall og lengde) er avhengig av flere cellulære faktorer, inkludert motorproteiner21,22,23,24,25,27,28. Få studier brukte Rab11 som markør for å merke RE-ene i melanocytter14,19. Mens i ikke-melanocytter har STX13 og KIF13A blitt brukt i tillegg til Rab11 for å visualisere og karakterisere REs13,17,19. I denne studien har vi brukt STX13 som markør for å merke RIT-ene i melanocytter etterfulgt av en sammenligning med Rab11-positive tubules (figur 2). Interessant nok resulterer N-terminal sletting i STX13 (GFP-STX13Δ129) i feillokalisering av SNARE til melanosomer (figur 1). Dermed visualiserer lokaliseringen av GFP-STX13WT og GFP-STX13Δ129 i melanocytter henholdsvis RE og melanosomer. Disse proteinene kan brukes som respektive organeller markører for steady-state lokaliseringsstudier.
Studier har vist at flere proteiner som TYRP1, TYR (tyrosinase), VAMP7, OCA2, Rab32/38 lokaliserer til melanosomene i tillegg til andre intracellulære organeller3,29. Våre studier viste at N-terminalen slettet STX13 mutant (GFP-STX13Δ129) lokaliserer til melanosomer og plasmamembran. Vi hypoteser at GFP-STX13Δ129 muligens kan brukes som reporter for å studere smuglingen fra RE til celleoverflaten og LRO. I motsetning til dette lokaliserer GFP-STX13WT til REer som ligner på Rab11. Våre studier illustrerte at GFP-STX13WT også kunne brukes til merking av REer i melanocytter. Vi spår at GFP-STX13WT kan være en bedre RE-markør enn Rab11 siden Rab11 overekspression endrer endosomal dynamikk. Til sammen fungerer GFP-STX13WT som en potensiell RE-markør for å studere dynamikken i steady-state-tilstand.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Forfatterne erklærer at de ikke har noen interessekonflikt.
Acknowledgments
Dette arbeidet ble støttet av Institutt for bioteknologi (BT/PR32489/BRB/10/1786/2019 til SRGS); Science and Engineering Research Board (CRG/2019/000281 til SRGS); DBT-NBACD (BT/HRD-NBA-NWB/38/2019-20 til SRGS) og IISc-DBT partnerskapsprogram (til SRGS). Infrastruktur i avdelingen ble støttet av DST-FIST, DBT og UGC. AMB ble støttet av DBT-JRF (DBT/2015/IISc/NJ-02).
Materials
| Name | Company | Catalog Number | Comments |
| anti-TYRP1 antibody (TA99) | ATCC | HB-8704 | |
| Fluoromount-G | Southern Biotech | 0100-01 | |
| Lipofectamine 2000 | ThermoFisher Scientific | 11668-500 | |
| Matrigel matrix | BD Biosciences | 356231 | |
| OPTI-MEM | ThermoFisher Scientific | 022600-050 | |
| Phorbol 12-myristate 13-acetate | Sigma-Aldrich | P8139 | |
| RPMI Medium 1640 | ThermoFisher Scientific | 31800-022 |
References
- Dell'Angelica, E. C. The building BLOC(k)s of lysosomes and related organelles. Current Opinion in Cell Biology. 16 (4), 458-464 (2004).
- Raposo, G., Marks, M. S. Melanosomes--dark organelles enlighten endosomal membrane transport. Nature Reviews in Molecular Cell Biology. 8 (10), 786-797 (2007).
- Ohbayashi, N., Fukuda, M. Recent advances in understanding the molecular basis of melanogenesis in melanocytes. F1000Research. 9, F1000 Faculty Rev-608 (2020).
- Theos, A. C., et al. Functions of adaptor protein (AP)-3 and AP-1 in tyrosinase sorting from endosomes to melanosomes. Molecular Biology of the Cell. 16 (11), 5356-5372 (2005).
- Di Pietro, S. M., et al. BLOC-1 interacts with BLOC-2 and the AP-3 complex to facilitate protein trafficking on endosomes. Molecular Biology of the Cell. 17 (9), 4027-4038 (2006).
- Setty, S. R., et al. BLOC-1 is required for cargo-specific sorting from vacuolar early endosomes toward lysosome-related organelles. Molecular Biology of the Cell. 18 (3), 768-780 (2007).
