

Abstract
La unión neuromuscular (UNM) es la sinapsis relé grande, colinérgica a través del cual las neuronas motoras de mamíferos controlan la contracción muscular voluntaria. Los cambios estructurales en la UNM pueden resultar en el fracaso de la neurotransmisión, resultando en debilidad, atrofia e incluso la muerte de la fibra muscular. Muchos estudios han investigado cómo las modificaciones genéticas o enfermedades pueden alterar la estructura de la UNM ratón. Desafortunadamente, puede ser difícil comparar directamente los resultados de estos estudios ya que a menudo emplean diferentes parámetros y métodos analíticos. Tres protocolos se describen aquí. El primero utiliza la proyección de intensidad máxima imágenes confocal para medir el área de receptor de acetilcolina (AChR) ricos en dominios de la membrana postsináptica en la placa terminal y el área de tinción de las vesículas sinápticas en la terminal nerviosa presináptica suprayacente. El segundo protocolo se comparan las intensidades relativas de inmunotinción para proteínas sinápticas en la membrana postsináptica. La tercera protocol utiliza la transferencia de energía de fluorescencia por resonancia (FRET) para detectar cambios en el embalaje de AChR postsinápticos en la placa terminal. Los protocolos se han desarrollado y perfeccionado a lo largo de una serie de estudios. Los factores que influyen en la calidad y la consistencia de los resultados se discuten y datos normativos se proporcionan para NMJs en ratones adultos jóvenes sanos.
Introduction
La unión neuromuscular (NMJ) es la sinapsis relé crítico que media la comunicación entre el sistema nervioso y músculo esquelético. Se requiere para todo el movimiento voluntario. La microscopía de fluorescencia se ha utilizado para estudiar los efectos de los transgenes en el ratón NMJ 1-3 o para comparar los efectos de la edad, la dieta, el ejercicio y la enfermedad en roedores NMJs 4-11. Tales estudios han enseñado mucho acerca de la fisiología y la fisiopatología de la UNM, pero los diversos parámetros informado (por ejemplo, área AChR, área de placa terminal, longitud del perímetro, los índices de fragmentación) a menudo hacen que sea difícil comparar los resultados de estos estudios. Hay una expectativa creciente de investigadores pre-clínicos para ser capaces de demostrar la reproducibilidad, en particular en estudios con modelos de roedores de enfermedad 12. Los protocolos descritos aquí fueron refinados a través de una serie de estudios que investigaron ch desarrollo, fisiológico y fisiopatológicoanges de la UNM. Tales estudios requieren la medición de la zona de especializaciones sinápticas en la placa motora terminal del ratón y la densidad relativa de embalaje de las proteínas sinápticas dentro de especializaciones postsinápticos 13-15.
La utilidad de estos métodos se ilustra por estudios recientes en un modelo de ratón de anti-MuSK miastenia gravis. Las inyecciones diarias de IgG de miastenia anti-MuSK positivos gravis pacientes en ratones adultos causó que se vuelven débiles dentro de 2 semanas 16. Imágenes de máxima proyección confocal de secciones musculares que fueron etiquetados doble para sinaptofisina (en terminales nerviosas) y AChR postsinápticos revelaron una disminución progresiva en el área de tinción AChR como el principal cambio. Es importante destacar que la tasa de disminución fue suficiente para explicar la disminución comparables en la amplitud de los potenciales sinápticos, fracaso de la transmisión sináptica y debilidad muscular 17,18. Cualitativamente hallazgos similares fueron reportados por otros grupos de investigación10,19. Los mismos métodos de medición NMJ ya se han utilizado para evaluar el impacto de tres fármacos para el tratamiento de la miastenia gravis anti-MuSK en este modelo de ratón 20,21.
Envejecimiento sedentario puede llevar a la pérdida de las conexiones neuromusculares. Los protocolos descritos aquí han revelado una disminución asociada a la edad en la zona de sinaptofisina terminación nerviosa en placas motoras como ratones progresan en la vejez. Los mismos métodos revelaron que el ejercicio voluntario podría evitar en gran medida la reducción de la zona de la terminal nerviosa presináptica 22, en consonancia con el trabajo previo de otros grupos 4. La pérdida de las conexiones neuromusculares también se produce en el modelo de ratón SOD1G93A de la esclerosis lateral amiotrófica 9,23.
Los estudios mencionados anteriormente demuestran que una variedad de condiciones de salud puede conducir a la reducción de la superficie de cualquiera de las especialidades pre o post-sináptica en la UNM. Esto puede producir problemas para la diversión sinápticacción o puede anunciar la pérdida completa de la conexión neuromuscular. Tres protocolos se describen que permiten la cuantificación de la zona y la densidad de especializaciones sinápticas. El propósito de la primera protocolo es proporcionar una medida práctica y reproducible de las áreas de pre y post-sinápticas especializaciones y su alineación en NMJs de mamíferos, usando microscopía de fluorescencia. Confocal de imágenes de proyección máximo de dos dimensiones y análisis de imagen con NIH ImageJ se utiliza para detectar cambios en el área de tinción sinaptofisina (vesículas sinápticas), AChR postsinápticos y área de superposición sináptica. Parámetros confocal de imágenes (ganancia y nivel de compensación) se han optimizado para cada NMJ a fin de maximizar la información visual utilizado para discernir el área de especialización sináptica. Insuficiencia neuromuscular también puede resultar de los cambios en la densidad de AChR postsináptica y / o otras proteínas sinápticas. El segundo protocolo se puede aplicar para detectar cambios en la densidad relativa de las proteínas postsinápticos talescomo el almizcle, RapSyn, distroglicano, fosforilada quinasa Src y fosforilada AChR 18,21.
En la miastenia gravis, una densidad reducida de AChR dentro de la membrana postsináptica es la causa inmediata del fracaso sináptica y la debilidad muscular. El tercer protocolo describe un método de fluorescencia la transferencia de energía de resonancia (FRET) para evaluar los cambios en la proximidad de AChR adyacentes dentro de las membranas postsinápticas 14,15. Este método detecta la transferencia de energía entre AChR vecinos marcados con fluorescente-α-bungarotoxina (BGT). FRET se produce sólo cuando el donante y aceptor de sondas fluorescentes son de menos de 10 nm de distancia. Esto puede revelar cambios (submicroscópicas) en la estanqueidad de embalaje AChR que pueden relacionarse directamente a la amplitud de los potenciales sinápticos.
Estos tres protocolos, refinados durante la última década, proporcionan medidas complementarias de integridad NMJ de una manera consistente y reproducible. El uso de protocolos estandarizados unaparámetros nd deben facilitar la comparación de los efectos de los genes y las intervenciones ambientales sobre los mamíferos NMJ.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
NOTA: El diseño, la realización y presentación de informes de los experimentos con animales deben tener en cuenta las directrices actuales 24. Este tipo de trabajo debe ser aprobado previamente por la autoridad local de protección de los animales (en nuestro caso, el Comité de Ética Animal de la Universidad de Sydney).
1. La eutanasia del animal y la disección muscular
- Transferir el ratón desde la sala de espera a una habitación separada donde se practicó la eutanasia con una inyección intraperitoneal de solución de pentobarbital (30 mg / kg), utilizando el método de manejo del ratón detallado por Shimizu 25. Coloca el ratón de nuevo en su jaula.
- Una vez que la respiración del ratón se ha detenido durante más de 1 minuto, probar el reflejo pies retirada pellizcando suavemente el pie, y el reflejo corneal por la ligera el cepillado de la córnea. Sólo cuando las respuestas reflejas están ausentes se puede preparar el ratón para la disección.
- Consulte con un atlas de anatomía roedor tal como Chiasson 26 y / o buscar la ayuda de una experienanatomista ced antes de intentar la disección del músculo de interés. En cada caso eliminar el vello de la piel que recubre el uso de una pequeña máquina de afeitar eléctrica antes de abrir la piel para exponer el músculo.
NOTA: La disección será diferente para cada músculo anatómicamente distinta. - Con unas pinzas romas liberar el músculo desde que recubre las membranas y los tejidos circundantes. Agarrar y cortar el tendón distal para separar el músculo de su inserción.
