

Summary
有糸分裂を受けている癌細胞でナントPJ-34の細胞傷害活性は、ライブ共焦点イメージングによりリアルタイムで文書化されました。 PJ-34根絶ヒト乳癌MDA-MB-231細胞分裂に余分中心体を保有する細胞。通常の双方向の焦点有糸分裂とは異なり、余分中心体は、PJ-34の存在下で2紡錘体極にクラスタ化されていなかった。
Abstract
強力PARP1阻害剤として作用しフェナントレン誘導体は有糸分裂におけるマルチ中心体ヒト癌細胞における過剰中心体の双方向の焦点クラスタリングを防いだ。ナントPJ-34は、最も強力な分子であった。余分中心体のデクラスタリングは、マルチ中心体細胞では分裂障害と細胞死を引き起こす。ほとんどの固体ヒトの癌は超中心体の高い発生を持っている。 PJ-34の活性はライブヒト乳癌MDA-MB-231中心体に高度に豊富である蛍光γ-チューブリン、および、蛍光ヒストンH2Bのプレゼントにコードするベクターでトランスフェクトされた細胞の共焦点イメージングによってリアルタイムで文書化されました染色。異常な染色体の手配やdeclustered中心体を表すデクラスタ化γ-チューブリン病巣はPJ-34で処理した後、トランスフェクトMDA-MB-231細胞で検出された。 2紡錘体極でアンクラスタ余分中心体は、細胞死に先行。これらの結果は、foのリンクさR初めて余分中心体細胞分裂におけるデクラスタリング、および細胞死に至る分裂不全のヒト癌細胞におけるPJ-34の最近検出された排他的な細胞傷害活性。 2中心体と双方向の焦点スピンドルと有糸分裂を受けて正常な細胞を損なうことなく、多中心体ともっぱら根絶癌細胞PJ-34、固定した細胞の共焦点イメージングにより観察前の調査結果による。 PJ-34のこの細胞傷害活性は、他の強力なPARP1阻害剤によって共有されていなかった、とPARP1阻害のその独立性を示唆し、PARP1欠損MEF宿すextracentrosomesで観察された。共焦点イメージングを生きるには、有糸分裂中の細胞を根絶する新しい分子を同定するための有用なツールを提供した。
Introduction
PJ-34を含むフェナントレン由来PARP1阻害剤は、ストレス条件下でのエネルギ消費PARP1媒介DNA修復(脳卒中または心筋梗塞)1によって誘導されるアポトーシス細胞死からの静止細胞を保護するように設計された。しかし、最近我々はPJ-34のことを発見し、その誘導PARP1阻害よりも倍高い濃度で、独占的にヒト癌細胞2,3における細胞死を引き起こす可能性があります。細胞の増殖であったより迅速に、細胞をより効率的に根絶であった。 PJ-34の細胞傷害活性は、有糸分裂2余分中心体デクラスタリングに起因するものであった。多くのヒト癌細胞の港4,5 multicentrosomes。静止細胞を損なうか、またはいくつかの良性の増殖細胞が有糸分裂で2中心体を保有することなく、20μMPJ-34と港過剰中心体は、効率的に72〜96時間以内にこれらの細胞を根絶ヒト乳癌細胞MDA-MB-231、のインキュベーション
バイポーラ中心体アセンブリは、有糸分裂4,5における双極紡錘体形成のために重要です。したがって、以上の2中心体を持つ細胞は、2つの極4-9で彼らの余分中心体をクラスタリング、ほとんど理解していない分子機構を開発しました。彼らの中心体の双極アセンブリの失敗は逮捕はG2 / Mの逮捕で、細胞周期、細胞死に至るが4,5分裂の失敗に起因することが多歪んスピンドルと異常な染色体の分離を引き起こす可能性があります。余分中心体デクラスタリングの基礎となる分子メカニズムは集中的に調査している<> 10(商標)。健康な組織の5,10を温存しながら、この死のメカニズムを理解することは、癌細胞の排他的な根絶が可能になります。
