Representative images of quick immuno-FISH are shown in Figure 1A. Co-localization of the Xist RNA cloud and H3K27me3 signal on the Xi was detected in differentiating female cells. At day 12 upon differentiation, more than 90% of EB cells had an Xist cloud (Figure 1B). Short oligonucleotide probes efficiently penetrated into the nuclei, leading to a visualization of almost all H3K27me3 signals co-localized with Xist RNA (Figure 1C; 97%, n = 150).
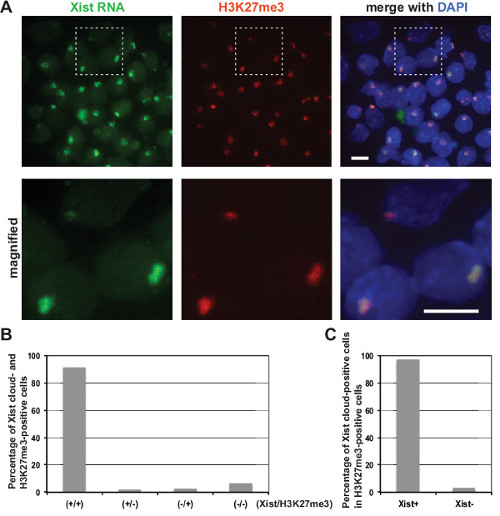
Figure 1: Xist RNA and H3K27me3 co-localize on the Xi in differentiating mouse EB. (A) Immuno-FISH for Xist RNA with H3K27me3 modification in differentiating female EB cells at day 12 upon differentiation. Xist probes were labeled with Alexa Fluor 488, and anti-H3K27me3 primary antibody was detected by anti-mouse IgG Alexa Fluor 555-conjugated secondary antibody. Nuclei were counterstained with DAPI. The boxed region is magnified in the bottom panels. Scale bars: 10 µm. (B) Frequency of Xist cloud- and H3K27me3-positive cells in EB at day 12 upon differentiation. (C) Frequency of co-localization of Xist cloud with H3K27me3 signal in EB at day 12 upon differentiation. Please click here to view a larger version of this figure.
| Xp1 | ggtaagtatccaaaaccccgttgg |
| Xp2 | cgatcagcagcaacagtacacg |
| Xp3 | gcaattggttgcttttatccagtcc |
| Xp4 | cgcaacaccgcacactaatacg |
| Xp5 | gccatcttagacacattcaagagcat |
| Xp6 | cacacgtgaagtaccaagcgaaac |
| Xp7 | gccacgtatagagcactgtaagagactatg |
| Xp8 | caaagcacactatcagacgtgtcg |
| Xp9 | ggagaatctagatgccataaaggcaag |
| Xp10 | tgatggacactgcattttagcactg |
| Xp11 | ggacactgcattttagcaatacgattc |
| Xp12 | gtgatgggcactgcattttagc |
| Xp13 | agatgggctatctcagtcttataggct |
| Xp14 | ggaagtcagtatggagggggtatg |
| Xp15 | tgggactgtgactactacagcaatga |
| Xp16 | aagactcaattcctagtcaggattatccac |
| Xp17 | gggctgtagctctatgacagtgcttt |
| Xp18 | gtcttcaccagatgcagattactacagtg |
| Xp19 | aatagtttgaggaaggggtttcaagtg |
| Xp20 | gctgttcaggtttccttctgtagtga |
| Xp21 | gcaagagatacaatggtccgaaaagt |
| Xp22 | agcacttcgtacaaccctctttctg |
| Xp23 | gaagagagcaggtcattcgtcagag |
| Xp24 | gcaactgagacactgtagccatatgaag |
| Xp25 | ttcctggaggaagaacggaaaga |
| Xp26 | tgattagaaggcttaggtcatcttcca |
| Xp27 | ttttgttcagagtagcgaggacttga |
| Xp28 | aatagagcagaatggcttcctcgaa |
| Xp29 | acattgcttgatcacgctgaagac |
| Xp30 | gcaaggaagaaatagacacacaaagc |
| Xp31 | ggaagaaatagatgtaacaaagaattagacaca |
| Xp32 | cacttcagagccacttgaatcctg |
| Xp33 | agtcacaggtgtcctgtagaaacagttc |
| Xp34 | cctttatgggcaatggcaacaat |
| Xp35 | ggcacatctgcatattgcttgtcta |
| Xp36 | gcaactaagaccatgaacccacaa |
| Xp37 | aaacacactggccttaagtatatggactg |
| Xp38 | cattcatttgcacacatggaacaat |
| Xp39 | tgggagacaatatttagcctccaggt |
| Xp40 | cctagcaagggcactgttttgtaataa |
| Xp41 | taacatttagcacactgccttgcac |
| Xp42 | cagtgatctacactaggtccacctcaca |
| Xp43 | ttatgttgaaggaatcttggccttg |
| Xp44 | aagtgagagctgtagtctcaaggtgtga |
| Xp45 | gtattcaacctctgaggcaaactgtg |
| Xp46 | agattgtggaacttagatggctgtca |
| Xp47 | tggaactgcattaaagtcccaacttag |
| Xp48 | gaactcccagacctcttcaacctg |
Table 1: Sequence of fluorescently labeled oligonucleotides for Xist RNA FISH. The 48 oligonucleotides were synthesized with a 5’-amino modification to prepare fluorescently labeled oligonucleotide probes for Xist RNA FISH.