- Embromar suavemente y recortar el músculo libre del tejido circundante de vuelta a su origen. Brevemente colocar el músculo recién diseccionado en 0,1 M tampón fosfato salino (PBS) solución o solución de Ringer antes del procesamiento adicional.
2. Preparación del músculo por cryosectioning
NOTA: la conservación estructural óptima se puede lograr por perfusión animal completo como se detalla anteriormente 27, o la fijación de inmersión (para los músculos pequeños) como se describe en el paso opcional de 2,1. Sin embargo,4% de paraformaldehído fijación puede perjudicar la posterior tinción con muchas sondas de anticuerpos y con fluorescente-BGT. El glutaraldehído en particular debe ser evitado. Si los músculos no han de ser fijados deben ser inmediatamente se congelaron rápidamente (proceder a 2,3).
- Fijación de inmersión Opcional: Pin del músculo a la cera en una placa de Petri en longitud de reposo. Cubrir el músculo con 2% w / v de paraformaldehído (recién disuelto en PBS) durante 2 horas a RT. Lavar con 3 cambios de PBS más de 30 min (3 x 10 min) y luego reemplace el PBS con 30% w / v de sacarosa en PBS e incubar O / N a 4 ° C.
- Hacer moldes ('') barcos de antemano por plegado de 2 cm x 1,5 cm trozos de papel de aluminio como se muestra en la Figura 1. Coloque un trozo de membrana de nitrocelulosa en la parte inferior de la embarcación. Vierta suavemente criostato incrustar matriz (tabla Materiales) en el barco hasta una profundidad de 2 mm, teniendo cuidado de evitar las burbujas de aire. Coloque el músculo en la barca, alineándola con las bolígrafo líneas enla nitrocelulosa. Añadir matriz de la incrustación de más de manera que cubra completamente el músculo (Figura 1).
- Tubos de polipropileno Pre-etiqueta con un marcador indeleble. Coloque una gota de agua en cada tubo y enfriar el tubo en nitrógeno líquido.
NOTA: La gota de agua congelada mantiene la presión de vapor y evita la desecación durante prolongada -80 ° C de almacenamiento - El uso de un protector de cara, gruesos guantes de protección y un gran par de fórceps romos, reducir parcialmente un vaso de precipitados de metal pequeño (3 cm de diámetro, 8 cm de profundidad) que contiene 2 cm profundidad de isopentano en un recipiente de nitrógeno líquido durante 30 seg. Retirar el vaso y lo coloca en la parte superior del banco. El uso de un par más pequeño de fórceps romos colocar el molde que contiene el músculo y la incrustación de la matriz en el isopentano enfriado. Tenga cuidado para evitar la mezcla de nitrógeno líquido con el isopentano.
- Permitir 2 min para el bloque de congelar completamente antes de usar fórceps romos para levantar el bloque congelado y sellarlo en el pr correctae-etiquetados y tubo de pre-enfriado (paso 2.3).
- Guarde los tubos temporalmente en el nitrógeno líquido antes de la transferencia a -80 ° C. Registrar todas las muestras en una hoja de cálculo de los contenidos del congelador.
3. cryosectioning y fluorescencia de tinción para En Face Imágenes de NMJs
- Despegar el molde de aluminio. Dentro de la -20 ° C cámara del criostato fijar el bloque congelado al mandril criostato de manera que corte 20 micras cryosections paralelos al eje largo de las fibras musculares (Figura 1). Recoge las secciones sobre poli-L-lisina o recubiertos con gelatina portaobjetos de microscopio.
- NOTA: Omita este paso si el tejido se fija antes de la congelación. Después de dejar 30 min para las secciones se sequen sobre los portaobjetos, fijarlos mediante la colocación de una gota de 2% de paraformaldehído en PBS sobre cada sección durante 15 minutos a RT.
- Lavar los portaobjetos 3 x 10 min en PBS en una jarra Coplin, y luego sumergir los portaobjetos en PBS que contiene glicina 0,1 M durante 30 min para bloquear los grupos aldehído residuales.
- Lavado de los portaobjetos durante 10 min en PBS, a continuación, sumergir en metanol (enfriada a -20 ºC) durante 7 min. Este paso permeabilización es una parte rutinaria de doble etiquetado con fluorescente-BGT y anti-sinaptofisina pero puede afectar negativamente a la inmunotinción para algunas otras proteínas.
- Lavar los portaobjetos 2 x 10 min en PBS luego colocar cada diapositiva en una cámara húmeda estable y nivelada. Inmediatamente cubrir cada sección con 20 l de solución de bloqueo (0,2% Triton X-100, 2% de albúmina de suero bovino (BSA) en PBS) durante 1 hora a RT. Secciones no se debe permitir que se seque en cualquier etapa del proceso de inmunotinción.
- Llevar a cabo la incubación primaria: Tomar una diapositiva a la vez retire con cuidado el exceso de solución de bloqueo de más de cada sección y reemplazarlo con 20 l de conejo anti-sinaptofisina (diluido 1: 200 en la solución de bloqueo).
- Incluir un portaobjetos de control negativo que se incubó con solución de bloqueo solamente. Este "control de anticuerpos no-primaria"Es esencial en cada carrera inmunotinción.
- Cuidar que el anticuerpo primario se mantiene en su lugar sobre cada sección, cierre la cámara húmeda e incubar durante 1-2 días a 4 ºC.
- Inspeccionar cada sección para confirmar que el anticuerpo primario permanece en su lugar. Utilice una pipeta Pasteur para enjuagar suavemente cada portaobjetos con PBS y colocarlo en un frasco Coplin. Lave todas las diapositivas 3 x 10 min en PBS.
- Llevar a cabo la incubación secundaria. Tomando una diapositiva a la vez, retire con cuidado el exceso de PBS, lo pondré en la cámara húmeda y cubrir cada sección con 20 l de una mezcla que contiene FITC-conjugado IgG de burro anti-conejo y BGT conjugado con tetrametil rodamina u otro fluoróforo rojo (TRITC- / redBGT; 5 g / ml) diluido en solución de bloqueo. Incubar a temperatura ambiente durante 2 hr.
- Lavar los portaobjetos 3 x 10 min en PBS en frascos Coplin.
- Tomando una diapositiva a la vez, retire con cuidado el exceso de PBS y montar con un cubreobjetos utilizando un volumen mínimo de, glicerobasado en l, se desvanecen-resistencia medio de montaje. Selle los bordes de los cubreobjetos con barniz de uñas transparente. Deje que se seque duro.
- Guarde las diapositivas de la oscuridad a 4 ° C durante un máximo de una semana, oa -20 ° C durante largos períodos de almacenamiento (hasta varios meses).
4. imparcial Muestreo y En Face Imaging de Motor Placas de extremo
- Blind las diapositivas mediante el etiquetado de cada diapositiva con un número de código aleatorio que permanece conocida sólo por un segundo investigador (que no participe en el análisis). Como resultado, el operador permanece ciego a los grupos de tratamiento hasta que la cuantificación de parámetros NMJ es completa para todas las muestras.
- Posicionar el cursor en la platina del microscopio y ver debajo amplia iluminación de campo con el conjunto filtro TRITC (63X aceite 1,3 objetivo NA). Mueva progresivamente (campo por campo) de izquierda a derecha y de regreso hasta que una placa terminal aparece en el campo (Figura 2).
Criterio de muestreo:: NOTA Cada estructura manchado-AChR que es relativamenteplana y se enfrenta el objetivo (es decir, se extiende <15 m en la dimensión z) se considera una placa terminal y se forma la imagen para el análisis (medias lunas de tinción AChR representan secciones transversales a través de las placas terminales y por lo tanto están excluidos). - Con el conjunto pinhole confocal a 1,0 unidad de Airy y láser de baja potencia optimizar la ganancia y el offset niveles para TRITC / rojo-BGT (láser 532 nm) en la placa terminal que va a ser fotografiado. Siguiente optimizar FITC / fluorescencia sinaptofisina usando el láser de 488 nm. Se recoge una z-pila de la placa terminal con un intervalo de 0,7 m entre cada rebanada óptica. Guarde las imágenes con un nombre de archivo que incluye la fecha de la sesión de imágenes, el nombre en clave de la diapositiva y el número de la placa terminal.