従って、細胞分裂異常細胞死を活性化する化合物は、共焦点画像は、超中心体のクラスタリングに影響する分子を同定するために使用することができることを示唆しているヒト固形cancers.Our結果の広い範囲で効率的であり得る選択的癌療法の新たなモードを提供有糸分裂2,3、薬剤候補をターゲットに、これらの化合物の癌をレンダリング。
我々は、固定やライブヒト癌細胞(有糸分裂で余分中心体の高い発生で)対正常細胞をスキャンすることによってナントPJ-34の細胞傷害活性を文書化している。ヒト癌細胞においてPJ-34の細胞傷害活性を同定するために用いられる撮像手順のステップバイステップの説明が以下に含まれる。
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1。細胞培養の準備
MDA-MB-231細胞は、ATCC( アメリカン·タイプ·カルチャー·コレクション )から購入し、液体窒素中で保存した。
- シード10 6 MDA-MB-231 Dulbeco改変イーグル培地(DMEM)、血清、1%L-グルタミン、及び1%Penstrep-アンホテリシンBのセルを許可し、10%の馬を含む10ミリリットル完全培地で92ミリメートル径シャーレ内の細胞約80-100%コンフルエントに増殖する。
- ディッシュから培地を取り外し、廃棄します。
- 0.25%血清のすべてのトレースを削除する(w / v)のトリプシン-EDTA溶液を用いて簡潔に細胞層を洗ってください。
- 細胞層(通常は5〜15分以内)に分散するまで倒立顕微鏡により細胞を観察すると、皿トリプシン-EDTA溶液2.0 mlを加える。
- 18ミリリットル完全増殖培地を加え、穏やかにピペットで細胞を吸引。チューブに移します。
- 1,200 rpmで細胞懸濁液を遠心。
- CUの24ミリリットルで細胞ペレットを再懸濁メディアをlture。
- 35ミリガラスボトムディッシュ(25%コンフルエント程度)とインキュベーター内の場所(5%CO 2、37℃)に細胞懸濁液2 mlを加える。
- ソリューション:
- 10%FBS、1%抗生物質(100単位/ mlペニシリンG、100μg/mlの/ mlのストレプトマイシン、ペン連鎖球菌 - 両性溶液)及び2mMのL-グルタミンを含むDMEM:細胞増殖のための完全培地。
- 0.25%トリプシン-EDTAを含有するトリプシン-EDTA溶液。
- 料理:
92ミリメートル直径ペトリ皿。
直径35mmのポリ-D-リジン被覆ガラス底培養皿。
2。ライブ共焦点イメージングのための細胞の調製
- シード2×10部1で述べたように2ミリリットル完全培地中のガラス底培養皿に5 MDA-MB-231細胞。細胞培養は、60〜70%の合流を(シャーレ当たり3-4×10 5細胞程度)に達すると、トランスフェクションに進みます。
- 2つのプラスミドは、エンコーディングで細胞をトランスフェクト製造プロトコルに従って、リポソームトランスフェクション試薬ジェット-PIを使用して融合タンパク質γ-チューブリン-GFP(中心体の蛍光検出用)およびヒストン-RED(H2B-RED、染色体の蛍光検出用)をING。簡単に言えば、100μlの塩化ナトリウム(150mMの)とチューブに各プラスミドから2μgのを混在させる。第2のチューブに100μlの塩化ナトリウム(150mMの)とトランスフェクション試薬(100μL)を混合し、室温(RT)で5分間インキュベートする。その後、二つの溶液を組み合わせたミックス(軽度の渦を使用)とダウンスピン。室温で30分間インキュベートする。
- トランスフェクション混合物のインキュベーション中、PBSで1回細胞を洗浄しないサプリメントと暖かいDMEM 2mlの(37°C)で細胞培地を交換してください。
- 穏やかにDMEMで細胞にトランスフェクション混合物を追加し、8時間(37℃、5%CO 2)インキュベーターにセルを返す。
- 24のためのインキュベーションの8時間後、2ミリリットル完全培地を含むDMEMを交換し、インキュベーターで細胞をインキュベートHR。
