

Abstract
Нервно-мышечного соединения (НМС) является большой, холинергическая реле синапс, через который млекопитающих моторные нейроны контролировать добровольное сокращение мышц. Структурные изменения в НМС может привести к поломке нервам, в результате слабости, атрофии и даже смерти мышечного волокна. Многие исследования изучали, как генетические изменения или болезнь может привести к изменению структуры НМС мыши. К сожалению, это может быть трудно непосредственно сравнивать результаты этих исследований, так как они часто используют различные параметры и аналитические методы. Три протокола описаны здесь. Первый использует Проекция максимальной интенсивности конфокальных изображений для измерения площади рецептора ацетилхолина (AChR) обогащенный постсинаптической мембраны доменов на концевой шайбы и области синаптической окрашивания пузырьков в вышележащей пресинаптических окончаний нервов. Второй протокол сравнивает относительные интенсивности иммунного окрашивания для синаптических белков в постсинаптической мембране. Третий прotocol использует флуоресцентного резонанса переноса энергии (FRET), чтобы обнаружить изменения в упаковке постсинаптических АХР на торцевой пластины. Протоколы были разработаны и уточнены в течение ряда исследований. Факторы, влияющие на качество и согласованность результатов обсуждаются и нормативные данные предоставляются НМС в здоровых молодых взрослых мышей.
Introduction
Нервно-мышечном соединении (НМС) является критическим реле синапсов, что опосредует связь между нервной системы и скелетных мышцах. Это требуется для всех произвольных движений. Флуоресцентная микроскопия уже давно используется для изучения влияния трансгенов на мышь NMJ 1-3 или сравнить эффекты от возраста, диеты, физические упражнения и болезни на грызунов NMJs 4-11. Такие исследования научили нас много о физиологии и патофизиологии НМС, но различные параметры сообщалось (например, AChR площадь, Концевая пластина площадь, длина периметра, индексы фрагментации) часто затрудняют сравнение результатов этих исследований. Существует растущая ожидание доклинические исследователи, чтобы иметь возможность продемонстрировать воспроизводимость, в частности, в исследованиях с грызунами моделей болезни 12. Протоколы, описанные здесь, были уточнены в ходе ряда исследований, которая расследовала развития, физиологических и патофизиологических чАнгелов в НМС. Такие исследования требуют измерения области синаптических специализаций на двигателе мыши концевой пластинки и относительной плотности упаковки синаптических белков в постсинаптической специализаций 13-15.
Полезность этих методов свидетельствуют недавние исследования в мышиной модели анти-мускуса миастения. Ежедневные инъекции IgG из анти-мускусный положительного миастения пациентов в взрослых мышей заставило их стать слабыми в течение 2 недель 16. Конфокальные максимальной проекции изображения мышечных разделов, которые были дважды мечеными для синаптофизина (в нервных терминалов) и постсинаптических АХР показали прогрессирующее снижение в области АХР окрашивания в качестве основного изменений. Важно темпы снижения было достаточно, чтобы объяснить, сопоставимые снижение амплитуды синаптических потенциалов, отказ от синаптической передачи и мышечной слабости 17,18. Качественно аналогичные результаты были получены и другими исследовательскими группами10,19. Те же НМС методы измерения, поскольку были использованы для оценки влияния трех препаратов для лечения анти-мускуса миастения в этой модели мыши 20,21.
Сидячий старение может привести к потере нервно-мышечных соединений. Протоколы, описанные здесь, выявили снижение, связанное с возрастом в области нервных окончаний синаптофизина на двигателе концевых пластинок, как у мышей прогресс в пожилом возрасте. Те же методы показали, что добровольным мероприятием можно в значительной степени предотвратить уменьшение площади 22 терминала пресинаптических нервных, в соответствии с предыдущей работой со стороны других групп 4. Потеря нервно-мышечных соединений также происходит в модели SOD1G93A мыши бокового амиотрофического склероза 9,23.
Исследования, упомянутые выше, показывают, что многие факторы здоровья могут привести к сокращению в области либо до или после синаптических специализаций НМС. Это может привести к нарушению синаптической удовольствияие или может предвещать полную потерю нервно-мышечного соединения. Три протокола описан, что позволит количественно оценивать площади и плотности синаптических специализаций. Цель первого протокола является создание практического и воспроизводимое измерение области до и после синаптических специализации и их выравнивание при НМС млекопитающих, с помощью флуоресцентной микроскопии. Двумерные максимальной проекцией конфокальной изображения и анализа изображений с NIH ImageJ используется для обнаружения изменений в области синаптофизин окрашивания (синаптические пузырьки), постсинаптических АХР и синаптической области перекрытия. Конфокальные параметры визуализации (усиление и смещение уровня) оптимизированы для каждого НМС так, чтобы максимизировать визуальную информацию, используемую для разглядеть площадь синаптической специализации. Нервно ошибка может также возникнуть в результате изменений в плотности постсинаптических AChR и / или других синаптических белков. Второй протокол может быть применен для обнаружения изменений в относительной плотности постсинаптических белков, такихкак мускус, rapsyn, Dystroglycan, фосфорилированного Src киназы и фосфорилирования AChR 18,21.
В миастения, снижение плотности АЧР в постсинаптической мембране Непосредственной причиной синаптической недостаточности и мышечной слабости. Третий протокол описывает метод флуоресцентного резонанса Передача энергии (FRET), чтобы оценить изменения в непосредственной близости от соседних АХР в постсинаптических мембранах 14,15. Этот метод определяет передачу энергии между соседними АХР, меченных флуоресцентной-α-бунгаротоксина (BGT). FRET происходит только тогда, когда флуоресцентные донорные и акцепторные зонды менее 10 нм друг от друга. Это может выявить (субмикроскопические) изменения в герметичности АХР упаковки, которые могут непосредственно связанные с амплитудой синаптических потенциалов.
Эти три протокола, совершенствуется на протяжении последнего десятилетия, обеспечивают дополнительные меры NMJ целостности в последовательной и воспроизводимой образом. Использование стандартных протоколов Ай параметры должны облегчить сравнение эффектов генов и окружающей среды вмешательств на млекопитающих НМС.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
ПРИМЕЧАНИЕ: Проектирование, проведение и отчетность экспериментов на животных следует учитывать нынешних руководящих принципов 24. Такая работа должна быть заранее одобрены местными властями защиты животных (в нашем случае комитет по этике животных из Университета Сиднея).
1. Эвтаназия животного и мышц Вскрытие
- Передача мышь от удерживающей комнате с отдельной комнате, где он эвтаназии путем внутрибрюшинной инъекции пентобарбитона раствора (30 мг / кг) с использованием метода обработки мыши список по Shimizu 25. Поместите мышь обратно в клетку.
- После того, как дыхание мыши была остановлена на более чем 1 мин, проверить футов снятие рефлекс, осторожно сжимая ноги, и роговицы рефлекс, слегка чистки роговицы. Только тогда, когда рефлекторные реакции отсутствуют можете мышь быть подготовлены для вскрытия.
- Обратитесь атлас грызунов анатомии, таких как Чиассон 26 и / или обратиться за помощью к experienCED анатом, прежде чем пытаться рассечение мышцы интерес. В каждом случае удаления волос из вышележащих кожи с помощью небольшого электробритву, прежде чем открывать кожу, чтобы выставить мышцы.
ПРИМЕЧАНИЕ: рассечение будет отличаться для каждого анатомически отдельной мышцы. - Использование тупых щипцы освободить мышцы от вышележащих оболочек и окружающих тканей. Возьмите и сократить дистального сухожилия, чтобы отделить мышцу от его введения.
- Аккуратно дразнить и отрезать мышцы, свободной от окружающих тканей обратно к своему источнику. Вкратце место недавно расчлененный мышцы в 0,1 М фосфатном растворе буферный солевой раствор (PBS) или раствор Рингера перед дальнейшей обработкой.
2. Подготовка мышц для Cryosectioning
Примечание: Оптимальное структурное сохранение может быть достигнуто за счет целого животного перфузии, как описано ранее 27 или погружением крепление (для небольших мышц), как описано в необязательной стадии 2.1. Тем не менее,4% параформальдегида фиксации может привести к ухудшению последующим окрашиванием со многими антител зондов и с люминесцентными-BGT. Глутаральдегид частности, следует избегать. Если мышцы не должны быть фиксированной, они должны быть немедленно замораживали (перейти к 2,3).