NOTA: Las exploraciones utilizando el 488 nm y 532 nm láseres (FITC y TRITC) deben recoger secuencialmente (no al mismo tiempo) para evitar la contaminación del canal FITC por fluorescencia del fluoróforo rojo y viceversa (sangrar a través). - Repetir el muestreo de unnd obtención de imágenes de los pasos 4.2 a 4.3 hasta 20 placas terminales se recogen de la corredera / muestra.
- Cambie a la diapositiva siguiente código y repetir 04.02 a 04.04. Repita esto para cada uno de los portaobjetos codificados.
- Recoge unas cuantas imágenes de placas terminales de la corredera de control (no-primaria control del anticuerpo) mediante la configuración confocal que se encontraron óptimo para las diapositivas experimentales (el canal de fluorescencia FITC debe aparecer oscuro).
- Al final de la sesión de transferencia de los archivos de imagen confocal a otro ordenador y una copia de seguridad de los archivos originales en una unidad o un servidor externo.
5. Medir el Área de Synaptic Especializaciones en Imágenes en face
- Utilice NIH ImageJ software gratuito (http://imagej.nih.gov/ij/) para preparar las imágenes de proyección máxima (MIP) de cada z-stack. Guardarlas como archivos TIFF (Figura 2A y B). Los nombres de archivo deben incluir la fecha de la sesión de imágenes, código de ejemplo, el número de placa de extremo y el canal fluorescente (por ejemplo, 060414_5723_7_FITC.tiff).
- Abra la imagen z-proyección en ImageJ. Seleccione la imagen del canal receptor de acetilcolina (Figura 3A) y seleccione: Imagen> Tipo> 8 bits para convertir la imagen de color RGB de 24 bits en tres imágenes en escala de grises de 8 bits en la pantalla.
- Con la herramienta polígono ImageJ dibujar un esbozo en torno a la placa terminal de interés en el redBGT manchado (CADH) canal a fin de incluir todas las regiones manchadas aparentes de la placa terminal individuo en particular, mientras que excluyendo cualquier tinción que no se originan en la placa terminal de interés ( Figura 3C).
- Aplique un umbral mínimo de intensidad de la imagen seleccionando: Imagen> Ajustar> Umbral (Figura 3E y capturas de pantalla ImageJ asociados).
- Ajuste el nivel de umbral para aislar las partes teñidas con AChR excluyendo rodea señal de fondo como sub-umbral (Figura 3E). Abra un segundo aireow con la imagen del original (tono continuo) inmediatamente junto a la ventana para la comparación, para facilitar la decisión sobre el valor de umbral. Registrar el valor umbral para análisis posterior uso en colocalización.
- Conservando el contorno del polígono alrededor de la placa terminal seleccione: Analizar> Analizar Partículas. En el menú emergente especificar el rango de tamaños como: 50 a infinito píxeles (esto elimina artefactos diminutos que surgen de ruido eléctrico en el fotomultiplicador).
- Analizar comando partículas genera una ventana con una lista de áreas supra-umbral discretos y sus valores de intensidad de fluorescencia numeradas que aparecen en la imagen binaria (Figura 3G y asociado ImageJ captura de pantalla). Copie estos datos en una hoja de cálculo de etiquetado.
- Medir la superficie total placa terminal (área dentro del polígono) seleccionando: Analizar> Medida. Esto produce el área total de la placa terminal. Copiar y pegar los datos para áreas AChR e intensidades enuna hoja de cálculo asegurándose de etiquetar apropiadamente columnas, las filas se utilizará para placas terminales individuales para diapositivas específicas.
- Cambia al canal de fluorescencia anti-sinaptofisina y repita los pasos 5.1 hasta 5.5, pero para el canal FITC (Figura 3B, D y F). El objetivo es ajustar el umbral de manera que crea una imagen binaria que, tan estrechamente como sea posible, coincide con los límites de tinción según lo percibido por el ojo. Registre el valor umbral.
- Mida el área de superposición mediante la aplicación de los siguientes pasos: Abra el archivo original que contiene las dos imágenes del canal, que se dividió en dos imágenes separadas seleccionando: Imagen> Pilas> Pila a las imágenes.
- Utilizando el plugin Colocalización (descargado e instalado desde la página web ImageJ) Seleccionar: Pluggin> Colocalización y de entrada los valores umbral registrados previamente para los canales CADH y nerviosas en el respectivo canal de consulta bbuey. Esto dará lugar a una imagen de superposición en píxeles blancos (Figura 3H y capturas de pantalla ImageJ asociados).
- Convertir la imagen de superposición recién creado en un formato de escala de grises y aplicar un umbral para el valor máximo. El umbral máximo sólo seleccionará los píxeles blancos, correspondiente a la zona de superposición de los dos canales anteriores. Registro en la hoja de cálculo el valor del área resultante de 'colocalización ", que representa el área de superposición de píxeles.
- Prepare una hoja de cálculo de medias de la muestra de datos, calcular y representar gráficamente las desviaciones estándar y los errores estándar como histogramas o diagramas de dispersión 20,22. Tenga en cuenta que el valor de n generalmente representa el número de ratones por grupo de muestra para fines estadísticos.
- Placa terminal Parcela áreas AChR como diagramas de dispersión o histogramas de frecuencia para determinar si los datos se distribuyen normalmente antes de la prueba estadística (Figura 6).
6. La tinción relativaIntensidades compararon mediante secciones transversales ópticas
NOTA: Para este proceso todas las muestras de músculo junto protocolo e imagen en una sola sesión confocal. En la planificación de un experimento permitirá hasta 30 minutos de tiempo de formación de imágenes por muestra muscular.
- Cortar 15 micras cryosections transversal al eje largo de las fibras musculares y recoger en portaobjetos como se ha descrito en el paso 3.1.
- Llevar a cabo la tinción de fluorescencia como se describe en los pasos 3.2 a 3.13.
- Código los portaobjetos teñidos de modo que las imágenes y los análisis se llevan a cabo con los ciegos operador al grupo de tratamiento, tal como se describe en el paso 4.1.
- Utilizando un objetivo de 40X de fluorescencia (NA 0.75) examinar brevemente una sección de cada diapositiva para determinar un único aumento y compensación ajuste del nivel de AChR que será adecuado para todas las placas terminales en todas las diapositivas de muestra. La placa terminal más brillante debe entonces estar justo por debajo de 256 en la escala de grises. Esta optimización se debe hacer por separado para el segundo fluorescenccanal e (recogido sucesivamente). Anote la ganancia fija y compensar ajustes de nivel y no alteran ellos durante toda la sesión de imágenes.
- Recoger imágenes de una diapositiva estándar de fluorescencia (por ejemplo, perlas fluorescentes no blanqueantes), utilizando los mismos parámetros, al principio y al final de la sesión confocal para detectar cualquier posible fluctuación en la intensidad del láser.
- Utilice el canal AChR para escanear la diapositiva progresivamente para localizar placas terminales.
- Enfoque de encontrar el plano de sección óptica individual en cada campo microscópico que contiene el mayor número de placas terminales teñidos con AChR.
- Analiza esta sección óptica solo dos veces y guardar la imagen en promedio (Figura 4G).
- Manteniendo el mismo interruptor de plano focal para el segundo canal de fluorescencia (proteína de interés) y recoger la imagen como en el paso 6.8. Guarde el archivo de imagen, incluso en el nombre de archivo: fecha de la sesión de imágenes, código de ejemplo, el número de la imagen y un símbolo para indicar el canal fluorescente.
- Mueva la etapa a la siguiente campo que contiene una o más placas de extremo y repita el paso 6.8 hasta 6.9. Repita este procedimiento hasta un total de 60 placas terminales se va a ver.
- Al final de la sesión de imágenes de transferencia de todos los archivos a otro ordenador y los respaldan.
- Abra cada archivo de imagen original y mientras ve el canal CADH, seleccionar: Imagen> Pilas> Pila de Imágenes, para dividir los canales.
- Seleccionar: Imagen> Tipo> 8 bits para convertir al formato de escala de grises de 8 bits en la pantalla. Haga esto por dos canales de fluorescencia.
- Seleccionar: Imagen> Pilas> Imágenes de pila. Abra una nueva pila de dos imágenes de 8 bits previamente separados. Entonces se puede cambiar convenientemente entre los dos canales de fluorescencia dentro de la ventanilla única.