- トランスフェクション後24時間では、20μMPJ-34を含む2ミリリットル完全培地で細胞の培地を交換してください。
- さらに18時間(37℃、5%CO 2)、細胞をインキュベートする。
- 5%CO 2、37℃で細胞を維持するイメージングチャンバー内に少なくとも16時間の共焦点イメージングを生きるために件名のセル
- 並行して、以下のように蛍光顕微鏡を用いてトランスフェクション効率36時間後にトランスフェクションを調べます
- シード2 2ミリリットルの完全培地でウェル当たり1カバースリップを含む6ウェルプレートにおいて5×10 MDA-MB-231細胞。
- セクション2.2から2.5で述べたように細胞をトランスフェクトする。
- 36時間後にトランスフェクションは、冷メタノールでのインキュベーションによりカバースリップ上に取り付け、トランスフェクトされた細胞を固定:アセトン(1:1)ソリューション、7分、-20℃を
- 固定液を吸引除去し、化学フードで乾燥するように取り付け細胞とカバースリップしてみましょう。
- DAPIでゴールド退色試薬を延長し、目を聞かせて適用するeはカバースリップで6時間暗所で乾燥させる。
- 蛍光顕微鏡下でスライドを調べ、細胞の総人口からトランスフェクションされた細胞(赤と緑の信号)(DAPIによるDNA染色)の割合を計算します。 100-200細胞がカウントされたとき、所望のトランスフェクション率は20〜40%程度である。
3。ライブ共焦点イメージングスキャナ設定の技術的パラメータ
- SCANMODE XYZT;ホール[エアリー] 1.00;ズーム3.5、解像度8ビット;レーザーDPSS 561 nmで、アルゴン、可視レーザー488nmの;レーザー彼/ Neの見える633 nmで、客観HCX PL APO CS 63X 1.40 OIL UV、開口数1.4;速度700 Hzのをスキャンし、屈折率1.52。
- 画像3次元プレゼンテーションはIMARISイメージングソフトウェア7.0で調製した。
4。固定した細胞における有糸分裂の共焦点イメージング
- シード2×10 5個のMDA-MB-231乳癌細胞(ATCC)、正常マウス胚線維芽細胞(MEF)、またはPARP1欠損ME2ミリリットル完全培地で6ウェルプレートにカバーガラス上のF(博士フランソワーズDantzerにより調製しPARP - / - )。カバーガラスを2時間乾燥させ、6 - ウェルディッシュの各ウェルに入れ、滅菌DD水で洗浄することにより、以下の、96%エタノールで洗浄した。
- 培地にPJ-34(10-30μM)を追加し、必要な期間(通常は最大96時間)のために、細胞をインキュベート。
- PBS(リン酸緩衝生理食塩水)で1回カバースリップを洗って、氷冷メタノール中でインキュベーションを用いて細胞を固定:アセトン(1:1)ソリューション、7分、-20℃
- 固定液を吸引し、カバースリップ(この段階では、カバーガラスが数週間のために-20℃に保つことができる)化学フード中で乾燥させてください。
- 細胞膜を透過性およびRTで1時間PBST中の10%NDS(通常ロバ血清)( 'ブロッキング溶液')で細胞をブロックするPBST(PBS、0.1%のTween-20を補充)で1回カバースリップを洗浄する。
- プライマリーantibodiで透過固定された細胞をインキュベート室温で2時間、ES(スピンドルと中心体染色のため)。抗αチューブリン(1:250希釈)および抗-γチューブリン(1:200希釈):抗体は、以下のようなブロッキング溶液で希釈する。一次抗体は、次のように適用されています:6ウェルプレートカバー(カバーは逆さまである)上の各カバースリップのためのブロッキング溶液中の抗体の混合物を100μl(ドロップ)を適用。優しく抗体ドロップは、ドロップに直面して播種した細胞上にカバースリップを置く。室温で2時間撹拌抗体と対向するカバースリップをインキュベートする。