- Дополнительное крепление погружения: Pin мышцы воска в чашку Петри в длину покоя. Накройте мышцы с 2% вес / объем параформальдегида (свеже, растворенного в PBS) в течение 2 ч при комнатной температуре. Промыть 3 с изменениями PBS в течение 30 минут (3 х 10 мин), затем заменить PBS с 30% вес / объем сахарозы в PBS и инкубировали O / N при 4 ° С.
- Сделать формы ('') лодки заранее спинка 2 см х 1,5 см кусочки алюминиевой фольги, как показано на фиг.1. Поместите кусок нитроцеллюлозную мембрану в нижней части шлюпки. Аккуратно влить криостата вложения матрицу (Материалы таблицу) в лодке на глубину 2 мм, заботясь, чтобы избежать воздушных пузырей. Поместите мышцы в лодку, совместив его с шариковой ручкой линий нанитроцеллюлозы. Добавить еще вложение матрицу так, чтобы полностью покрыть мышцы (рисунок 1).
- Pre-этикеток полипропиленовые трубы с несмываемым маркером. Поместите каплю воды в каждую пробирку и охладить трубку в жидком азоте.
ПРИМЕЧАНИЕ: замороженные капли воды поддерживает давление паров и предотвращает высыхание при длительном -80 ° C Температура хранения - Использование маску, толстые защитные перчатки и большой пару тупых щипцы, частично снизить небольшой металлический стакан (диаметр 3 см, 8 см глубиной), содержащий 2 см Глубина изопентана в контейнер с жидким азотом в течение 30 сек. Снимите стакан и поместить его на скамейке наверху. Использование меньшего пару тупых щипцы разместить форму, содержащую мышцы и вложение матрицу в охлажденной изопентана. Будьте осторожны, чтобы избежать смешивания с жидким азотом с изопентана.
- Разрешить 2 мин для блока заморозить перед использованием тупых щипцы, чтобы поднять замороженного блока, и запечатать его в правильном прэлектронной помечены и предварительно охлажденной трубки (этап 2.3).
- Хранить трубы временно в жидком азоте до передачи -80 ° С. Записывать все образцы в таблице содержания морозильную камеру.
3. Cryosectioning и флуоресценции окрашивания для ванной изображений лиц НМС
- Удаляйте алюминиевые пресс-формы. В -20 ° C криостата камеры прикрепить замороженный блок криостата патрона таким образом, чтобы сократить 20 мкм криосрезы параллельно длинной оси мышечных волокон (фиг.1). Возьмите в разделах, посвященных поли-L-лизина или покрытые желатином предметные стекла.
- ПРИМЕЧАНИЕ: Пропустите этот шаг, если ткань фиксируется до замораживания. После того как 30 мин в течение участки высохнуть на слайдах, исправить их, помещая капли 2% параформальдегидом в PBS в течение каждого раздела в течение 15 мин при комнатной температуре.
- Промыть слайды 3 х 10 мин в PBS в банку Коплин, а затем погрузить слайдов в PBS, содержащем 0,1 М глицин в течение 30 мин, чтобы блокировать остаточные альдегидных групп.
- Промыть слайдов в течение 10 мин в PBS, а затем погружают в метаноле (охлажденного до -20 ° С) в течение 7 минут. Этот шаг пермеабилизации является обычной частью двойной маркировки с люминесцентными BGT и анти-синаптофизина но это может отрицательно сказаться на иммуноокрашивания для некоторых других белков.
- Вымойте слайды 2 х 10 мин в PBS затем поместить каждый слайд в стабильной и сравняли влажной камере. Сразу охватывать каждую секцию с 20 мкл блокирующего раствора (0,2% Triton X-100, 2% бычьего сывороточного альбумина (БСА) в PBS) в течение 1 ч при комнатной температуре. Разделы не должно быть позволено высохнуть на любом этапе процесса иммуноокрашивания.
- Провести первичный инкубации: взять один слайд в то время, осторожно удалите излишки раствора блокирующий из более чем каждой секции и заменить его 20 мкл кроличьего анти-синаптофизина (разбавленный 1: 200 в блокирующем растворе).
- Включить негативного контроля слайд, который будет инкубировали с блокирующим только решение. не Это «нет первичной контрольное антитело"Необходимо в каждом Иммуноокрашивание перспективе.
- Забота, что первичное антитело остается на месте в течение каждого раздела, закройте влажной камере и инкубировать в течение 1-2 дней при 4 ° С.
- Проверьте каждый раздел, чтобы подтвердить, что первичное антитело остается на месте. Используйте пипетку Пастера осторожно промыть каждый слайд с PBS и поместить его в банку Коплин. Вымойте все слайды 3 х 10 мин в PBS.
- Провести вторичную инкубацию. Принимая один слайд в то время, осторожно удалите излишки PBS, положите его в увлажненной камере и охватывают каждую секцию с 20 мкл смеси, содержащей FITC-конъюгированного осел анти-кролика IgG и BGT конъюгированный с Тетраметил родамин или еще один красный флуорофор (TRITC- / redBGT; 5 мкг / мл) разводили в блокирующем растворе. Инкубируют при комнатной температуре в течение 2 часов.
- Вымойте слайды 3 х 10 мин в PBS в Коплин банки.
- Принимая один слайд в то время, осторожно удалите излишки PBS и смонтировать с покровным использованием минимального объема, глицерол основе, нарастающий сопротивление монтажа среды. Печать края покровных с четким лака для ногтей. Позвольте ему высохнуть трудно.
- Храните слайды в темноте при 4 ° С на срок до одной недели или при -20 ° С для длительного хранения (до нескольких месяцев).
4. Объективное Отбор проб и анфас Визуализация Motor концевых пластин
- Blind слайды, маркируя каждый слайд со случайным кодовым номером, который остается известным только второй исследователя (которые не участвовали в анализе). В результате оператор остается слепым к группам лечения до количественное определение NMJ параметров не будет завершена для всех образцов.
- Поместите слайд на предметный столик микроскопа и просматривать его под широким освещением поля с множеством TRITC фильтра (63X масло 1,3 NA объективной). Перемещение постепенно (поле за полем) слева направо и обратно, пока не лобовая плита появляется в поле (рис 2а).
ПРИМЕЧАНИЕ: Отбор проб критерий: Каждый AChR окрашенных структура, которая является относительноплоской и сталкивается с целью (то есть, проходит <15 м в Z-измерении) считается концевой шайбы и отображены для анализа (полумесяца окрашивания AChR представляют поперечные сечения через концевых шайб и, следовательно, исключены). - С Регулируемая щель от 1,0 Эри устройства и низкой мощности лазера оптимизации усиления и смещения уровня для TRITC / красный-БГТ (532 нм) лазера на концевой шайбы, которые должны быть отображены. Следующая оптимизировать FITC / синаптофизин флуоресценции с помощью 488 нм лазера на. Сбор Z-стек концевой шайбы с интервалом 0,7 мкм между каждым оптическим среза. Сохранение изображений с именем файла, которое включает в себя дату изображений сессии, кодовое имя слайда и номер концевой шайбы.
ПРИМЕЧАНИЕ: Сканирование с использованием 488 нм и 532 нм лазеров (FITC и TRITC) должны быть собраны последовательно (не одновременно), чтобы избежать загрязнения канала FITC флуоресценции, от красного флюорофора и наоборот (проступание). - Повторите отбора пробй изображений шагов 4,2-4,3 до 20 торцевые пластины собраны из слайд / образец.
- Переход к следующей кодированной слайд и повторите 4,2-4,4. Повторите эту процедуру для каждого из кодируемых слайдов.
- Соберите несколько изображений концевых пластин из управления слайд (без первичной управления антитела), используя настройки конфокальной, которые были найдены оптимальные для экспериментальных слайдов (флуоресценции канал FITC должен появиться темные).
- В конце конфокальной передачи сеанса файлы изображений на другой компьютер и обратно до оригинальных файлов с внешнего жесткого диска или сервера.
5. Измерение площади синаптические специализации в фас изображений
- Используйте NIH ImageJ бесплатно (http://imagej.nih.gov/ij/) подготовить максимальный выступ (MIP) изображения от каждого Z-стека. Сохранить их в качестве TIFF файлов (рис 2А и В). Имена файлов должны содержать дату изображение сеанса, примеры кода, номер концевой шайбы и флуоресцентный канал (например, 060414_5723_7_FITC.tiff).
- Откройте изображение Z-проекции в ImageJ. Выберите изображение канала рецептора ацетилхолина (рис 3а) и выберите: Image> Type> 8-бит, чтобы преобразовать 24-битовый RGB цветного изображения на три 8-битных изображений в градациях серого на экране.