- Utilice la herramienta de polígono para dibujar un line firmemente alrededor de la frontera de la tinción AChR (Figura 4I).
- Seleccionar: Analizar> Medir para medir la intensidad media de píxeles para AChR en la zona delimitada (tenga en cuenta la importancia de la elaboración de la línea de fuerza). Copia este valor en una hoja de cálculo de etiquetado.
- Conservando el mismo esquema polígono (para definir el área a medir), cambiar a la segunda cadena fluorescente (por ejemplo, la Figura 4B, D, F) y seleccione: Analizar> medida. Esto dará lugar a la intensidad media de la tinción de la proteína de interés en la zona sináptica definido por tinción AChR.
- Elija un área lejos de tinción placa terminal visible a continuación, seleccione: Analizar> Medir para medir la intensidad media de fluorescencia de fondo. Repita esto para el otro canal de fluorescencia / s y copiar los valores de fondo en la hoja de cálculo de los valores de fluorescencia.
- SubTracto los valores promedio de los valores de fondo de placa terminal para obtener las intensidades corregidas para CADH y la proteína de interés en cada placa terminal.
- Divide los valores de intensidad de placa terminal corregidos de la proteína de interés por la intensidad de la fluorescencia BGT corregido para dar los ratios de intensidad de fluorescencia 14,21
7. La comparación de la densidad postsináptica Membrana AChR Usando FRET
NOTA: Este protocolo evalúa el grado en que AChR están estrechamente empaquetados (<10 nm espaciamiento) en la membrana postsináptica. La combinación de los donantes y fluoróforo aceptor precisa es crítico para este ensayo FRET. Los nombres y detalles de los fluoróforos se dan en la tabla de Materiales. Sus propiedades espectrales, en relación con FRET, se discuten en nuestros trabajos anteriores 14,15.
- Preparar cryosections transversales fijas como se describe en la sección 6.1. Todos los grupos de la muestra deben ser procesados juntos e imagend en el mismo período de sesiones confocal.
- Mezclar bien 2,5 g / ml-BGT rojo (FRET donante) con 10 g / ml rojo lejano-BGT (FRET aceptor) con solución de bloqueo en un pequeño tubo de plástico pipeteando arriba y abajo 12 veces. Esta mezcla 1: 4 molar maximiza la eficiencia de FRET 14.
- Coloque cada portaobjetos en una cámara humidificada, cubrir cuidadosamente cada sección con una gota (12 l) de la mezcla anterior y se incuba durante 1,5 horas a RT.
- Control de las secciones: cubren un pequeño número de secciones con 2,5 g / ml rojo-BGT (sólo donante; controles C1 etiquetados), y también algunas secciones con 10 g / ml rojo lejano-BGT (sólo aceptor; controles C2 marcados). Incubar estos controles como en el paso 7.3.
- Lavar los portaobjetos 3 x 10 min en PBS y montaje en basado en glicerol, fade-resistiendo medio de montaje (consulte el paso 3.12).
- Tomar muestras de placas terminales como en el paso 6.7. La fluorescencia del donante y aceptor debe ser perfectamente co-localiza en placas terminales debido a la aleatoria de unión de las moléculas fluorescentes-BGT.
- Imágenes de control: Usando el objetivo de 40X y láser de baja potencia optimizar la ganancia redBGT y compensar los ajustes de nivel para placas terminales de un C1 corredera de control. Optimizar la ganancia extrema redBGT y los niveles de desplazamiento de las placas terminales de corredera de control C2. Confirme la ausencia de fluorescencia de purga a través.
- Sin cambiar la potencia del láser, ganan o configuración de nivel de desplazamiento se mueven a las diapositivas experimentales y recogen imágenes (pre-fotoblanquear) para ambos canales de fluorescencia.
- Selectivamente photobleach el rojo lejano-BGT sobre una porción de una sola placa de extremo por el zoom en el área de escaneo y luego escanear 10 veces con el láser de 633 nm a 100% de potencia. La fluorescencia en la zona escaneada debe convertirse tenue.
- Restablecer la potencia del láser y zoom y recoger imágenes post-blanqueador en ambos canales fluorescentes utilizando la configuración confocal establecidos en 7.7.
- Calcular la eficiencia de FRET (E) del porcentaje de aumento de los donantes (rojo-BGT) fluorescencia siguiente fotoblanqueador del aceptor (rojo lejano-BGT) de acuerdo con la siguiente fórmula *:

* Para todas las situaciones en las que la fluorescencia del donante aumenta después de photobleaching aceptor.

Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Medición de Synaptic Área en la UNM
Cualquier estimación del área se basa en el dibujo de un límite para definir la extensión de especializaciones sinápticas. En los músculos adultos jóvenes sanos imágenes NMJ deben mostrar los límites bien definidos, tanto para AChR y tinción sinaptofisina (Figura 2 A y B). La intensidad de fluorescencia para ambos AChR y sinaptofisina aumenta de forma pronunciada en el límite entre la porción peri-sináptica y sináptica de la placa motora terminal (Figura 5A 'y B'). Para este tipo de imágenes de un umbral mínimo (justo por encima del fondo extrasynaptic fluorescencia) se aislar fácilmente el área de AChR-rico o sinaptofisina-rica de la placa terminal (líneas de trazos en la horizontal (Figura 5A 'y B')). En ratones de edad avanzada y en algunos estados de la enfermedad tinción placa terminal para AChR puede ser menos intenso, bordes de racimo AChR pueden aparecer borrosa y lare pueden ser mayores niveles de autofluorescencia extrasynaptic (Figura 5D; 17,22). Tinción de fluorescencia con límites indistintos puede introducir errores en las estimaciones de área sináptica. En todas las situaciones, el objetivo es elegir un umbral que produce una imagen binaria similar en forma y tamaño a la AChR- o áreas sinaptofisina-rica tal como aparecen a la vista en la imagen original, de tono continuo. La realización de análisis de los ciegos al grupo de tratamiento debería reducir el riesgo de sesgo subjetivo en el paso de umbral (paso 4.1). A veces las imágenes de placa terminal débil o borrosa puede ser resultado de procesamiento de sub-óptima. Figura 2C y D muestra un ejemplo de una imagen de baja calidad de una placa terminal 2 meses de edad ratón sano. Bordes borrosos y manchas sinaptofisina débil pueden haber surgido en este caso de la descongelación parcial y recongelación del músculo antes de cryosectioning. Algunas secciones de (control positivo) muscular joven y sana se debe seccionar unnd procesado en paralelo con las muestras experimentales para asegurar que cualquier deterioro evidente en las imágenes NMJ no se debe a problemas con la inmunotinción. Los lotes de imágenes comprometidas por el procesamiento sub-óptima deben ser excluidos de los análisis.
Para en face imágenes z-stack, 15-20 placas terminales es un tamaño de muestra razonable para estimar las zonas sinápticas. Una amplia diversidad de formas y tamaños de NMJs se encuentran dentro de cualquier músculo determinado. Diagramas de dispersión revelan un rango considerable en el área de AChR-rica entre placas terminales del músculo tibial anterior de cualquier ratón individual (Figura 6A). Sin embargo, el área de AChR promedio (en base a 15-20 en images cara placa terminal) fue similar entre los siete ratones de muestra (~ 200 m 2; Figura 6A). El área de tinción sinaptofisina placa terminal también varió considerablemente entre placas terminales de un músculo determinado. Una vez más, el uso de un tamaño de muestra de 15 a 20 placas terminales de la zona media de la sinaptofisina placa terminal fue similar entre los ratones 7 estudió (~ 170 m 2; Figura 6B). Los histogramas de frecuencia de los datos agrupados revelaron distribuciones aproximadamente normales para el área de AChR placa terminal y sinaptofisina (Figura 6 A 'y B'). Sin embargo una distribución normal de las zonas sinápticas no se puede asumir en estados de enfermedad tales como la miastenia gravis 16,20. Esto puede afectar a la elección de la prueba estadística.
Tabla I enumera las áreas de especialización de pre y post-sinápticas para NMJs para sana 2 meses de edad (adultos jóvenes) hembra C57BL / 6J de estudios previos. Las áreas de especialización tanto en pre y post-sinápticas disminuyeron con el envejecimiento sedentario 22. Área de AChR también se redujo notablemente en ratones inyectados con IgG de miastenia gravis anti-MuSK pacientes 17,21. Ratones miasténicas tratados con el fármaco inhibidor de la colinesterasa, piridostigmina, muestran una reducción adicional de la placa terminal área AChR 20 </ Sup>.