- ウェルに戻ってカバーガラスを置き、PBSTで細胞を3回洗浄する。その後、蛍光二次抗体を用いてカバーガラス上の細胞を標識するために4.6で説明したのと同じ手順を使用します。暗闇の中で、1時間、RT用の二次抗体を用いてカバーガラス上の細胞をインキュベートする。抗体は、以下のようブロッキング溶液で希釈する。;とアレクサフルーア568(1:1,000希釈アレクサフルーア488(緑1:1,000希釈);赤)。
- 使用してカバースリップはDAPI(染色体染色用)とGold退色防止試薬を延長し、乾燥させる暗闇の中で室温で一晩インキュベートマウント。
- 共焦点顕微鏡によるカバースリップを調べます。
5。 ATP生産により測定し、細胞の生存率
ATP産生は、発光ATP検出アッセイキットにより測定される。
- 各ウェルに800μlの培地中の種子96ウェルプレート中の細胞、約20,000のセルを。三ブランクウェルは、媒体のバックグラウンド発光を決定するために使用されるべきである。
- 100μMに約10μMからATP標準希釈系列を用意し、氷上に保つ。
- 各ウェルに洗剤液50μlを加え、オービタルシェーカー、700回転で5分間プレートを振る。
- キット内の '基質緩衝' 5mlの '凍結乾燥基質'の再構成する各バイアル。
- ウェルに再構成された基質溶液50μlを加え、振とうオービタルシェーカー、700回転で5分間プレート。
- 10分間、暗闇の中でプレートを保管してください。
- ELISAマイクロプレートリーダーで各ウェルのluminiscenceを測定します。
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
PJ-34安定した水溶性のフェナント1( 図1)である。我々の以前の結果は、PJ-34で処理した固定マルチ中心体癌細胞のいくつかのタイプに細胞死および非クラスタ化された余分中心体を明らかにした。対照的に、正常な増殖細胞を2,3損なわれていなかった。中心体は、2つの固定余分中心体細胞でcentrine1とγ-チューブリンに対する抗体で二重標識により同定した。
ここでは、PJ-34の細胞傷害活性は、ライブ共焦点顕微鏡を用いて、リアルタイムでこれらのライブ余分中心体細胞で記述されていました。 4,5余分中心体の高い発生を(> 50%)を持っているMDA-MB-231細胞は、γ-チューブリン-GFPをトランスフェクトした細胞に焦点を当てた共焦点イメージングにより、少なくとも16時間のためにスキャンされた(ヒト乳癌ライブ蛍光γ-チューブリン巣2のラベル)とヒストンH2B-RED(蛍光ラベと染色体の玲)。六から十生きたトランスフェクションされた細胞は、それぞれの実験で平行にスキャンした。 centrin1でトランスフェクトした細胞におけるγ-チューブリン病巣の二重免疫標識は技術的に不可能であった。
分散されたγ-チューブリン病巣および異常染色体配置はほとんど有糸分裂でランダムに選択された未処理のMDA-MB-231細胞では検出されなかった。余分中心体の遠近両用クラスタリングを表すγ-チューブリン病巣の遠近両用クラスタリングが、ライブ未処理のMDA-MB-231細胞( 図2)の大部分で文書化され、それとは対照的に、非クラスタ化された中心体と染色体の異常な配置は、ライブで検出されたPJ-34(20μM)、および細胞死( 図3)によって終了これらの細胞における有糸分裂と共にインキュベートMDA-MB-231細胞をトランスフェ。これらの細胞は18 PJ-34とインキュベートした- 24時間を( 図3)スキャン中にスキャンし、追加の16時間前。セルデのリアルタイムドキュメント有糸分裂中にアテローム強く有糸分裂とPJ-34(20μM)2でインキュベートされた細胞における細胞死の割合が多極紡錘によるヒト悪性細胞の数との間に以前に定義された正の相関をサポートしています。