- Использование Polygon Tool ImageJ нарисуйте контур вокруг торцевой пластины интереса к redBGT окрашенных (АЧР) канал так, чтобы включать в себя все видимые окрашенные регионы конкретного индивидуального концевой пластинки, в то время исключая окрашивание, что не происходит из торцевой пластины интереса ( Фигура 3С).
- Применить минимальный порог интенсивности к изображению, выбрав: Изображение> Коррекция> Порог (рис 3E и связанные с ними скриншоты ImageJ).
- Отрегулируйте пороговый уровень так, чтобы изолировать AChR окрашенных частей, исключая окружающий фоновый сигнал, как суб-порог (рис 3Е). Откройте второе дыханиеОй с оригинальным (однотонных) снимка сразу рядом с окном для сравнения, чтобы облегчить решение о пороговом значении. Запишите пороговое значение для дальнейшего использования в колокализации анализа.
- Сохраняя полигон контур вокруг концевой пластинки выберите: Анализ> Анализ частиц. В всплывающем меню укажите диапазон размеров, как: 50 до бесконечности пикселей (это исключает крошечные артефакты, связанные с электрическими помехами в ФЭУ).
- Анализ Частицы команда создает окно со списком дискретных выше пороговых областей и их значения интенсивности флуоресценции пронумерованы, как они появляются в бинарном изображении (рис 3G и связанного с экрана ImageJ). Скопируйте эти данные в меченого таблицы.
- Измерьте Общая площадь концевой шайбы (область внутри полигона), выбрав: Анализ> Измерить. Это дает общую площадь концевой шайбы. Скопируйте и вставьте данные для AChR районов и интенсивности втаблица убедившись, что метка колонки надлежащим образом, строки будут использоваться для отдельных торцевых пластин для конкретных слайдов.
- Переключитесь на анти-синаптофизин канала флуоресценции и повторите шаги 5,1 - 5,5, а для канала FITC (3В, D и F). Цель состоит в том, чтобы настроить порог так, что он создает бинарный образ, который, насколько это возможно, соответствует границам окрашивания, как воспринимается глазом. Запишите пороговое значение.
- Измерьте площадь перекрытия, применяя следующие действия: Откройте исходный файл, содержащий два канала изображения и разделить его на два отдельных изображения, выбрав: Image> Стеки> Стек изображений.
- Используя плагин колокализационные (загрузка и установка с веб-страницы ImageJ) Выберите: Pluggin> колокализационные и входные пороговые значения ранее записанную для каналов AChR и нервных в соответствующем канале запрос Бвол. Это даст перекрытие изображения в белых пикселей (рис 3Н и связанных с ними скриншотов ImageJ).
- Преобразование созданную перекрытия изображения в черно-белом формате и применить порог максимального значения. Максимальный порог будет выбирать только белые пиксели, соответствующие области перекрытия двух предыдущих каналов. Запись в таблице полученное значение площадь "колокализации", которая представляет собой область перекрытия в пикселях.
- Подготовьте таблицу выборки данных с помощью, рассчитать и построить стандартные отклонения и стандартные ошибки, как гистограммы или диаграммы рассеивания 20,22. Следует отметить, что значение N в основном представляет собой количество мышей в группе выборки для статистических целей.
- Участок концевых пластин АЧР областях, как диаграммы рассеивания или частотных гистограмм, чтобы определить, является ли данные нормально распределены до статистических испытаний (рисунок 6).
6. Относительная ОкрашиваниеИнтенсивность сравнивали с помощью поперечных оптических Разделы
ПРИМЕЧАНИЕ: Для этого процесса протокола все образцы мышц вместе, и изображение на одной сессии конфокальной. При планировании эксперимента позволяют до 30 мин времени съемки на пробу мышц.
- Вырезать 15 мкм криосрезы поперек длинной оси мышечных волокон и собирать на слайдах, как описано на шаге 3.1.
- Провести флуоресцентного окрашивающего как описано в шагах 3.2-3.13.
- Код окрашенные слайды, так что изображения и анализы, проведенные с оператором слепых в группу лечения, как описано в шаге 4.1.
- Использование флуоресценции цели 40X (NA 0,75) краткий обзор раздела из каждого слайда, чтобы определить единого усиления и смещения настройки уровня для AChR, что будут пригодны для всех концевых пластин во всех образцов слайдов. Яркая Концевая пластина должна затем быть чуть ниже 256 серый по шкале. Эта оптимизация должно быть сделано отдельно для второго fluorescencе канала (собранные подряд). Запишите фиксированного усиления и смещения настройки уровня и не изменяют их в течение сессии изображений.
- Собирают изображения стандартной слайд флуоресценции (например, не-отбеливающие флуоресцентные шарики), с теми же параметрами, в начале и в конце конфокальной сессии для обнаружения любого возможного колебания интенсивности лазерного.
- Используйте канал AChR для сканирования слайдов постепенно, чтобы найти торцевых пластин.
- Фокус, чтобы найти единый оптической части плоскости в каждом поле микроскопа, который содержит наибольшее количество AChR окрашенных концевых пластинок.
- Сканирование этот единственный оптической части в два раза и сохранить усредненный образ (рис 4G).
- Сохраняя ту же фокальной плоскости переключатель на второй канал флуоресценции (белок интерес) и собрать изображение, как на этапе 6.8. Сохраните файл изображения, в том числе в имени файла: Дата визуализации сессии, примеры кода, номер изображения и символ для обозначения флуоресцентного канала.
- Перемещение этап к следующему полю, которое содержит один или несколько концевых пластин и повторите шаг 6.8-6.9. Повторите это, пока в общей сложности 60 торцевых пластин изображаются.
- В конце передачи изображений сессии все файлы на другой компьютер и обратно их.
- Откройте каждый исходный файл изображения и при просмотре канала AChR, выберите: Image> Стеки> Стек к изображениям, чтобы разделить каналы.
- Выберите: Image> Type> 8 бит преобразовать в 8-битные оттенки серого формате на экране. Делайте это в течение двух флуоресцентных каналов.
- Выберите: Image> Стеки> Изображения укладывать. Откройте новую пачку из двух ранее разделенных 8-битных изображений. То можно переключить удобно между двумя каналами флуоресценции в одном окне.
- Используйте инструмент многоугольник сделать линэлектронной плотно вокруг границы AChR окрашивания (фиг 4I).
- Выберите: Анализ> Измерение измерить среднюю интенсивность пикселя для AChR в закрытом помещении (обратите внимание на важность рисования линии сильно). Скопируйте это значение в меченого таблицы.
- Сохранение же многоугольника план (определить область, которая будет измерено), переключитесь на второго флуоресцентного канала (например, рис 4В, D, F) и выберите: Анализ> меры. Это даст среднюю интенсивность окрашивания для белка, представляющего интерес в синаптической области, определенной с помощью окрашивания AChR.
- Выберите область от видимого окрашивания концевой пластинки затем выберите: Анализ> Измерение измерить среднюю интенсивность фоновой флуоресценции. Повторите эту процедуру для другой флуоресценции канала / с и скопировать фоновые значения в таблицу значений флуоресценции.
- Subурочищу средних фоновых значений из значений шайбами, чтобы получить исправленные интенсивности для AChR и интерес белок при каждом концевой пластинки.
- Разделить скорректированные значения яркости концевой шайбы для интересующего белка от исправленной интенсивности флуоресценции БГТ с получением соотношения интенсивности флуоресценции 14,21
7. Сравнение плотности постсинаптической мембраны AChR Использование FRET
ПРИМЕЧАНИЕ: Этот протокол оценивает, в какой степени АХР, плотно упакованными (<10 нм пробелами) в постсинаптической мембране. Точный донорных и акцепторных флуорофор комбинации имеет решающее значение для этой FRET анализа. Имена и детали флуорофоров приведены в таблице материалов. Их спектральные свойства по отношению к ладу обсуждаются в наших предыдущих работах 14,15.
- Подготовка фиксированные поперечные криосрезы, как описано в разделе 6.1. Все группы образцов должны быть обработаны вместе и изображенияд в том же конфокальной сессии.
- Тщательно перемешайте 2,5 г / мл красного-BGT (FRET донора) с 10 мкг / мл до красно-BGT (FRET акцептором) с блокировкой решение в небольшой пластиковой трубки с помощью пипетки вверх и вниз 12 раз. Это 1: 4 молярная смесь максимизирует эффективность FRET 14.
- Поместите каждый слайд в увлажненной камере, тщательно покрыть каждую секцию с каплей (12 мкл) полученной выше смеси и инкубировать в течение 1,5 ч при комнатной температуре.