La intensidad relativa de placa terminal Fluorescencia de etiquetado
La intensidad relativa de etiquetado inmunofluorescencia puede revelar cambios en la densidad de una proteína de interés-de-sináptica con la edad, el genotipo y / o estado de enfermedad. AChR fluorescencia (rojo-BGT o rojo lejano-BGT) se utilizó por primera vez para definir la ubicación de la UNM. El brillo de la fluorescencia dentro de la zona AChR ricos en 8 bits se utiliza para evaluar los cambios en la concentración de la proteína de interés, en relación con los animales control. En las secciones transversales de los AChR placa terminal aparecen típicamente como una forma de media luna, pero esta forma es a menudo irregular (Figura 4A, C, E, H). Baja intensidad de fluorescencia de fondo por lo general revela si un parche de tinción AChR representa una única placa terminal, o dos placas terminales separados situados en las fibras musculares adyacentes. Muchas proteínas sinápticas (como RapSyn, almizcle y Src) soncolocalized con AChR en la placa terminal (Figura 4A - D). La tinción de inmunofluorescencia con anticuerpos fosfo-específicos también se puede utilizar para comparar el efecto de las intervenciones experimentales en el estado de fosforilación de determinadas proteínas de la membrana postsináptica 21.
La fiabilidad y la reproducibilidad de la medición de la intensidad de fluorescencia depende en gran medida de la integridad del músculo congelado y la calidad de la inmunotinción. Los músculos deben ser disecados y se congelaron inmediatamente o paraformaldehído-fijo (a pocos minutos de la muerte del animal) para evitar los cambios degenerativos de la UNM. La inmunotinción depende altamente de la calidad de los reactivos y optimizar el protocolo de tinción para anticuerpos específicos. Para cualquier nuevo lote de anticuerpos primaria inmunoticción experimentos piloto son necesarios. Cryosections recién cortadas de los músculos jóvenes sanos se incuban con diluciones en serie de 2 veces del anticuerpo primario. Laconocido anticuerpo secundario fiable se utiliza y los resultados se comparan. Si se conoce la proteína de interés para ser restringida a la NMJ entonces la mejor concentración de anticuerpo es la que produce la mayor proporción de intensidad de fluorescencia NMJ relativa a la encontrada en partes extrasinápticos del músculo (fluorescencia de fondo). Extrasynaptic intensidad de fluorescencia (no específica presumiblemente) normalmente no debe superar el 15% de la intensidad de fluorescencia placa terminal. Del mismo modo 'sin control de anticuerpo primario (' secciones incubadas sólo con el anticuerpo secundario) debe aparecer oscuro, confirmando que el anticuerpo secundario no se unen de forma no específica. La calidad de los diferentes lotes de (policlonales) anticuerpos secundarios pueden variar lotes de anticuerpos secundarios marcadamente alternativas debe compararse antes de establecer un protocolo estándar. La prueba ideal para la especificidad de la inmunofluorescencia implica una comparación directa de las secciones de los ratones de tipo salvaje y las secciones de control negativo de los ratones tsombrero carecen de la (génica) proteína de intereses. Las siguientes referencias describen la cuantificación de la tinción de fluorescencia placa terminal de almizcle, RapSyn, distroglicano, src y AChR 13,14,18,21.
Los tamaños de muestra pequeños introducir errores en las estimaciones de la intensidad de fluorescencia relativa. Placas terminales individuales variaron considerablemente en la intensidad de fluorescencia. Es de suponer que esta variabilidad entre placas terminales dentro de un músculo dado refleja la diversidad de la estructura y de azar NMJ diferencias en la sección óptica de la muestra. Sin embargo, aumentar el número de placas terminales incluidos en la muestra resulta en una estimación más estable de la intensidad de fluorescencia media (Figura 7). Para estimar la intensidad media de fluorescencia placa terminal 40-60 placas terminales de cada muestra de músculo deben ser promediados.
AChR-AChR FRET
Cada AChR es un pentámero con dos sitios de unión para BGT (situados uno en cada subunidad alfa). La unión derojo-BGT y rojo lejano-BGT a estos dos sitios le permitan establecer una separación donante-aceptor de alrededor 9nm 28-30. Por lo tanto la baja eficiencia de FRET se puede detectar incluso antes AChR se reúnen en grupos de 14. Sin embargo, la eficiencia de FRET en placas terminales murinos más o menos se duplicó después del nacimiento, en consonancia con la incorporación más eficiente de AChR en una celosía membrana postsináptica fuertemente empaquetada 14. Usando rojo-BGT y rojo lejano-BGT como el donante de FRET y aceptor (respectivamente), las placas terminales de 1-2 meses de edad ratones producen eficiencias promedio de FRET que van desde 20 hasta 37% (Tabla 2). Eficiencia de FRET de 20% o más se cree que representan apretado embalaje de AChR 14. La eficiencia de FRET placa terminal se redujo ligeramente tras la denervación 14, y reduce notablemente después de ratones fueron inyectados con IgG de miastenia anti-MuSK-positivo gravis pacientes 18. Estas son condiciones en las que AChR individuales están menos estrechamente empaquetados en el postsyandamio membrana Naptic por el sistema MuSK / RapSyn postsináptica 31.
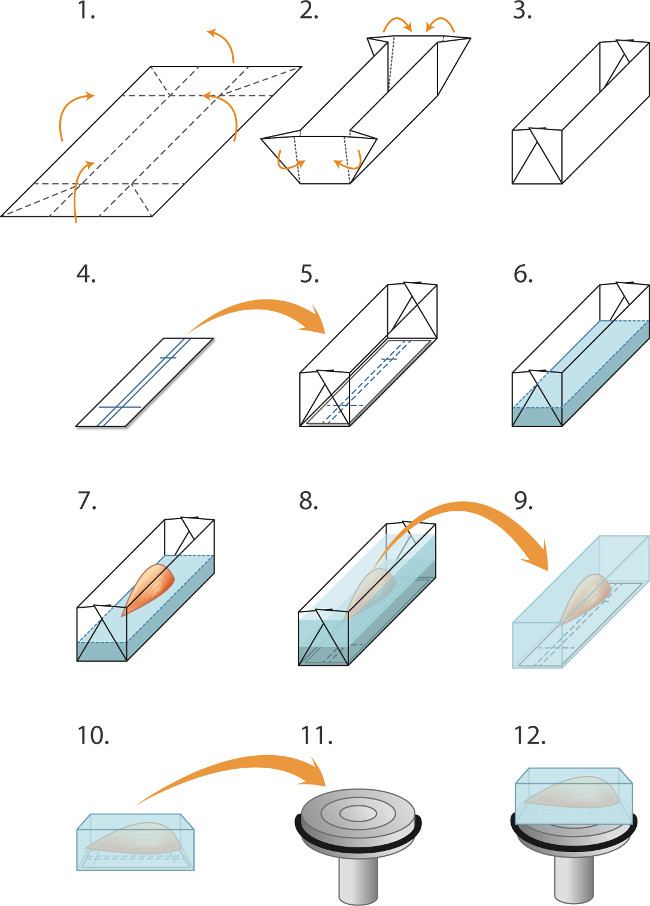
Figura 1. Incorporación y músculos Congelar cryosectioning (1-5) Preparación de moldes ('barcos') antes de la congelación de un lote de músculos:. (1) El papel de aluminio se corta en rectángulos (2,0 x 3,0 cm) y (2). Doblado como acusado de crear un molde / barco (3). (4) Los rectángulos de papel inmunotransferencia nitrocelulosa se cortan para caber en el molde y un bolígrafo se utiliza para descartar líneas para orientar el músculo. (5) El rectángulo de nitrocelulosa se coloca en el molde. (6-8) Incorporación y la congelación de los músculos: (6) Cryostat fluido matriz incrustación se vierte suavemente en el molde (en la parte superior de la nitrocelulosa) a una profundidad de 2 mm. (7) unas pinzas finas se usan para bajar el músculo, por su tendón en la matriz de la incrustación, en alineación con el nitrocelluperder resoluciones, (8) matriz incrustación adicional se vierte suavemente en el molde para cubrir el músculo, teniendo cuidado de evitar la creación de burbujas. El músculo incrustado es entonces congelado rápidamente, sellado en tubos y se almacenaron a -80 ° C como se describe en el texto. (9-12) Preparación para cryosectioning: (9) Bellas fórceps se utilizan para despegar el molde de aluminio y el bloque congelado se coloca entonces en el -20 ° C cámara de criostato, (10) Las marcas en la nitrocelulosa se utilizan para alinear el músculo paralela a la cara de la pinza de sujeción (11) para la sección longitudinal (12). Una gota de medio de crio-incrustación de líquido se utiliza para unir el bloque al plato frío. Fórceps Blunt se utilizan para manipular el bloque de músculo congelado. Para obtener secciones transversales, el bloque se monta en lugar de modo que el músculo es perpendicular a la superficie del mandril (no mostrado).