PJ-34強力PARP1阻害剤1として機能します。したがって、我々は分裂障害に関連付けられているPARP1阻害引き起こす細胞死の可能性を検討した。ライブイメージングとは異なり、固定MDA-MB-231細胞のイメージングすることにより、ヒト癌細胞株の様々なPARP1阻害剤の効果の統計的分析を可能にする、細胞培養物中の細胞の大集団の検査を可能にした。 ( 図4)PJ-34の活性は正常で他の強力な、非フェナントレンPARP1阻害剤またはPARP1欠損細胞( - / - )マウス胚線維芽細胞(MEF) すなわち正常とPARP1()の活性と比較した。 PARP1欠損MEF港有糸分裂における多中心体が、彼らはNOですtの腫瘍細胞11。これらの細胞は、博士フランソワDantzer、ストラスブール、フランスによって調製した。
通常の固定およびPARP1( - / - )MEFは2の前に報告されているように、それぞれ、そのスピンドルと中心体のラベルが付いたα-およびγ-チューブリンのために免疫標識された。調べた細胞培養のいくつかは、ABT-888とAG01469、PARP1の酵素活性を阻害し、BSI-201、明らかにPARP1結合を減衰する化合物を含むPJ-34やその他の強力な、非フェナントレンPARP1阻害剤で処理したDNA 12-14ニック。濃度阻害PARP1活動( 図4)で、通常のMEFを損なわテスト済みPARP1阻害剤なし。 ( - / - )MEF( 図4AおよびB)とは対照的に、PJ-34用量依存的にγ-チューブリン病巣、PARP1でスピンドルと細胞死の歪みの未クラスタリングに起因する。これは、PJ-34(FIGURで処理正常MEFでは観察されなかった電子4B)またはPARP1で( - / - )MEFはで処理非phenenthrene PARP1阻害剤ABT-888またはAG014699( 図4C)。 ( - / - )MEFそれは通常のMEFがPARP1よりPJ-34活性に対してより耐性であったが、20μMを超える濃度でPJ-34は、通常のMEFを損なわなかったことに留意すべきである。
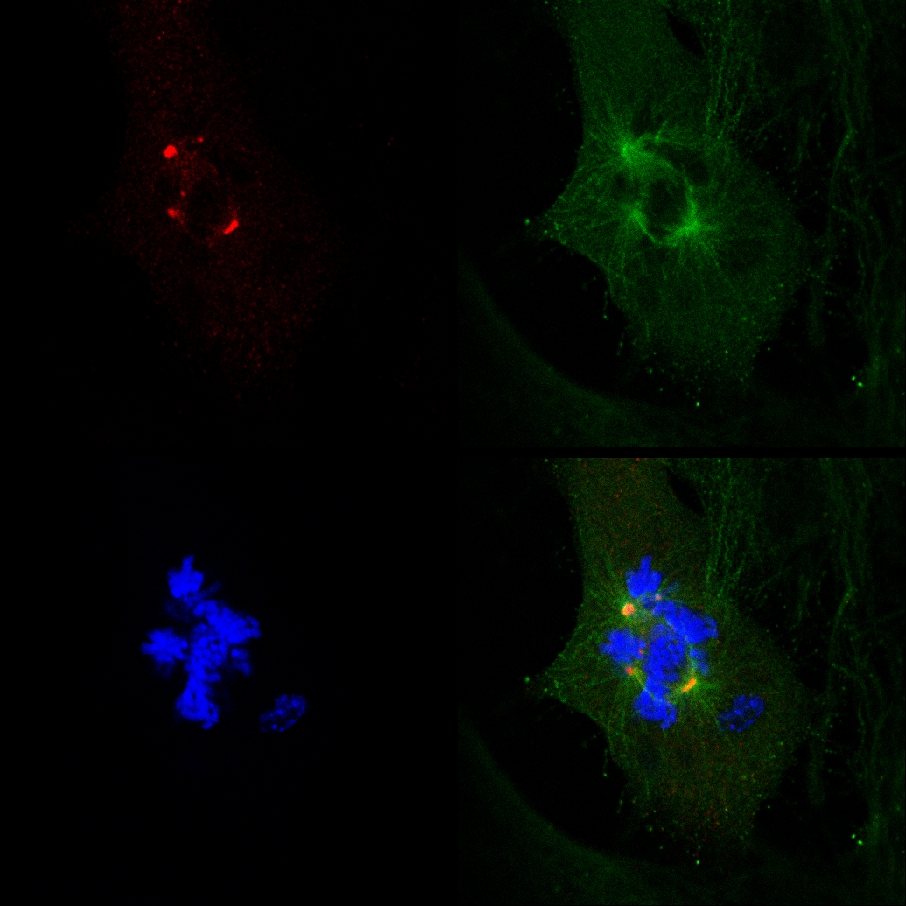
MEFは、より高い濃度でPJ-34とインキュベート自分PARP1欠乏にもかかわらずMEF、およびPARP1における多焦点スピンドルと細胞根絶の形成の間の相関関係( - / - )( - / - )PJ-34根絶PARP1という事実( - / - )MEFとPARP1阻害( 図4A)PARP1阻害に必要なものは、余分中心体PARP1でデクラスタリングの間に因果関係と一致しませんでした。 PARP1でPJ-34の細胞傷害活性( - / - )MEFが良いマルチ中心体細胞での余分中心体デクラスタリング剤2としての活動によって説明できる( 補足情報 "/> 図3。 余分中心体ラベル、非クラスタ化されたγ-チューブリンと有糸分裂にランダムに選択されたMDA-MB-231細胞における細胞死のライブ共焦点イメージングのマニュアルを参照してください。 MDA-MB-231細胞を走査する前に、ライブ共焦点画像の16時間の間に24時間PJ-34でインキュベートした。 PJ-34はγ-チューブリン-GFP(緑色、γ-チューブリン焦点と中心体)とH2B-RED(赤、染色体)を発現するベクターでトランスフェクション後24時間を適用した。 
図1。ナントPJ-34:N-(6 -オキソ-5,6 -ジヒドロ-phenanthridin -2 -イル)-N、N-ジメチルアセトアミド。 
