Nevronale vev er svært sammensatt; en heterogen celle blanding som er romlig organisert innenfor definerte lag og avdelinger, og med plast tilkobling via celle kontakter og spesielt via axon og dendrite utvekster. Nye teknikker er nødvendig for å gi større eksperimentell frihet til å få dypere innsikt og avdekke mekanismene er sentrale for sykdom, utvikling og sunn funksjon. Den Campenot kammeret 1,2 og mer nylig microfabricated utførelses 3,4 kan anvendes for ex vivo fremstilling av nettverks neuronale ko-kulturer med evne til selektivt å forstyrre de forskjellige somatiske populasjoner og også deres neurite utvekster. Disse microfluidic enheter har for eksempel blitt brukt til å studere axon degenerasjon og regenerasjon følgende kjemiske 5,6 eller laser axotomy 6-8, tauopathy 9, viral spredning 10,11, og mRNA lokalisering i axons fire.
ent ">For å utvide rekkevidden av nevrobiologer, er den teknologiske utviklingen som kreves for å forberede minimalis nevrale co-kulturer. Dette gjør det mulig floke av nevronale nettverk for etterforskningen av systemet med én celle og sub-cellulære presisjon. Kravet til minimale celletall åpner muligheten for å analysere sjeldne celletyper, inkludert substantia nigra dopaminerge celler som er relevante for Parkinsons sykdom, spiral ganglia fra øret, perifere nerveceller, og stamceller. Utover dette, er mobilnettet økonomi relevant for 3Rs initiativet. Ved hjelp av disse microfluidic plattformer, storskala toksisitets skjermer eller annen høy gjennomstrømning, kan data rik eksperimentelle serien krever dyre nevroner nå vurderes.
I dette papir protokoller for fremstilling og bruk av en mikrofluidenhet er beskrevet. Microfluidic Oppstillings i kombinasjon med en in situ biomaterial mønster metoden kan brukes for registrering av sterkt sammenhengende neuronale ko-kulturer ved bruk av minimale antall celler. Microfluidic oppstillings er basert på en differensialstrømnings tilnærming 12-15, idet mikrostrukturerte feller er plassert innenfor et fluidic krets (illustrert sammen med SEM-bilder i figur 1). Banen 0 → 1 har det nedre fluidic motstand (R 2> R 1) for transport av neuroner på en lineær oppstilling av mikrostrukturerte åpninger – inntakene til de neurite utvekst kanaler. Occupancy av fellen av en enkelt celle lokalt hindrer flyten å avlede strømlinjene for fangst påfølgende celler i nabo feller. Fullstendig belegg av fellene i matrisen kopler fluidic forholdet (R1> R 2) for å avlede strømlinjene i det serpentinbane (0 → 2) for å frembringe en bypass-modus for operasjonen for fjerning av overskytende neuroner.