Figura 2. In caras de NMJs desde el músculo tibial anterior de 2 meses de edad ratones C57BL6J femeninos. El músculo fresca era congelaron rápidamente y las secciones fijas en la diapositiva como se describe en el paso 3.2. Imágenes de proyección máxima intensidad de z-pilas se obtuvieron como se describe en el primer protocolo. Tinción (A) rojo-BGT revela una sola placa motora consiste en dos conjuntos de AChR-ricos canaletas postjunctional primarios (es opinión de la cara). (B) Sinaptofisina tinción con anticuerpo secundario conjugado con FITC revela la terminación nerviosa presináptica, ocupando los canales primarios sinápticas. Una parte del axón pre-terminal también es visible en la parte superior del panel. (C + D) Un ejemplo de una imagen pobre NMJ calidad de un ratón joven y sana. La tinción es débil y los límites de las especializaciones de pre y post-sinápticas son borrosas. Esto se atribuyó a deficiencias en tque el procesamiento del tejido y / o tiempo inadecuado permitió la incubación del anticuerpo primario. La barra de escala en el panel D representa 10 micras. Por favor, haga clic aquí para ver una versión más grande de esta figura.

Figura 3. Pasos en el procesamiento de imágenes en la cara NMJ (como indica el protocolo 1). (A & B) MIP imágenes de tono continuo original Mostrar fluorescencia roja-BGT revelando AChR e inmunofluorescencia verde para sinaptofisina, respectivamente. (C) la tinción de AChR después de la conversión a una imagen de escala de grises de 8 bits y el uso de la herramienta de polígono para delinear la placa terminal (delgada línea amarilla). (D) línea limítrofe placa terminal lleva a la imagen sinaptofisina verde. (E) Applicación de un comando de umbral mínimo de intensidad para crear una imagen binaria que aísla supraumbral rojo-BGT (CADH) fluorescencia. La secuencia de imágenes ImageJ de interfaz de usuario se muestra a la izquierda de las imágenes. Sinaptofisina imagen binaria después de la aplicación de un umbral separado (F). (G) Identificación de suprathreshold discreta dominios AChR-ricos dentro de la placa terminal mediante la aplicación de la orden partículas Analizar a la imagen de color rojo-BGT binario. El ImageJ correspondiente (izquierda del panel (G) muestra los datos de entrada requeridos. El tamaño mínimo de las zonas de píxeles suprathreshold requeridos debe ser ingresado. (H) Identificación de las regiones de solapamiento de la sinaptofisina binario y las imágenes CADH. Overlap está representada por píxeles blancos . ImageJ capturas de pantalla de la interfaz de usuario (por debajo del panel H) mostrar los pasos para llegar a la imagen de superposición binario. Los valores de umbral de intensidad mínimos elegidos para cada fluorescencia channel debe introducirse dentro de la ventana 'Colocalización'. La barra de escala representa 10 micras. Por favor, haga clic aquí para ver una versión más grande de esta figura.

Figura 4. Ejemplos de secciones transversales ópticos utilizados para comparar las intensidades fluorescentes relativas (protocolo 2). El músculo se congeló rápidamente y las secciones fijas en el portaobjetos como se describe en el paso 3.2. (A & B) Una sola placa de extremo doble etiquetado con mucho -red-BGT (CADH; se muestra aquí en azul pseudocolor) y FITC-anti-RapSyn ilustra la co-localización de estas dos proteínas que interactúan en la membrana postsináptica (C & D) Dos placas terminales de las fibras musculares adyacentes pantalla co-localizado AC. RRHH y almizcle. (E & F) Una placa de extremo de doble etiquetado para CADH y -dystroglycan (-DG). El -DG extiende derecha alrededor del perímetro de la fibra muscular, pero se enriquece en la placa terminal (barra de escala en F, para los paneles AF: 25 micras) (G - I) Aislar una placa terminal para la medición de intensidad (G) Un campo microscópico típico,.. que contiene tres placas terminales teñidas con rojo lejano-BGT (barra de escala en el panel G: 40 m (H) Una imagen ampliada de la placa terminal en caja (I) La misma placa terminal convierte en una imagen en escala de grises de 8 bits y delineó con el polígono.. . herramienta de ImageJ (línea amarilla delgada) Intensidad media de fluorescencia se mide dentro de esta línea de límite. (barra de escala para H & I: 10 micras) Haga clic aquí para ver una versión más grande de esta figura.

Figura 5. Influencia de la calidad de imagen en la evaluación de área sináptica. (A y B) de alta calidad en imágenes de la cara de un NMJ saludable de un viejo ratón 2 meses visto en el rojo-BGT y canales de fluorescencia anti-sinaptofisina. (A 'y B ') La intensidad de fluorescencia correspondientes perfiles a la línea trazada a través de la placa terminal para A y B, respectivamente. La línea roja punteada horizontal indica el umbral mínimo utilizado para crear la imagen binaria. (C + D) Placa terminal de un ratón ancianos. Tinción sinaptofisina placa terminal generalmente es menos intenso. (C 'y D') perfiles de intensidad muestran un alto nivel de extrasynaptic (línea de base) de fluctuación de fluorescencia en el (FITC) canal de sinaptofisina (backsuelo) que afecta a la determinación de un umbral adecuado. Mucho de esto es de amplio espectro autofluorescencia del tejido. Las barras de escala representan 10 micras. Por favor, haga clic aquí para ver una versión más grande de esta figura.

Figura 6. La variabilidad de las zonas sinápticas entre NMJs dentro de un músculo y entre los ratones. (A) Diagramas de dispersión muestran la AChR-rica área total de las placas terminales del músculo tibial anterior de siete ingenuas 2 meses de edad C57BL / 6J obtenidos por el autor NT. Cada símbolo representa una placa terminal. Cada barra representa la media ± SD para placas terminales incluidos en la muestra de un ratón. (B) Diagramas de dispersión que muestra el área sinaptofisina ricos por las mismas placas terminales. (A ') (B ') Distribución de frecuencias para el área sinaptofisina ricos de placas terminales (datos agrupados). Haga clic aquí para ver una versión más grande de esta figura.