図2。有糸分裂にランダムに選択された生きたMDA-MB-231細胞に余分中心体のバイフォーカルクラスタリング。A.上部パネル :γ-チューブリン-GFP 下のパネルでトランスフェクションランダムに選択された生きたMDA-MB-231細胞内で標識された中心体:ヒストンH2B-REDでトランスフェクションランダムに選択されたMDA-MB-231細胞における有糸分裂時の染色体の再アレンジ。ランダムに選択されたカルトで識別クラスタ余分中心体とB.バイフォーカル有糸分裂ured MDA-MB-231細胞。とヒストンH2B-RED(ラベリング染色、赤)、細胞が両方のγ-チューブリン-GFP(緑色ラベルγ-チューブリン病巣)でトランスフェクトした。 48時間のトランスフェクション後、細胞を16時間ライブ共焦点画像に暴露した。 6つのセルを各実験で平行にスキャンした。四つの異なる実験を行った。また、補足情報を参照してください。 より大きい数字を表示するには、ここをクリックしてください 。 
図3。 PJ-34で処理されたライブMDA-MB-231細胞。細胞死によって終了散乱中心体と有糸分裂にランダムに選択された生きたMDA-MB-231細胞( 左側の1 番目のフレーム () 内に細胞死を前に余分中心体デクラスタリング2 番目と3 番目のフレーム )。このCEと(ヒストンH2B-RED; LLはランダムγ-チューブリン-GFP(緑色の中心体を含むラベルγ-チューブリン病巣)を発現するベクターでトランスフェクション後24時間を適用してPJ-34(20μM)で24時間インキュベートし、細胞培養で選択されたラベリング染色、赤)。細胞は、ライブ共焦点撮像して16時間スキャンした。 6つのセルを各実験で平行にスキャンした。三つの異なる実験を行った。また、補足情報を参照してください。 より大きい数字を表示するには、ここをクリックしてください 。 
図4。 PARP1でPJ-34の細胞傷害活性( - / - )マウス胚性線維芽細胞。 A.(左 )は、nで計算多焦点スピンドルの割合指示された濃度でPJ-34で48時間インキュベート(灰色の線)MEF、 - ormal(黒線)とPARP1 - /。多極紡錘の割合は、3つの異なる実験で検出された合計20スピンドルの外に算出した。コントロール未処理細胞の生存率( - / - (灰色の線)MEFノーマル(黒線)とPARP1)に対してPJ-34(20μM)で72時間インキュベートした細胞培養で検出された( 右 )減少細胞の生存。細胞生存は細胞のATP産生(プロトコル5)により測定した。 3異なる実験の各セルラインの4の測定の平均値は、B.スピンドルがランダムに選択された固定正常に提示しPARP1ている- / -細胞分裂におけるMEF、未処理(対照)または指定で48時間、PJ-34とインキュベート濃度。 PJ-34は、多極紡錘を引き起こした。細胞は、固定されて透過性とのために免疫標識されたα-およびγ-チューブリン(緑スピンドルのラベリングとそれぞれ中心体の赤ラベル、)。染色体はDAPI試薬(青)で標識した。 3異なった実験の代表的な結果は、C.強力な非フェナントレンPARP1阻害剤はPARP1でクラスタリング中心体に影響を及ぼさなかった( - / - )MEFを。ランダムに選択されたノーマルのスピンドルとPARP1は- / - MEFが提示され、未処理のMEF(コントロール)またはMEF非フェナントレンPARP阻害剤、AG01469(20μM)またはABT888(20μM)で48時間処理した。染色体はDAPI試薬(青)で標識される。同様の結果が、3つの異なる実験で得られた。 より大きい数字を表示するには、ここをクリックしてください 。
大きい数字を表示するには、ここをクリック 。 
大きい数字を表示するには、ここをクリック 。
図2B。補足図。余分中心体ラベルされた双方向の焦点クラスタ化されたγ-チューブリンと後期にランダムに選択されたMDA-MB-231細胞は48時間γ-チューブリン-GFPを発現するベクターでトランスフェクトされた後、ライブ共焦点イメージングにより、16時間のためにスキャンされ、 H2B-RED(固定した細胞(緑)でラベルγ-チューブリン焦点と中心体と染色体のヒストンH2Bラベル(赤)、それぞれ)。
Subscription Required. Please recommend JoVE to your librarian.