- Контрольных участках: Обложка небольшое число секций с 2,5 г / мл красно-BGT (единственным донором; меченые управления C1), а также некоторые разделы с 10 мкг / мл далеко красно-BGT (только акцептора; меченые управляет С2). Выдержите эти элементы управления, как на этапе 7.3.
- Вымойте слайды 3 х 10 мин в PBS и смонтировать в основе глицерина, нарастающий сопротивление монтажа среды (см шаг 3,12).
- Выполните выборку концевых пластин, как в шаге 6.7. Флуоресценции от донора и акцептора должны быть полностью со-локализованы на концевых пластин из-за случайного связывания молекул флуоресцентного-BGT,
- Контрольные изображения: Использование цели 40X и низкой мощности лазерного излучения оптимизировать прибыль redBGT и смещение настройки уровня для торцевых пластин от управляющего слайд-C1. Оптимизация далеко redBGT усиления и смещения уровней для торцевых пластин из управления слайд-С2. Подтвердите отсутствие каких-либо флуоресценции проступание.
- Не изменяя мощность лазера, получить или настройки смещения уровня перейти на экспериментальных слайдов и собрать снимков (предварительно photobleach) для обоих флуоресценции каналов.
- Избирательно photobleach в пор-красно-BGT над частью одной концевой шайбы за счет увеличения области сканирования, то сканирование в 10 раз с помощью лазера на 633 нм при 100% мощности. Флуоресценции в области сканирования должна стать тусклым.
- Сброс мощности лазера и увеличить и собирать после отбеливателя изображения на обеих люминесцентных каналов с помощью конфокальной настройки, установленные на 7,7.
- Рассчитать эффективность FRET (E) от процентное увеличение донора (красно-BGT) флуоресценции после photobleach акцептора (FAR-красно-BGT) в соответствии со следующей формулой *:

* Для всех ситуаций, в которых флуоресценция донора возрастает после фотообесцвечивания акцептора.

Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Измерение Synaptic Района сразу после NMJ
Любое оценка области опирается на чертеже границы, чтобы определить степень синаптических специализаций. У здоровых молодых взрослых мышц НМС изображения должны отображаться четко определенные границы как для AChR и синаптофизин окрашивания (рис 2А и В). Интенсивность флуоресценции как для AChR и синаптофизина резко возрастает на границе между пери-синаптической и синаптической части концевой шайбы двигателя (фиг 5А 'и B'). Для таких снимков минимального порога (чуть выше внесинаптического фоновой флуоресценции) будет легко выделить АКПЧ или богатый синаптофизин богатых площадь торцевой пластины (горизонтальная пунктирные линии на (рис 5А »и В ')). У мышей в возрасте, а в некоторых болезненных состояний концевой пластинки окрашивания для AChR может быть менее интенсивным, АЧР кластера края могут выглядеть смазанным иRe может быть выше уровня внесинаптического флуоресценции (рис 5D; 17,22). Флуоресценции окрашивания с нечеткими границами можно ввести погрешность в оценках синаптической области. Во всех ситуациях, цель заключается в выборе порога, который дает бинарный изображение, аналогичное по форме и размерам с AChR- или синаптофизин богатых областях, как они будут выглядеть глаза в оригинале, непрерывного тона изображения. Проведение анализа слепы к группе лечения следует уменьшить риск субъективности на этапе выбора порога (этап 4.1). Иногда слабые или размытые концевых пластин изображения может привести к неоптимальным обработки. Рисунок 2С и D показан пример низкого качества концевой пластинки изображения от здорового 2-месячного мыши. Размытые края и слабый окрашивание синаптофизин может возникнуть в этом случае частичного оттаивания и повторным замерзанием мышцы до cryosectioning. Некоторые разделы здорового молодого (положительный контроль) мышц должны быть срезый обрабатываются параллельно с экспериментальными образцами, чтобы гарантировать, что любое ухудшение проявляется в НМС изображений не из-за проблем с иммунной окраски. Партии изображений взломанных к югу от оптимальной обработки должны быть исключены из анализа.
Для ванной изображений лица г-стека, 15-20 торцевые пластины является разумным размер выборки для оценки синаптических области. Широкое разнообразие в форме и размерам НМС находятся в пределах любого данного мышцы. Диаграммы рассеяния показывают значительный диапазон в АКПЧ-богатой области между концевыми пластинами из передней большеберцовой мышцы в любой отдельной мыши (фиг.6А). Тем не менее, средняя площадь AChR (на основе 15-20 фас шайбами изображений) был похож на семи образцов мышей (~ 200 м 2; рис 6А). Площадь окрашивания синаптофизин концевой пластинки также значительно различались между торцевыми пластинами из данного мышц. Еще раз, с использованием образца размером 15-20 концевых шайб средняя площадь концевой шайбы синаптофизина был СимиLAR среди 7 мышей изучена (~ 170м 2; рис 6В). Частотные гистограммы объединенных данных показал, примерно нормальное распределение для области концевой пластинки AChR и синаптофизина (рис 6A 'и В'). Однако нормальное распределение синаптических областях, не может быть принято в болезненных состояний, таких как миастения 16,20. Это может повлиять на выбор статистического анализа.
В таблице приведены области до и после синаптических специализаций для НМС для здорового 2 месяца (молодой взрослый) Женский C57BL / 6J мышей от предыдущих исследований. Области как до, так и после синаптических специализаций отказался с сидячий старения 22. AChR область была также заметно снижается у мышей, которым вводили IgG из анти-MuSK миастения пациентов 17,21. Миастеничным мышей, обработанных ингибитором холинэстеразы наркотиков, пиридостигмин, отображается на дальнейшее значительное снижение концевой шайбы AChR области 20 </ SUP>.
Относительная интенсивность маркировки Торцевая пластина флуоресценции
Относительная интенсивность иммунофлуоресценции маркировки может выявить изменения в плотности синапсов белок интересов, с возрастом, генотипом и / или болезненного состояния. AChR флуоресценции (красный-БГТ или далеко-красно-БГТ) сначала используется для определения местоположения НМС. Яркость флуоресценции в 8-битном AChR-богатой области затем используется для оценки изменений в концентрации белка, представляющего интерес, по сравнению с контрольными животными. В поперечных срезах концевой шайбы АХР, как правило, появляются в форме полумесяца, но эта форма часто нерегулярно (4А, С, Е, Н). Низкая интенсивность фоновой флуоресценции обычно показывает, является ли патч окрашивания АХР одного торцевую крышку, или два отдельных торцевые пластины, расположенные на соседних мышечных волокон. Многие синаптические белки (такие как rapsyn, мускуса и) ДЗОлокализуется с AChR на концевой пластине (рис 4А - D). Иммунофлуоресценции окрашивание фосфо-специфичных антител также может быть использован, чтобы сравнить эффект экспериментальных вмешательств на состояние фосфорилирования частности постсинаптических мембранных белков 21.
Надежность и воспроизводимость измерения интенсивности флуоресценции в значительной степени зависит от целостности замороженной мышцы и качества иммунной окраски. Мышцы должны быть расчленены и сразу быстро замораживают или параформальдегид фиксированной (в течение нескольких минут после смерти животного), чтобы избежать дегенеративные изменения в НМС. Иммуноокрашивание сильно зависит от качества используемых реагентов и оптимизации протокола окрашивание на специфических антител. Для любой новой партии первичного антитела экспериментов пилот иммуноокрашивания необходимы. Свежесрезанных криосрезы здоровых молодых мышц инкубируют с серийными 2-кратных разведений первичного антитела.Известно, надежность вторичного антитела используют, и результаты сравниваются. Если интересующий белок, как известно, ограничивается НМС то лучше концентрация антитела является то, что дает высокий коэффициент интенсивности флуоресценции по отношению к NMJ, что найдено в внесинаптических частей мышцы (фоновой флуоресценции). Внесинаптического (предположительно неспецифический) интенсивность флуоресценции обычно не должен превышать 15% от интенсивности флуоресценции лобовая плита. Точно так же «нет управления первичное антитело» в разделах (инкубировали только с вторичным антителом) не должны быть темными, подтверждая, что вторичное антитело не связывался неспецифически. Качество различных партий (поликлональные) вторичные антитела может изменяться значительно, так альтернативных партий вторичного антитела должны быть сопоставлены до создания стандартного протокола. Идеально подходит тест на специфичность иммунофлюоресценции включает в себя прямое сравнение секций от мышей дикого типа и секций негативного контроля от мышей тшляпа не хватает белка интересов, (ген нокаутом). Следующие ссылки описывают количественное концевой пластинки флуоресценции окрашивания для мускуса, rapsyn, Dystroglycan, ДЗО и АХР 13,14,18,21.