Figura 7. Efecto del tamaño de la muestra en las estimaciones de la intensidad de fluorescencia placa terminal. Secciones ópticas transversales se utilizaron para medir la intensidad de fluorescencia (en unidades arbitrarias) en 40 placas de extremo 60 desde el músculo tibial anterior de un 2 meses de edad ratón sano. Promedios acumulados se trazan contra el número de placas terminales incluidas en la media. (A) Placa terminal intensidad de fluorescencia roja-BGT obtiene autor AV. (B) Anti-synaptophysin intensidad inmunofluorescencia para las mismas placas terminales como en el panel A. intensidad (C) Placa terminal rojo lejano-BGT fluorescencia obtenida a partir de una segunda muestra de músculo por autor NG. (D) Anti-RapSyn intensidad de inmunofluorescencia de las mismas placas terminales como en el panel C.
| Estudio | Músculo | número de ratones | Área AChR | Área Sinaptofisina | Área de solapamiento |
| media ± DE (μm2) | media ± DE (μm2) | media ± DE (μm2) | |||
| Morsch et al. (2012) 1 | gastrocnemio | 3 | 181 ± 7 | 163 ± 24 | |
| diafragma | 3 | 130 ± 41 | 102 ± 21 | 76 ± 26 | |
| Morsch et al. (2013) 1 | tibial anterior | 3 | 166 ± 26 | 117 ± 21 | nd |
| Cheng et al. (2013) 2 | tibial anterior | 4 | 226 ± 18 | 150 ± 44 | 110 ± 34 |
| Tse (inédito) 2 | tibial anterior | 7 | 213 ± 27 | 157 ± 23 | 111 ± 11 |
| 1 imagen sobre un microscopio Zeiss LSM 510 Meta pero con ganancia fija y los niveles de compensación. Mediciones de umbrales y zonas sinápticas utilizan Metamorph software por MM. | |||||
| 2 proyectado sobre un microscopio Leica DM IRE2 y analizados siguiendo el protocolo actual por el primer autor indica, ciego al grupo de tratamiento. | |||||
| nd no determinado. | |||||
Tabla 1. zonas sinápticas para NMJs de 2 meses de edad hembra C57BL / 6J (controles sanos)
| Estudio | Músculo | FRET eficiencia (%) * | Rango (%) |
| (Media ± SEM) | |||
| Brockhausen et al. (2008) | tibial anterior | 24 ± 1 | n / a |
| Cole et al. 2010 | tibial anterior | 26 ± 1 | 22 - 30 |
| Morsch (inédito) | diafragma | 37 ± 1 | 24-47 |
| Ghazanfari (inédito) | tibial anterior | 30 ± 1 | 20 - 45 |
| * FRET entre (radio rojo-BGT y rojo lejano-BGT Förster para FRET par = 51A (Life Technologies). | |||
| datos NA No disponible | |||
Tabla 2. eficiencias para AChR FRET a partir de ratones jóvenes adultos ratones C57BL6J
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Los protocolos descritos aquí nos han permitido medir y cuantificar los cambios en las propiedades de la UNM en toda una serie de condiciones, incluyendo los estados normales de envejecimiento y la enfermedad. Los métodos descritos para en face imágenes NMJ permitirán a los investigadores comparan el área de especialidades pre y postsinápticos y el área de superposición sináptica / alineación. Para comparar la intensidad relativa de las proteínas pre y postsinápticos el segundo protocolo, que utiliza secciones ópticas transversales, se prefiere. El tercer protocolo a prueba específicamente para los cambios en la proximidad de embalaje de AChR en la membrana postsináptica.
Controles de especificidad son vitales en la microscopía de inmunofluorescencia. Al utilizar cualquier anticuerpo primario para inmunofluorescencia indirecta es necesario para asegurar primero que se une específicamente a su proteína diana en secciones musculares. Diferentes tipos de procesamiento del tejido y la fijación pueden alterar la especificidad de diferencialmenteanticuerpos. Es importante confirmar que la tinción de inmunofluorescencia (por ejemplo RapSyn) realmente se concentra con AChR en la placa motora. Secciones de control negativas también deben ser inspeccionados para asegurar que la unión del anticuerpo es específico. Por ejemplo, el mejor control negativo para inmunofluorescencia RapSyn sería secciones de RapSyn - / - ratones. Éstas no deberán presentar manchas placa terminal con anti-RapSyn. Fluorescencia no específica también puede surgir de los productos químicos fluorescentes endógenas en el tejido (autofluorescencia) o de la unión no específica por el conjugado anticuerpo secundario fluorescente. Tal fluorescencia a menudo se ve agravada por la fijación de aldehído. Además, la tinción TRITC-BGT de placas terminales a veces puede ser detectada en el canal de fluorescencia FITC y esto fluorescente de purga a través podría ser confundido con inmunofluorescencia específica FITC. Para protegerse de las tres últimas formas de fluorescencia no específica, cada lote de diapositivas que se tiñó debe incluir algún 'n'secciones de control de anticuerpos o-primaria (pasos 3.7 y 4.6). Imágenes de placas terminales de estas secciones de control deben ser comparados con los de las diapositivas experimentales para asegurar que la tinción de inmunofluorescencia indirecta de NMJs realmente refleja la unión del anticuerpo primario.
Transversal secciones confocales son particularmente útiles para la evaluación de las diferencias en la intensidad relativa de la inmunotinción en la sinapsis. En las secciones transversales confocal es más fácil juzgar precisa co-localización de las proteínas sinápticas. El perfil de placa terminal en forma de media luna representa sólo un corte a través de muestra de la UNM en cuestión. Sin embargo, el fondo (extrasynaptic) fluorescencia es generalmente menor en comparación con la cara en las imágenes z-proyección. Por lo tanto, puede ser más fácil de discriminar inmunotinción 'real' (específico) y establecer la ganancia confocal fija y valores de desplazamiento utilizando secciones transversales ópticas 13-15,18. Por ejemplo, en un modelo de ratón de la miastenia gravis (where placa terminal tinción AChR se reduce notablemente) placas terminales están claramente delineadas en las secciones transversales ópticas 18,21. Las diferencias en la intensidad media de fluorescencia a la NMJ es probable que reflejen la densidad alterada de la proteína objetivo dentro de la especialización sináptica. Una advertencia es que, en algunas situaciones, un cambio estructural en la proteína diana u oclusión de la unión del anticuerpo por las proteínas vecinas podría explicar intensidad de la tinción alterada.
El diseño de experimentos requiere cierta consideración. En muchos casos el experimento tendría como objetivo para probar el impacto de un transgén, desmontables gen o estado de enfermedad en el tamaño de la NMJ. El grupo muestra experimental podría compararse a los jóvenes (de tipo salvaje) ratones sanos del mismo sexo y los antecedentes genéticos. Los valores de referencia para la zona de sinaptofisina placa terminal, CADH y superposición sináptica durante varios músculos se dan en la Tabla 1. Tamaño de la muestra dependerá del grado de animalesvariación -a-animal dentro de los grupos de tratamiento y el tamaño del efecto (diferencia de medias para el experimental frente los grupos de control por desviación estándar). Cuando el análisis se limita a imágenes de buena calidad se encontró un alto grado de consistencia en los medios de la muestra para las zonas de placa terminal entre los 2 meses de edad ratones C57BL6J femeninos sanos (Figura 6 A y B). Así, fue posible demostrar significativas reducciones del 30-40% en el área sináptica en ratones inyectados con IgG de miastenia anti-MuSK positivos gravis pacientes, en comparación con los controles con un tamaño de muestra de tres ratones 17,20,32. Ratones de edad avanzada muestran una mayor variación de animal a animal en los parámetros de la placa terminal de ratones jóvenes 22. En consecuencia experimentos con ratones viejos pueden requerir muestras de mayor tamaño.
Si la principal preocupación es medir el tamaño de la placa terminal en la cara entonces la ganancia y ajustes de nivel de compensación debe ser optimizada para cada NMJ individual. IndivNMJs idual pueden variar considerablemente en el brillo de AChR y tinción sinaptofisina, en particular cuando se examinan estados de enfermedad. Además, la intensidad de la fluorescencia extra-sináptica (no específica) es a menudo mayor y más variable en los músculos de los animales de envejecimiento, en comparación con los de los animales jóvenes sanos (Figura 5C y D). El 1-256-escala de grises que debe recurrirse a fin de maximizar la información tonal que se retiene en las imágenes finales. Esto implicará el ajuste de la ganancia y niveles para cada NMJ para el cual un z-pila se debe desechar por offset. La Figura 5D muestra un ejemplo de una imagen NMJ donde la información tonal podría ser crítico en la definición de los límites de la zona de pre- y post- especializaciones sinápticas.
Las mediciones de las zonas sinápticas se pueden aplicar a diferentes preparaciones de músculo y experimentos. La mayor parte de nuestras mediciones de zonas sinápticas han empleado cryosections longitudinales de presión músculos congelados. Congelación de la muscular antes de la fijación mantiene la antigenicidad de una amplia gama de proteínas. Cuando compatible con el antígeno, la fijación de paraformaldehído y la infiltración de sacarosa antes de cryosectioning (paso 2.1) puede proporcionar una mejor preservación de la estructura NMJ. Preservación estructural óptima podría obtenerse mediante perfusión cardíaca con paraformaldehído. Los artefactos de congelación y seccionamiento pueden evitarse por completo por placas terminales de etiquetado en la superficie de las NMJs musculares y de imagen intactas en fascículos burlado desde el músculo fija 21. Independientemente de la preparación, los procedimientos de muestreo, las imágenes y el área cuantificación se mantienen sin cambios (protocolo de los pasos 4-5). La aplicación coherente de los protocolos de muestreo, de imágenes y análisis de ciegos (utilizando diferentes operadores, diferentes muestras de ratones y tiempos diferentes), puede dar lugar a valores medios bastante reproducibles (comparar Cheng et al. Y Tse resultados de la Tabla 1).