Discussion
ライブ共焦点イメージングは、有糸分裂( 図3と補足情報)の間にライブマルチ中心体細胞ではPJ-34の細胞毒性効果をリアルタイムに資料を提供した。これは、PJ-34 5-9で細胞分裂異常細胞死の誘導を示唆し、余分な中心体デクラスタリングと細胞死にヒト癌細胞におけるPJ-34の細胞毒性を帰初のライブドキュメントだった。対照的に、超正規の中心体の双方向の焦点クラスタリングは、双方向focaliクラスタ化余分中心体( 図2、補足情報)を通常の有糸分裂を受けるライブ未処理のMDA-MB-231細胞で観察された。
これらの結果によると、トランスフェクションされた細胞のライブ共焦点イメージングは、有糸分裂5,9,15,16時のトランスフェクションされた細胞内の余分-中心体クラスタリングに関与するタンパク質の移行を検出するために有用である可能性があります。マルチcentrosでPJ-34の影響を受けたタンパク質の同定OMAL細胞は余分中心体デクラスタリングによって活性死メカニズムを理解するためのいくつかの手がかりを提供することがあります。
生細胞内の有糸分裂の間にリアルタイム情報を提供するライブ共焦点イメージングの利点はまた、いくつかの限界にバインドされている。実験ごとにスキャンされた細胞の数は限られている。したがって有糸分裂における細胞を検出する可能性は低く、いくつかの繰り返し実験は、スキャンされた細胞における有糸分裂のリアルタイムドキュメントに必要とされる。また、成功は標識タンパク質を発現するベクターによる細胞のトランスフェクションの高効率化に大きく依存している。したがって、信頼性があるにも関わらず、ライブ共焦点イメージングは時間がかかり、非常に経験豊富な働き者が求められています。
比較では、固定された細胞の免疫細胞化学および共焦点画像は、信頼性の統計分析のために必要とされる実験当たりのセルの数が多いの検査を可能にする。私たちは、比較するためにこのメソッドを使用過剰中心体2,3と有糸分裂を受けている癌細胞におけるその効果への通常の良性細胞の有糸分裂でPJ-34の影響。同様に、この方法は、PARPにおける非フェナントレンPARP1阻害剤のそれにPJ-34の効果を比較するために使用された( - / - )MEFは、余分中心体(マルチ中心体11の高い発生に細胞)( 図4)を保有する。
要約すると、我々の結果は、固定された細胞の細胞化学および共焦点解析を有糸分裂における生細胞の共焦点画像( 図2および図3)によって提供される最も貴重なリアルタイムの情報を組み合わせることの利点を示している。これらの方法の組み合わせは、細胞の生存のための重要な特定のメカニズムをターゲットに、PJ-34のように、小分子の細胞傷害活性を同定するために有用かもしれない。その増殖やsurvivのための余分中心体双極クラスタリングで多くのヒト癌細胞のユニークな依存アルは、PJ-34癌治療のための可能な候補をレンダリングします。
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
著者は、彼らが競合する経済的利益を持っていないことを宣言します。
Acknowledgments
本研究の資金源:テルアビブ大学の技術移転会社、运动とシェバ·メディカルセンター(M. CA及びSI)、ICRFの共同ファンド - イスラエルのがん研究財団(M. CA。)とイスラエル科学財団( SI)。
Materials
| Name | Company | Catalog Number | Comments |
| REAGENTS | |||
| DMEM | Invitrogen (GIBCO) | 41965 | |
| FBS (Fetal bovine Serum) | Invitrogen (GIBCO) | 12657 | |
| Pen-Strep-Ampho solution | Biological Industries, Israel | 03-033-1B | |
| L-glutamine | Invitrogen (GIBCO) | 25030-024 | |
| 0.25% Tripsin-EDTA | Invitrogen (GIBCO) | 25200 | |
| 92 mm Petri dishes | Nunc,Thermo scientific | 150350 | |
| 35 mm poly-D-lysine coated glass bottom culture dishes | MatTek Corporation, USA | P35GC-0-14-C | |
| Luminescent ATP detection assay kit | Abcam | ab113849 | |
| NDS (Normal Donkey Serum) | Jackson ImmunoResearch | 017-000-121 | |
| Anti α-tubulin antibody | Sigma | T9026 | 1:250 dilution (IF) |
| Anti γ-tubulin antibody | Sigma | T5192 | 1:200 dilution (IF) |
| Alexa Fluor 488 Goat Anti-Mouse IgG | Invitrogen | A-11017 | 1:1,000 dilution (IF) |
| Alexa Fluor 568 Donkey Anti-Rabbit IgG | Invitrogen | A-10042 | 1:1,000 dilution (IF) |
| ProLong Gold antifade reagent with DAPI (mounting) | Invitrogen | P36935 | |
| JetPEI (liposomal transfection reagent ) | Polyplus | 101-10 | |
| EQUIPMENT | |||