Небольшие размеры выборки ввести ошибку в оценках относительной интенсивности флуоресценции. Отдельные торцевые пластины значительно варьироваться по интенсивности флуоресценции. Предположительно эта изменчивость между торцевыми пластинами внутри данного мышц отражает разнообразие НМС структуры и случайных различий в оптической части пробы. Тем не менее, увеличение числа концевых пластин, отобранных результатов в более стабильной оценке средней интенсивности флуоресценции (рис 7). Для оценки среднего концевой шайбы интенсивность флуоресценции 40-60 торцевые пластины из каждого образца мышечной должны быть усреднены.
AChR-АЧР FRET
Каждый AChR является пентамер с двумя сайтами связывания для BGT (расположенных по одному на каждой альфа-субъединицы). Связываниекрасно-BGT и далеко-красно-BGT для этих двух сайтов дали бы разделение донор-акцепторного около 9nm 28-30. Таким образом, низкая эффективность FRET можно обнаружить еще до АХР собираться в кластеры 14. Тем не менее, эффективность FRET в мышиных концевых шайб примерно в два раза после рождения, в соответствии с более эффективного включения АХР в плотно упакованной решетки постсинаптической мембраны 14. Использование красных BGT и далеко-красно-BGT в качестве донора рвать и акцептора (соответственно), концевые шайбы 1-2-месячного мышам, средние эффективность FRET, начиная от 20-37% (таблица 2). FRET КПД 20% и более, как полагают, представляют герметичной упаковке в АХР 14. Эффективность Торцевая пластина FRET несколько снизилась после денервации 14, и заметно снижается после инъецировались IgG из анти-мускусный положительного миастения пациентов 18. Эти условия, в которых отдельные АХР менее плотно упакованные в postsynaptic мембрана леса на постсинаптической системы MuSK / rapsyn 31.
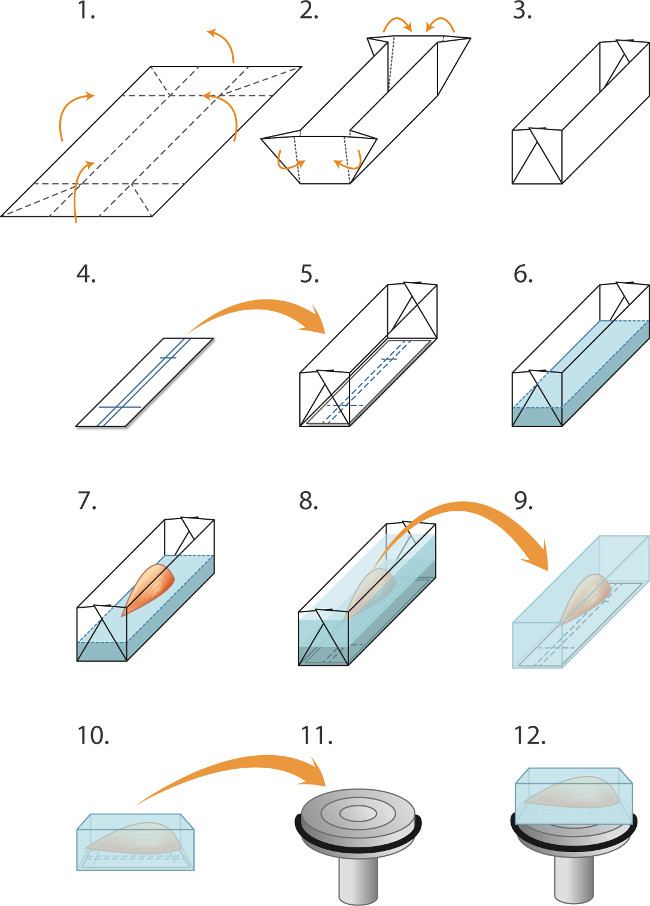
Рисунок 1. Встраивание и морозильные мышцы для cryosectioning (1-5) Подготовка формы ('лодки') перед замораживанием партию мышц:. (1) Алюминиевая фольга разрезается на прямоугольники (2,0 х 3,0 см) и (2). Сложенный как обвинение, чтобы создать формы / лодку (3). (4) Прямоугольники из нитроцеллюлозы иммуноблоттинга бумаги вырезать, чтобы соответствовать в форме и Шариковая ручка используется, чтобы исключить линии для ориентирования мышцы. (5) нитроцеллюлозы прямоугольника помещается в пресс-форму. (6-8) Встраивание и замораживания мышцы: (6) Криостат встраивания матрицы жидкость аккуратно выливают в пресс-форму (в верхней части нитроцеллюлозы) на глубину 2 мм. (7) декоративные щипцы используются для снижения мышцы, его сухожилия на вложения матрицы, в соответствие с nitrocelluпотерять постановления, (8) Дополнительная вложение матрица аккуратно заливается в форму, чтобы покрыть мышцы, стараясь избежать образования пузырьков. Встроенный мышцы затем быстро замораживают, запечатанный в пробирки и хранили при -80 ° С, как описано в тексте. (9-12) Подготовка к cryosectioning: (9) тонким пинцетом используются, чтобы очистить от алюминиевую форму и замороженный блок помещают в -20 ° С криостата камере, (10) Маркировка в нитроцеллюлозу используются для выравнивания мышц параллельно поверхности патрона (11) для продольной срезов (12). Капля жидкости крио-вложение среды используется для крепления блока к охлажденной патрона. Тупые щипцы используются для манипулирования замороженного мышц блок. Чтобы получить поперечные срезы, блок вместо установлен так, чтобы мышца перпендикулярна поверхности патрона (не показан).

F2. igure анфас изображения НМС от передней большеберцовой мышцы 2 месячных самок мышей C57Bl6J. Свежий мышцы быстро замораживали и секции закреплены на слайде, как описано на этапе 3.2. Были получены, как описано в первом протоколе Проекция максимальной интенсивности изображения Z-стеков. Окрашивание (А) красно-БГТ показывает один двигатель концевой шайбы, состоящий из двух наборов AChR-богатых первичных желобов постсинаптических (EN вид спереди). (Б) синаптофизина окрашивания FITC-конъюгированного вторичного антитела показывает пресинаптического нервные окончания, занимая основные синаптические желоба. Часть предварительно терминала аксона также видна в верхней части панели. (С & D) Пример плохого качества изображения NMJ от молодых здоровых мышей. Окрашивание слаб и границы до и после синаптических специализаций размыты. Это было связано с недостатками в тОн обработка ткани и / или недостаточной времени, отведенного на инкубации первичных антител. Бар Шкала в панели D представляет 10 мкм. Пожалуйста, нажмите здесь, чтобы увидеть увеличенную версию этой цифры.

Рисунок 3. Шаги в обработке фас НМС изображений (как указано в протоколе 1). (& B) Оригинальные непрерывный сигнал MIP изображения показывают красно-BGT флуоресценции, раскрывая АЧР и зеленый иммунофлюоресценции синаптофизина соответственно. (C) AChR окрашивание после преобразования в 8-битной оттенков серого изображения и использования Polygon Tool, чтобы очертить концевой шайбы (тонкая желтая линия). (D) Торцевая пластина линия границы перенесена на зеленой синаптофизин изображения. (E) Application минимальной пороговой интенсивности команду, чтобы создать бинарный файл, который изолирует сверхпороговой красно-BGT (AChR) флуоресценции. Последовательность ImageJ скриншотов пользовательского интерфейса отображается с левой стороны изображения. (F) Двоичный синаптофизина изображение после применения отдельного порога. (G) Определение дискретного надпороговых AChR-богатых доменов в концевой шайбы с применением частиц команды Analyze в двоичном красно-БГТ изображения. Соответствующие ImageJ (слева от панели (G) показывает входные данные, необходимые. Минимальный размер надпороговых районах пикселей, необходимых должны быть введены. (H) Выявление областей перекрытия двоичной синаптофизина и AChR изображений. Перекрытие представлена белых пикселей . ImageJ пользовательский интерфейс Скриншоты (ниже панели H) показывают шаги, направленные на достижение бинарной перекрытия изображения. Избранный минимальный порог интенсивности значения для каждого флуоресценции сhannel должны быть введены в окне "колокализационные. Масштабная линейка представляет 10 мкм. Пожалуйста, нажмите здесь, чтобы увидеть увеличенную версию этой цифры.