<p class = "jove_content" fo: keep-together.within-page = "always">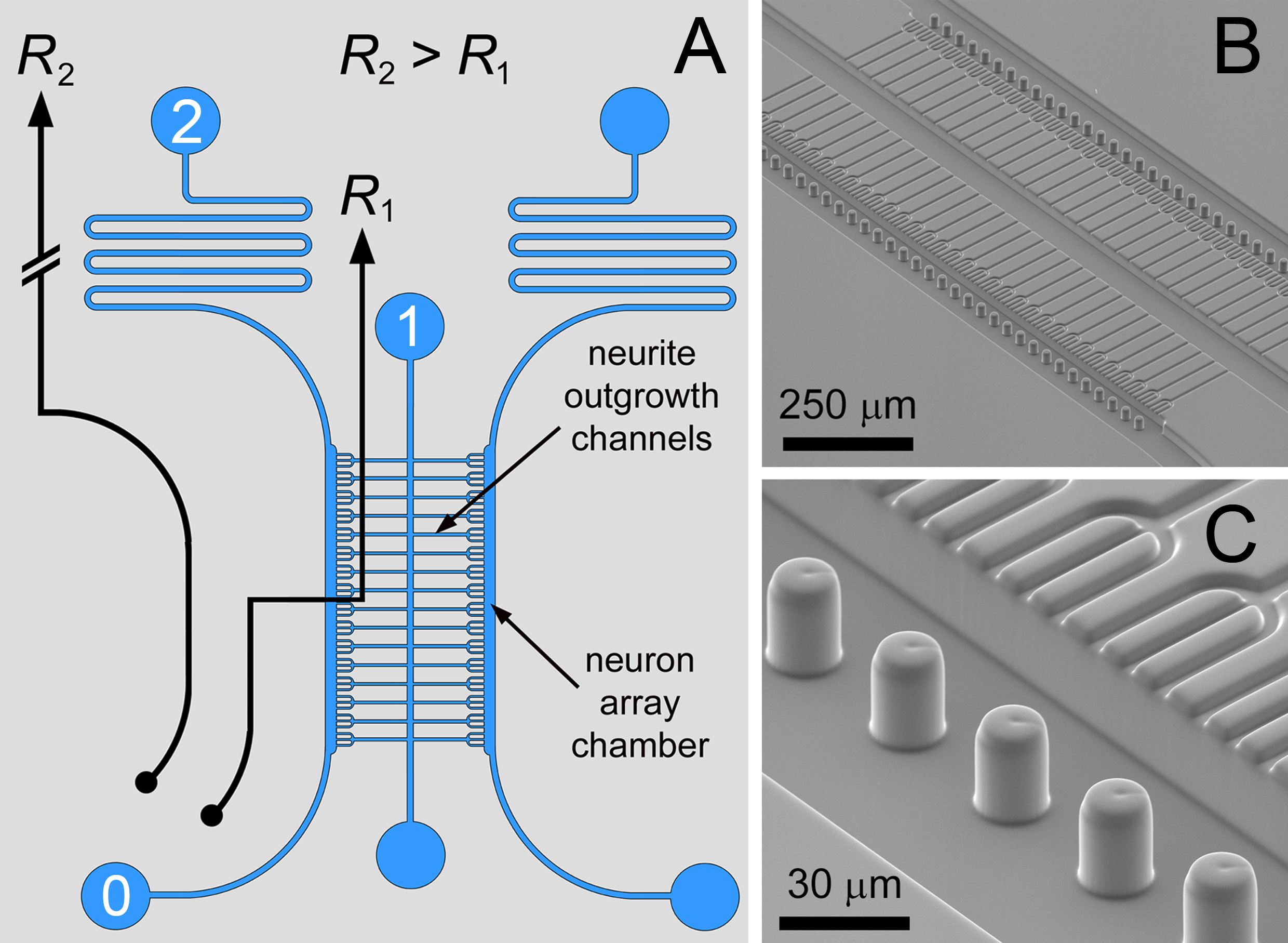
Figur 1 Microfluidic Circuit.. A) Den differensielle motstand fluidic krets for enkelt neuron oppstillings, med flankekulturkamre sammen av neurite utvekst kanaler. B, C) SEM bilder av bilayer compartmentalized neuron ko-kultur matrise med menisk låsing micropillars. Med denne designen, ble Trident-formet nevron fangst strukturer brukes til å fremme Fascikulasjoner av neurite utvekster. Figur og legende gjengitt med tillatelse fra Royal Society of Chemistry (RSC) 12. Klikk her for å se større bilde.
Fremstillingen av micropatterned nevrale nettverk på plane underlag kan lett oppnås (for exampl es fra vår gruppe, se Frimat et al. 16 og Heike et al. 17). Men innkapsle bioaktive materielle mønstre innen PDMS-enheter og med kravet om mikrometer-skala justering av disse til microfluidic kanalene utgjør en stor teknisk utfordring. I avsnitt 3.1 en protokoll for i-chip, eller in situ, utarbeidelse av biomateriale mønstre presenteres. Disse mønstrene gjør at nervecellen registrering under langvarige kultur tidsrammer og fremme utvekster mellom avdelinger. Menisk-låsing mikrostrukturer brukes til å justere en såkalt vann maske med nervecellen arraying nettsteder og neurite utvekst kanaler. Vannet masken beskytter adhesjonsmolekyl belegg under plasmabehandling, mens det eksponerte overflater er oppløst for å definere biomateriale mønster. I tillegg er protokoller levert for cellekultur, og for fluidic isolasjon som er nødvendig for selektiv behandling av de forskjellige ko-kultur seksjonene.
ve_content "> er Protokollene utviklet for å utnytte brukervennlige prinsipper for myk litografi for replikering av poly (dimethylsiloxane) (PDMS) microfluidic enheter 18. Tilsvarende, i situ biomateriale mønster er grei, utnytte fordampning og overflatespenning fenomener, og bare krever en billig håndplasmakilde. microfluidic kretsen effektivt programmer celle lasting og kupé spesifikke behandlinger gjør disse operasjonene bare et spørsmål om dispensermaterialer inn i riktig bunn port og aspirere ovenfra. På denne måte er det til hensikt å gi neurobiologists frihet til å forberede og bruke microfluidic enheter i egne laboratorier.