| Confocal microscope | Leica (Mannheim, Germany) | TCS SP5II | |
References
- Jagtap, P., et al. Novel phananthridine inhibitors of poly(adenosine 5'-diphosphate-ribose) synthetase: Potent cytoprotective and antishock agents. Crit. Care Med. 30, 1071-1082 (2002).
- Castiel, A., et al. A small molecule exclusively eradicates human cancer cells: Extra-centrosomes de-clustering agent. BMC Cancer. 11 (1), 412 (2011).
- Inbar-Rozensal, D., et al. A selective eradication of human nonhereditary breast cancer cells by phenanthridine -derived polyADP-ribose polymerase inhibitors. Breast Cancer Res. 11 (6), R78 (2009).
- Gergely, F., Basto, R. Multiple centrosomes: together they stand, divided they fall. Genes Dev. 22, 2291-2296 (2008).
- Godinho, S. A., Kwon, M., Pellman, D. Centrosomes and cancer: how cancer cells divide with too many centrosomes. Canc. Met. Rev. 28, 85-98 (2009).
- Doxsey, S. Re-evaluating centrosome function. Nat. Rev. Mol. Cell Biol. 2, 688-698 (2001).
- Walczak, C. E., Heald, R. Mechanisms of mitotic spindle assembly and function. International Rev. of Cytology. 265, 111-158 (2008).
- Ogden, A., Rida, P. C. G., Aneja, R. Let's huddle to prevent a muddle: centrosome declustering as an attractive anticancer strategy. Cell Death Differ. 19, 1255-1267 (2012).
- Kramer, A., Anderhub, S., Maier, B. Mechanisms and Consequences of centrosomes clustering in cancer cells. The Centrosome: Cell and Molecular mechanisms of functions and disfunctions in disease. Schatten, E. , Humana Press Springer. 255-277 (2012).
- Galimberti, F., et al. Anaphase Catastrophe Is a Target for Cancer Therapy. Clin. Cancer Res. 17, 1218-1222 (2011).
- Kanai, M., et al. Haploinsufficiency of poly(ADP-ribose) polymerase-1-mediated poly(ADP-ribosyl)ation for centrosome duplication. Biochem. Biophys. Res. Commun. 359, 426-430 (2007).
- Gartner, E. M., Burger, A. M., Lorusso, P. M. Poly(ADP-ribose) polymerase inhibitors: a novel drug class with a promising future. Cancer J. 16, 83-90 (2010).
- Wahlberg, E., et al. Family-wide chemical profiling and structural analysis of PARP and tankyrase inhibitors. Nature Biotechnology. 30, 283-288 (2012).
- Rouleau, M., Patel, A., Hendze, M. J., Kaufmann, S. H., Poirier, G. G. PARP inhibition: PARP1 and beyond. Nature Rev. Cancer. 10, 293-301 (2010).
- Leber, B., et al. Proteins Required for Centrosome Clustering in Cancer Cells. Sci. Transl. Med. 2 (33), 33-38 (2010).
- Kwon, M., et al. Mechanisms to suppress multipolar division in cancer cells with extra centrosomes. Gene Dev. 22, 2189-2203 (2008).

{kind=link}
{kind=link}