Рисунок 4. Примеры поперечных оптических секций, используемых для сравнения относительных интенсивностей флуоресцентных (Протокол 2). Мышца быстро замораживали и секции, закрепленные на слайде, как описано на этапе 3.2. (A & B) одного концевой шайбы дважды меченное пор Красное-БГТ (АКПЧ; показанный здесь в синем псевдоцвете) и FITC-анти-rapsyn иллюстрирует совместную локализацию этих двух взаимодействующих белков в постсинаптической мембране (C & D) два концевых шайб на соседних мышечных волокон отображения совместно локализованными AC. ч и мускуса. (E & F) лобовая плита двойной меткой для AChR и -dystroglycan (-dg). -DG Простирается вплоть вокруг мышечных волокон периметру, но обогащается в концевой пластинке (бар шкалы в F, для панелей AF: 25 мкм) (G - I) выделение в торцевую крышку для измерения интенсивности (G) на обычном поле микроскопа,.. содержащий три далеко-красно-BGT-окрашенных концевых пластин (шкала бар в панели G: 40 мкм (H) увеличенное изображение коробочной концевой пластинки (I) же лобовая плита преобразуется в 8-битном черно-белое изображение и по форме, используя полигон.. . инструмент ImageJ (тонкая желтая линия) Средняя интенсивность флуоресценции измеряется в пределах этой границы. (масштабная линейка для H & I: 10 мкм) Пожалуйста, нажмите здесь, чтобы увидеть увеличенную версию этой цифры.

Рисунок 5. Влияние качества изображения при оценке синаптической области. (& B) Высокое качество фас изображений здорового NMJ из 2-месячного мыши смотреть на красно-BGT и анти-синаптофизин флуоресценции каналов. ('& B ') Интенсивность флуоресценции профили, соответствующий линии, проведенной через концевой шайбы на А и В соответственно. Горизонтальная красная пунктирная линия показывает минимальный порог, используемый для создания бинарного изображения. (C & D) Торцевая пластина с пожилой мыши. Торцевая пластина окрашивание синаптофизина целом является менее интенсивным. (С 'и D') профили интенсивности показывают высокий уровень внесинаптического (базового) флуоресценции колебания в синаптофизина (FITC) канала (назадземля), который влияет на определение подходящего порога. Многое из этого широкого спектра тканей аутофлуоресценция. Шкала бары представляют 10 мкм. Пожалуйста, нажмите здесь, чтобы увидеть увеличенную версию этой цифры.

Рисунок 6. Изменчивость синаптических областях между НМС внутри мышцы, а также между мышами. (А) диаграммы рассеяния показано общее AChR-богатый участок концевых шайб из передней большеберцовой мышцы семи наивных 2 мес старых самок C57BL / 6J мышей, полученных автором NT. Каждый символ представляет собой торцевую крышку. Каждый столбик представляет собой среднее ± стандартное отклонение для концевых шайб, отобранных от одной мыши. (B) диаграммы рассеяния, показывающий синаптофизина богатых область для одних и тех же концевых шайб. (А ') (В ') с распределением частот для синаптофизин богатых области концевых пластинок (объединенные данные). Пожалуйста, нажмите здесь, чтобы увидеть увеличенную версию этой цифры.

Рисунок 7. Влияние размера выборки при оценке интенсивности флуоресценции лобовая плита. Поперечные оптические срезы используются для измерения интенсивности флуоресценции (в произвольных единицах) в 40 60 торцевых пластин из передней большеберцовой мышцы здорового 2-месячного мыши. Совокупные средние приведены по отношению к количеству концевых шайб, включенных в среднем. (А) Торцевая пластина красно-БГТ интенсивность флуоресценции получено автора AV. (В) анти-synaptophysiп интенсивность иммунофлуоресценции для тех же концевых пластин, как в панели A. Интенсивность (C) Торцевая пластина далеко-красно-BGT флуоресценции получены из второго образца мышечной По автору NG. (D) Anti-rapsyn интенсивность иммунофлуоресценции из тех же концевых пластин, как в панели C.
| Исследование | Мышца | Количество мышей | AChR область | Синаптофизин область | Перекрытие область |
| значит ± SD (мкм2) | значит ± SD (мкм2) | значит ± SD (мкм2) | |||
| Morsch и др. (2012) 1 | икроножной | 3 | 181 ± 7 | 163 ± 24 | |
| диафрагма | 3 | 130 ± 41 | 102 ± 21 | 76 ± 26 | |
| Morsch и др. (2013) 1 | передней большеберцовой | 3 | 166 ± 26 | 117 ± 21 | й |
| Чен и др. (2013) 2 | передней большеберцовой | 4 | 226 ± 18 | 150 ± 44 | 110 ± 34 |
| Це (не опубликовано) 2 | передней большеберцовой | 7 | 213 ± 27 | 157 ± 23 | 111 ± 11 |
| 1 распечатанных на Zeiss LSM 510 Мета микроскопом, но с фиксированным коэффициентом усиления и смещения уровней. ВЫДЕЛЕНИЕ и синаптические участки измерения используется Metamorph Softwarе ММ. | |||||
| 2 распечатанных на микроскопе Leica DM IRE2 и проанализированы в соответствии с существующей протокола по указанному первого автора, слепой к группе лечения. | |||||
| й не определена. | |||||
Таблица 1. Synaptic области для НМС в 2-месячный женщин C57Bl / 6J (здоровых)
| Исследование | Мышца | FRET эффективность (%) * | Диапазон (%) |
| (Средний ± SEM) | |||
| Brockhausen и др. (2008) | передней большеберцовой | 24 ± 1 | не доступно |
| Коул и др. 2010 | передней большеберцовой | 26 ± 1 | 22 - 30 |
| Morsch (не опубликовано) | диафрагма | 37 ± 1 | 24 - 47 |
| Газанфари (не опубликовано) | передней большеберцовой | 30 ± 1 | 20 - 45 |
| * FRET между красно-BGT и далеко красно-BGT (Ферстер радиуса для FRET пар = 51A (Life Technologies). | |||
| Данные НС не доступно | |||
Таблица 2. Эффективность для AChR FRET из молодых взрослых мышей C57Bl6J мышей
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Протоколы, описанные здесь, позволили нам надежно измерять и количественной оценки изменений в свойствах НМС по целому ряду условий, в том числе нормального старения и болезненных состояний. Методы, описанные в фас НМС изображения позволит исследователям сравнить площадь до и постсинаптических специализаций и область синаптической перекрытия / выравнивания. Для сравнения относительной интенсивности до и постсинаптических белков Второй протокол, который использует поперечных оптических секций, является предпочтительным. Третий протокол конкретно проверяет на изменения в непосредственной близости от упаковки АХР в постсинаптической мембране.
Управления Специфика имеют жизненно важное значение в иммунофлюоресценции микроскопии. При использовании любого первичного антитела для непрямой иммунофлуоресценции необходимо сначала убедиться, что она специфически связывается с его белком-мишенью в секциях мышц. Различные виды обработки ткани и фиксации может дифференцированно изменяют специфичностьантитела. Важно, чтобы подтвердить, что иммунофлюоресценции окрашивание (скажем для rapsyn) действительно концентрируют с AChR на моторной концевой шайбы. Отрицательные контрольные участки также должны быть проверены, чтобы гарантировать, что антитело является специфическим связывания. Например, подходящим для отрицательного контроля rapsyn иммунофлуоресценции бы участки из rapsyn - / - мышей. Они не должны иметь концевой шайбы окрашивание анти-rapsyn. Неспецифическое флуоресценции может также возникать из эндогенных люминесцентных химических веществ в ткани (флуоресценции) или из неспецифического связывания с флуоресцентного конъюгата вторичных антител. Такое флуоресценции часто ухудшается альдегид фиксации. Кроме того, TRITC-БГТ окрашивание концевых шайб иногда могут быть обнаружены в флуоресценции FITC канал, и это люминесцентные проступание может путать с конкретным FITC иммунофлюоресценции. Чтобы защититься от последних трех формах неспецифического флуоресценции, каждая партия слайдов, которые окрашивали должна включать в себя некоторые 'Nконтрольное антитело о первичной 'разделы (этапы 3.7 и 4.6). Изображения концевых пластин из этих контрольных участках должны быть по сравнению с теми из экспериментальных слайдов для того, чтобы косвенного иммунофлуоресцентного окрашивания НМС действительно отражает связывание первого антитела.