- Delevoye, C., et al. AP-1 and KIF13A coordinate endosomal sorting and positioning during melanosome biogenesis. Journal of Cell Biology. 187 (2), 247-264 (2009).
- Bultema, J. J., Ambrosio, A. L., Burek, C. L., Di Pietro, S. M. BLOC-2, AP-3, and AP-1 proteins function in concert with Rab38 and Rab32 proteins to mediate protein trafficking to lysosome-related organelles. Journal of Biological Chemistry. 287 (23), 19550-19563 (2012).
- Sitaram, A., et al. Differential recognition of a dileucine-based sorting signal by AP-1 and AP-3 reveals a requirement for both BLOC-1 and AP-3 in delivery of OCA2 to melanosomes. Molecular Biology of the Cell. 23 (16), 3178-3192 (2012).
- Nag, S., et al. Rab4A organizes endosomal domains for sorting cargo to lysosome-related organelles. Journal of Cell Science. 131 (18), (2018).
- Dennis, M. K., et al. BLOC-2 targets recycling endosomal tubules to melanosomes for cargo delivery. Journal of Cell Biology. 209 (4), 563-577 (2015).
- Jani, R. A., Purushothaman, L. K., Rani, S., Bergam, P., Setty, S. R. STX13 regulates cargo delivery from recycling endosomes during melanosome biogenesis. Journal Cell Science. 128 (17), 3263-3276 (2015).
- Shakya, S., et al. Rab22A recruits BLOC-1 and BLOC-2 to promote the biogenesis of recycling endosomes. EMBO Reports. 19 (12), 45918 (2018).
- Bowman, S. L., et al. A BLOC-1-AP-3 super-complex sorts a cis-SNARE complex into endosome-derived tubular transport carriers. Journal of Cell Biology. 220 (7), 202005173 (2021).
- Wei, M. L. Hermansky-Pudlak syndrome: a disease of protein trafficking and organelle function. Pigment Cell Research. 19 (1), 19-42 (2006).
- Bowman, S. L., Bi-Karchin, J., Le, L., Marks, M. S. The road to lysosome-related organelles: Insights from Hermansky-Pudlak syndrome and other rare diseases. Traffic. 20 (6), 404-435 (2019).
- Prekeris, R., Klumperman, J., Chen, Y. A., Scheller, R. H. Syntaxin 13 mediates cycling of plasma membrane proteins via tubulovesicular recycling endosomes. Journal of Cell Biology. 143 (4), 957-971 (1998).
- Mahanty, S., et al. Rab9A is required for delivery of cargo from recycling endosomes to melanosomes. Pigment Cell Melanoma Research. 29 (1), 43-59 (2016).
- Delevoye, C., et al. Recycling endosome tubule morphogenesis from sorting endosomes requires the kinesin motor KIF13A. Cell Reports. 6 (3), 445-454 (2014).
- Ha, L., et al. ARF functions as a melanoma tumor suppressor by inducing p53-independent senescence. Proceedings of the National Academy of Science U. S. A. 104 (26), 10968-10973 (2007).
- Soldati, T., Schliwa, M. Powering membrane traffic in endocytosis and recycling. Nature Reviews Molecular Cell Biology. 7 (12), 897-908 (2006).
- Grant, B. D., Donaldson, J. G. Pathways and mechanisms of endocytic recycling. Nature Reviews Molecular Cell Biology. 10 (9), 597-608 (2009).
- Hsu, V. W., Prekeris, R.
- Taguchi, T.
- Goldenring, J. R.
- Delevoye, C., Marks, M. S., Raposo, G. Lysosome-related organelles as functional adaptations of the endolysosomal system. Current Opinion in Cell Biology. 59, 147-158 (2019).
- Hsu, V. W., Bai, M., Li, J. Getting active: protein sorting in endocytic recycling. Nature Reviews in Molecular Cell Biology. 13 (5), 323-328 (2012).
- Desfougeres, Y., D'Agostino, M., Mayer, A. A modular tethering complex for endosomal recycling. Nature Cell Biology. 17 (5), 540-541 (2015).
- Le, L., Sires-Campos, J., Raposo, G., Delevoye, C., Marks, M. S. Melanosome biogenesis in the pigmentation of mammalian skin. Integrated Computational Biology. 61 (4), 1517-1545 (2021).