Поперечный конфокальной участки являются особенно полезными для оценки различий в относительной интенсивности иммунного окрашивания в синапсах. В поперечных срезах конфокальных легче судить точную совместной локализации синаптических белков. В форме полумесяца профиль Концевая пластина представляет собой лишь пример сквозной НМС в вопросе. Тем не менее, фон (внесинаптического) флуоресценции, как правило, ниже по сравнению с фас г-проекции изображений. Таким образом, это может быть легче различать «настоящую» (специфических) иммуноокрашивания и установить коэффициент усиления фиксированной конфокальной и смещения с помощью поперечных оптических срезов 13-15,18. Например, в модели мыши миастения (WHERе лобовая плита AChR окрашивание заметно снижается) концевые шайбы были четко разграничены в поперечных оптических срезов 18,21. Различия в средней интенсивности флуоресценции на НМС могут отражать измененную плотность белка-мишени в пределах синаптической специализации. Предостережение в том, что в некоторых ситуациях, структурные изменения в белке-мишени или окклюзии связывания антител соседними белков может объяснить измененный интенсивность окрашивания.
Конструкция экспериментов требует внимания. Во многих случаях эксперимент будет стремиться проверить влияние трансгенного, генной нокдаун или болезненного состояния от размера НМС. Экспериментальный образец группа может быть по сравнению с здоровым молодым (дикого типа) мышей того же пола и генетического фона. Исходные значения для области концевой шайбы синаптофизин, AChR и синаптической перекрытия в течение нескольких мышц даны в таблице 1. Пример размер будет зависеть от степени животного-в-животного различия внутри групп лечения и величина эффекта (разница в средствах для экспериментальной против контрольной группы в стандартное отклонение). Когда анализ ограничивается изображений хорошего качества достаточная степень согласованности был найден в образец средства для шайбами районах среди здоровых 2-месячных самок C57Bl6J мышей (рис 6A и В). Таким образом, можно было продемонстрировать значительное снижение синаптической области 30-40% в мышах, инъецированных IgG из анти-мускусный положительного миастения пациентов, по сравнению с контрольной группой с размером образца из трех мышей 17,20,32. Пожилые мышей отображается больший разброс животных к животному в параметрах шайбами, чем молодых мышей 22. Следовательно эксперименты с участием мышей в возрасте может потребовать большего объема выборки.
Если основной задачей является измерение размер фас концевой пластинки затем усиления и смещения настройки уровня должны быть оптимизированы для каждого отдельного НМС. Individual НМС могут значительно различаться в яркости AChR и окрашивания синаптофизин, особенно когда болезненные состояния рассматриваются. Кроме того, интенсивность дополнительной-синаптических (неспецифической) флуоресценции часто выше и более переменных в мышцах стареющих животных, по сравнению с теми здоровых молодых животных (фиг 5С и D). 1 256 серо-шкала должна быть в полной мере так, чтобы максимизировать тонального информацию, которая будет сохранена в последние изображений. Это будет включать регулировки усиления и смещения уровней для каждого НМС, для которых Z-стек, чтобы быть собраны. Рисунок 5D показан пример NMJ изображения, где тональная информация может иметь решающее значение в определении границ области до и после синаптические специализации.
Измерения синаптических областях может быть применен к различным препаратов мышечных и экспериментов. Большинство наших измерений синаптических областях использовали продольные криосрезы от оснастки замороженные мышцы. Замораживание мышцы до фиксации содержит антигенность в широком диапазоне белков. При совместимы с антигеном, фиксацией параформальдегидом и сахарозы инфильтрации до cryosectioning (этап 2.1) может обеспечить лучшую сохранность структуры NMJ. Оптимальная структурно сохранение может быть получено путем сердечной перфузии с параформальдегидом. Артефакты замораживания и секционирования может быть полностью избежать, маркировки концевых пластин на поверхности неповрежденных мышечных и обработки изображений НМС по брошюр дразнили из плотной мышцы 21. Независимо от подготовки, процедуры отбора проб, изображений и области количественного остаются неизменными (протокол шаги 4-5). Последовательное применение слепых проб, анализа изображений и протоколов (с использованием различных операторов, различные образцы мышей и разное время), может привести к довольно воспроизводимых средних значений (ср Cheng и др., И TSE результаты в таблице 1).
"> Концевых шайб были описаны как стать" фрагментарный "в различных болезненных состояний. Например, в процессе старения мышцы мыши, спорадический дегенерацию мышечного волокна (с последующим его регенерации) в результате ремоделирования кренделя, как концевой шайбы AChR доски образуют несколько небольших кластеров AChR 6. У мышей, которым вводили IgG из анти-MuSK миастения пациентов, фрагментация концевой пластинки была довольно разные. лобовая плита AChR кренделя в основном разошлись, оставив созвездии маленьких (<4 м 2) микроагрегатов АЧР " "20,21. Эти два примера подчеркивают необходимость сравнить распределения по размерам для AChR кластеров на торцевых пластин контроля по сравнению с экспериментальными животными 21.Другие методы были зарегистрированы для оценки синаптической области или окрашивания интенсивность в НМС. Моторные торцевые пластины могут быть иногда складывается так, что двумерные г-проекции изображения, используемые здесь, возможно, недооценивают синaptic области. Трехмерные конфокальной реконструкции может обеспечить более точные меры, если абсолютное синаптической области должны быть определены 33. Ключевым преимуществом протокола Z-проекции, описанной здесь, однако, является его относительная простота, которая позволила большое количество концевых шайб измеряется от нескольких группах лечения и надежной идентификации потенциальных изменений. Протокол для сравнения интенсивности окрашивания концевой шайбы может быть адаптирован для изучения изменений в уровнях различных синаптических белков. Метод ограничен, однако, в соответствии с требованием, что все образцы обрабатываются для иммунофлуоресценции то изображаемого в тот же конфокальной сессии. Недавнее исследование, проведенное Ямпольского и др. 5 описан способ для измерения концевой шайбы AChR плотность, которая может помочь преодолеть это ограничение. В этом исследовании, те же поля концевых пластин были обследованы в нескольких различных настроек мощности лазера. Наклон отношения между мощностью лазера и rhodamiИнтенсивность флуоресценции пе-BGT была использована для оценки относительных изменений в плотности АХР на концевых пластин в различных мышей 5. Этот метод может быть полезным для сравнения AChR интенсивность в образцах отображенных в различные моменты времени в течение длительного исследования.
AChR-АЧР FRET обеспечивает конкретную и дополнительную информацию об организации шайбами АХР. Электронно-микроскопическое авторадиография с помощью 125. I-α-BGT показал АХР быть плотно упакованы с плоской плотности 10 4 м -2 непосредственно под каждого пресинаптической сайте высвобождения трансмиттера, в то время как прилегающие мембранные infoldings содержать гораздо меньшие плотности AChR 34. AChR-АЧР FRET позволяет сравнительно легко обнаружить (SUB-микроскопические) изменения в АХР упаковки. Снижение эффективности FRET отражает субмикроскопическом перераспределение АХР в постсинаптической мембране, которые не могут быть обнаружены по изменению средней интенсивности флуоресценции BGT. Несколько Fсубъекты могут привести к изменению эффективности FRET. Они включают в себя расстояние донор-акцепторной и относительные ориентации, а также молекулярного окружения 35,36. Снижение эффективности лобовая плита FRET может, возможно, возникают из-за изменений в геометрии решетки АХР. Однако, скорее всего, было бы связано с уменьшением доли АХР, которые упакованы в нано-масштаба постсинаптических молекулярной решетке 14.
Потеря связи между двигательными нейронами и мышечных волокон, кажется, непосредственной причиной мышечной слабости в двигательных нейронов заболевания и в сидячей старения 9,22,23. Общие методы и параметры для измерения НМС должны сделать его проще для различных исследовательских групп, чтобы сравнить и сопоставить опубликованные данные. Обмен подробных протоколов (и будущих улучшений на них) может помочь ускорить прогресс в понимании механизмов обеспечения НМС, и как это может повлиять на болезненных состояний.
Subscription Required. Please recommend JoVE to your librarian.
Materials
| Name | Company | Catalog Number | Comments |
| Scanning confocal microscope | Leica | DM 2000 with TCS SP2 system | Most scanning confocal microscopes should be suitable. |
| Zeiss | LSM 510 Meta | ||
| Leica | SPE-II | ||
| Alexa555-a-bungarotoxin (red-BGT) | Life technologies | B35451 | Used for labelling AChRs |
| Alexa647-α-bungarotoxin (far-red-BGT) | Life technologies | B35450 | Far red fluorescence: barely visible through the eyepiece |
| rabbit anti-synaptophysin | Life technologies | 18-0130 | Different batches of primary antibody differ in effective working dilution |
| FITC-anti-rapsyn mab1234 | Milipore | FCMAB134F | Monoclonal antibody conjugated to FITC |
| FITC-donkey anti-rabbit IgG | Jackson | 711-095-152 | Polyclonal secondary antibodies can vary in quality according to source and batch |
| Optimal Cutting Temperature compound (O.T.C.) | ProSciTech | IA018 | Cryostat embedding matrix for freezing muscles |
| DABCO | Sigma | 10981 | Mounting medium that slows photobleaching of fluorophores |
References
- Schmidt, N., et al. Neuregulin/ErbB regulate neuromuscular junction development by phosphorylation of α-dystrobrevin. J Cell Biol. 195, 1171-1184 (2011).
- Amenta, A. R., et al. Biglycan is an extracellular MuSK binding protein important for synapse stability. J Neurosci. 32, 2324-2334 (2012).
- Samuel, M. A., Valdez, G., Tapia, J. C., Lichtman, J. W., Sanes, J. R. Agrin and Synaptic Laminin Are Required to Maintain Adult Neuromuscular Junctions. PLOS ONE. 7, e46663 (2012).
- Valdez, G., et al. Attenuation of age-related changes in mouse neuromuscular synapses by caloric restriction and exercise. Proc Natl Acad Sci (USA). 107, 14863-14868 (2010).
- Yampolsky, P., Pacifici, P. G., Witzemann, V. Differential muscle-driven synaptic remodeling in the neuromuscular junction after denervation). Eur J Neurosci. 31, 646-658 (2010).
- Li, Y., Lee, Y., Thompson, W. J. Changes in Aging Mouse Neuromuscular Junctions Are Explained by Degeneration and Regeneration of Muscle Fiber Segments at the Synapse. J Neurosci. 31, 14910-14919 (2011).
- Zhu, H., Bhattacharyya, B. J., Lin, H., Gomez, C. M. Skeletal muscle IP3R1 receptors amplify physiological and pathological synaptic calcium signals. J Neurosci. 31, 15269-15283 (2011).
- Valdez, G., Tapia, J. C., Lichtman, J. W., Fox, M. A., Sanes, J. R. Shared resistance to aging and ALS in neuromuscular junctions of specific muscles. PLoS ONE. 7, e34640 (2012).
- Perez-Garcia, M. J., Burden, S. J. Increasing MuSK Activity Delays Denervation and Improves Motor Function in ALS Mice. Cell reports. 2, 1-6 (2012).
- Klooster, R., et al. Muscle-specific kinase myasthenia gravis IgG4 autoantibodies cause severe neuromuscular junction dysfunction in mice. Brain. 135, 1081-1101 (2012).
- Pratt, S. J., Shah, S. B., Ward, C. W., Inacio, M. P., Stains, J. P., Lovering, R. M. Effects of in vivo injury on the neuromuscular junction in healthy and dystrophic muscles. J Physiol. 591, 559-570 (2013).
- Landis, S. C., et al. A call for transparent reporting to optimize the predictive value of preclinical research. Nature. 490, 187-191 (2012).
- Gervásio, O. L., Phillips, W. D. Increased ratio of rapsyn to ACh receptor stabilizes postsynaptic receptors at the mouse neuromuscular synapse. J Physiol. 562, 673-685 (2005).
- Gervásio, O. L., Armson, P. F., Phillips, W. D. Developmental increase in the amount of rapsyn per acetylcholine receptor promotes postsynaptic receptor packing and stability. Dev Biol. 305, 262-275 (2007).
- Brockhausen, J., Cole, R. N., Gervásio, O. L., Ngo, S. T., Noakes, P. G., Phillips, W. D. Neural agrin increases postsynaptic ACh receptor packing by elevating rapsyn protein at the mouse neuromuscular synapse. Dev Neurobiol. 68, 1153-1169 (2008).
- Cole, R. N., Reddel, S. W., Gervásio, O. L., Phillips, W. D. Anti-MuSK patient antibodies disrupt the mouse neuromuscular junction. Ann Neurol. 63, 782-789 (2008).
- Morsch, M., Reddel, S. W., Ghazanfari, N., Toyka, K. V., Phillips, W. D. Muscle Specific Kinase autoantibodies cause synaptic failure through progressive wastage of postsynaptic acetylcholine receptors. Exp Neurol. 237, 237-286 (2012).
- Cole, R. N., Ghazanfari, N., Ngo, S. T., Gervasio, O. L., Reddel, S. W., Phillips, W. D. Patient autoantibodies deplete postsynaptic Muscle Specific Kinase leading to disassembly of the ACh receptor scaffold and myasthenia gravis in mice. J Physiol. 588, 3217-3229 (2010).
- Viegas, S., et al. Passive and active immunization models of MuSK-Ab positive myasthenia: Electrophysiological evidence for pre and postsynaptic defects. Exp Neurol. 234, 506-512 (2012).
- Morsch, M., Reddel, S. W., Ghazanfari, N., Toyka, K. V., Phillips, W. D. Pyridostigmine but not 3,4-diaminopyridine exacerbates ACh receptor loss and myasthenia induced in mice by Muscle Specific Kinase autoantibody. J Physiol. 591, 2747-2762 (2013).
- Ghazanfari, N., Morsch, M., Reddel, S. W., Liang, S. X., Phillips, W. D. Muscle Specific Kinase autoantibodies suppress the MuSK pathway and ACh receptor retention at the mouse neuromuscular junction. J Physiol. 592, 2881-2897 (2014).
- Cheng, A., Morsch, M., Murata, Y., Ghazanfari, N., Reddel, S. W., Phillips, W. D. Sequence of age-associated changes to the mouse neuromuscular junction and the protective effects of voluntary exercise. PLoS One. 8, e67970 (2013).
- Schaefer, A. M., Sanes, J. R., Lichtman, J. W. A compensatory subpopulation of motor neurons in a mouse model of amyotrophic lateral sclerosis. J Comp Neurol. 490, 209-219 (2005).
- Kilkenny, C., Browne, W. J., Cuthill, I. C., Emerson, M., Altman, D. G. Improving bioscience research reporting: the ARRIVE guidelines for reporting animal research. PLos Biol. 8, e1000412 (2010).
- Shimizu, S. Routes of Administration. The Laboratory Mouse. Hedrich, H. J., Bullock, G. , Elsevier. (2004).
- Chiasson, R. B. Laboratory anatomy of the white rat. , Brown. Dubuque, Iowa. (1988).
- Gage, G. J., Kipke, D. R., Shain, W. Whole Animal Perfusion Fixation for Rodents. J. Vis. Exp. (65), e3564 (2012).
- Mitra, A. K., Stroud McCarthy, M. P., M, R. Three-dimensional structure of the nicotinic acetylcholine receptor and location of the major associated 43-kD cytoskeletal protein, determined at 22A by low dose electron microscopy and x-ray diffraction to 12.5A. J Cell Biol. 109, 755-774 (1989).
- Paas, Y., et al. Electron microscopic evidence for nucleation and growth of 3D acetylcholine receptor microcrystals in structured lipid-detergent matrices. Proc. Natl Acad. Sci. (USA). 100, 11309-11314 (2003).
- Samson, A. O., Scherf, T., Eisenstein, M., Chill, J. H., Anglister, J. The mechanism for acetylhcoline receptor inhibition by α-neurotoxins and species-specific resistance to α-bungarotoxin revealed by NMR). Neuron. 35, 319-332 (2002).
- Ghazanfari, N., et al. Muscle Specific Kinase: Organiser of synaptic membrane domains. Int J Biochem Cell Biol. 43, 295-298 (2011).
- Ghazanfari, N., Morsch, M., Tse, N., Reddel, S. W., Phillips, W. D. Effects of the β2-adrenoceptor agonist, albuterol, in a mouse model of anti-MuSK myasthenia gravis. PLoS ONE. 9, e87840 (2014).
- Prakash, Y. S., Miller, S. M., Huang, M., Sieck, G. C. Morphology of diaphragm neuromuscular junctions on different fibre types. J Neurocytol. 25, 88-100 (1996).
- Salpeter, M. M., Harris, R. Distribution and turnover rate of acetylcholine receptors throughout the junction folds at a vertebrate neuromuscular junction. J Cell Biol. 96, 1781-1785 (1983).
- Soper, S. A., Nutter, H. L., Keller, R. A., Davis, L. M., Shera, E. B. The photophysical constants of several fluorescent dyes pertaining to ultrasensitive fluorescence spectroscopy. Photochem Photobiol. 57, 972-977 (1993).
- Panchuk-Voloshina, N., et al. Alexa dyes, a series of new fluorescent dyes that yield exceptionally bright, photostable conjugates. J Histochem Cytochem. 47, 1179-1188 (1999).