"> Placas de extremo se han descrito como convertirse 'fragmentado' en una variedad de estados de enfermedad. Por ejemplo, en el envejecimiento de los músculos de ratón, la degeneración esporádica de una fibra muscular (seguido de su regeneración) dio lugar a la remodelación de la placa terminal como pretzel-AChR placa para formar múltiples agrupaciones más pequeñas AChR 6. En los ratones inyectados con IgG de miastenia gravis anti-MuSK pacientes, la fragmentación de la placa terminal era bastante diferente. La placa terminal AChR pretzel en gran medida dispersa, dejando atrás una constelación de pequeños (<4 m 2) microagregados AChR ' '20,21. Estos dos ejemplos ponen de relieve la necesidad de comparar las distribuciones de tamaño para los clústeres AChR en placas terminales de control frente a los animales de experimentación 21.Otros métodos se han reportado para evaluar el área sináptica o intensidad de la tinción en la UNM. Placas motoras veces se pueden plegar de manera que las imágenes z-proyección bidimensional utilizados aquí pueden subestimar synáreas APTIC. Reconstrucciones confocal tridimensionales podrían proporcionar medidas más precisas si el área sináptica absoluta debe ser definido 33. Una ventaja clave del protocolo z-proyección descrito aquí, sin embargo, es su simplicidad relativa, que ha permitido un gran número de placas terminales a ser medido a partir de múltiples grupos de tratamiento y la identificación fiable de los cambios potenciales. El protocolo para la comparación de intensidades de tinción de placa terminal puede ser adaptado para el estudio de los cambios en los niveles de muchos diferentes proteínas sinápticas. El método es limitada, sin embargo, por la exigencia de que todas las muestras se procesaron para inmunofluorescencia entonces fotografiado durante la misma sesión confocal. Un estudio reciente realizado por Yampolsky et al. 5 describe un método para medir la densidad de la placa terminal AChR que podrían ayudar a superar esta limitación. En este estudio, los mismos campos de placas terminales fueron imágenes a varios ajustes de potencia láser diferentes. La pendiente de la relación entre la potencia del láser y rhodamiintensidad de fluorescencia ne-BGT se utilizó para evaluar los cambios relativos en la densidad de AChR en placas terminales en diferentes ratones 5. Este método podría ser útil para comparar la intensidad de AChR en muestras fotografiadas en diferentes momentos durante el curso de un estudio prolongado.
AChR-AChR FRET proporciona información específica y complementaria sobre la organización de AChR placa terminal. Electron autorradiografía microscópica utilizando 125. I-α-BGT ha mostrado AChR a envasar herméticamente con una densidad planar de 10 4 m -2 inmediatamente debajo de cada sitio presináptica de la liberación del transmisor, mientras contiguas repliegues de membrana contienen densidades mucho más bajas de AChR 34. AChR-AChR FRET hace que sea relativamente fácil de detectar cambios (sub-microscópicos) en el embalaje AChR. Una reducción en la eficiencia de FRET refleja una redistribución sub-microscópica de AChR en la membrana postsináptica que podrían no ser detectadas por un cambio en la intensidad media de fluorescencia BGT. F múltipleactores podrían causar un cambio en la eficiencia de FRET. Estos incluyen la separación donante-aceptor y orientaciones relativas, así como el entorno molecular 35,36. Una reducción en la eficiencia de FRET placa terminal, posiblemente, podría surgir de un cambio en la geometría de la red AChR. Sin embargo, lo más probable sería debido a una reducción en el porcentaje de AChR que se colocan en la nano-escala de celosía molecular postsináptica 14.
La pérdida de la conexión entre las neuronas motoras y las fibras musculares parece ser la causa inmediata de la debilidad muscular en la enfermedad de la neurona motora y en el envejecimiento sedentaria 9,22,23. Métodos y parámetros compartidos para NMJs medición deben facilitar a los diferentes grupos de investigación para comparar y hallazgos de contraste publicada. El intercambio de protocolos detallados (y futuras mejoras en ellos) puede ayudar a acelerar el progreso en la comprensión de los mecanismos de mantenimiento NMJ y cómo puede verse afectada en estados de enfermedad.
Subscription Required. Please recommend JoVE to your librarian.
Materials
| Name | Company | Catalog Number | Comments |
| Scanning confocal microscope | Leica | DM 2000 with TCS SP2 system | Most scanning confocal microscopes should be suitable. |
| Zeiss | LSM 510 Meta | ||
| Leica | SPE-II | ||
| Alexa555-a-bungarotoxin (red-BGT) | Life technologies | B35451 | Used for labelling AChRs |
| Alexa647-α-bungarotoxin (far-red-BGT) | Life technologies | B35450 | Far red fluorescence: barely visible through the eyepiece |
| rabbit anti-synaptophysin | Life technologies | 18-0130 | Different batches of primary antibody differ in effective working dilution |
| FITC-anti-rapsyn mab1234 | Milipore | FCMAB134F | Monoclonal antibody conjugated to FITC |
| FITC-donkey anti-rabbit IgG | Jackson | 711-095-152 | Polyclonal secondary antibodies can vary in quality according to source and batch |
| Optimal Cutting Temperature compound (O.T.C.) | ProSciTech | IA018 | Cryostat embedding matrix for freezing muscles |
| DABCO | Sigma | 10981 | Mounting medium that slows photobleaching of fluorophores |
References
- Schmidt, N., et al. Neuregulin/ErbB regulate neuromuscular junction development by phosphorylation of α-dystrobrevin. J Cell Biol. 195, 1171-1184 (2011).
- Amenta, A. R., et al. Biglycan is an extracellular MuSK binding protein important for synapse stability. J Neurosci. 32, 2324-2334 (2012).
- Samuel, M. A., Valdez, G., Tapia, J. C., Lichtman, J. W., Sanes, J. R. Agrin and Synaptic Laminin Are Required to Maintain Adult Neuromuscular Junctions. PLOS ONE. 7, e46663 (2012).
- Valdez, G., et al. Attenuation of age-related changes in mouse neuromuscular synapses by caloric restriction and exercise. Proc Natl Acad Sci (USA). 107, 14863-14868 (2010).
- Yampolsky, P., Pacifici, P. G., Witzemann, V. Differential muscle-driven synaptic remodeling in the neuromuscular junction after denervation). Eur J Neurosci. 31, 646-658 (2010).
- Li, Y., Lee, Y., Thompson, W. J. Changes in Aging Mouse Neuromuscular Junctions Are Explained by Degeneration and Regeneration of Muscle Fiber Segments at the Synapse. J Neurosci. 31, 14910-14919 (2011).
- Zhu, H., Bhattacharyya, B. J., Lin, H., Gomez, C. M. Skeletal muscle IP3R1 receptors amplify physiological and pathological synaptic calcium signals. J Neurosci. 31, 15269-15283 (2011).
- Valdez, G., Tapia, J. C., Lichtman, J. W., Fox, M. A., Sanes, J. R. Shared resistance to aging and ALS in neuromuscular junctions of specific muscles. PLoS ONE. 7, e34640 (2012).
- Perez-Garcia, M. J., Burden, S. J. Increasing MuSK Activity Delays Denervation and Improves Motor Function in ALS Mice. Cell reports. 2, 1-6 (2012).
- Klooster, R., et al. Muscle-specific kinase myasthenia gravis IgG4 autoantibodies cause severe neuromuscular junction dysfunction in mice. Brain. 135, 1081-1101 (2012).
- Pratt, S. J., Shah, S. B., Ward, C. W., Inacio, M. P., Stains, J. P., Lovering, R. M. Effects of in vivo injury on the neuromuscular junction in healthy and dystrophic muscles. J Physiol. 591, 559-570 (2013).
- Landis, S. C., et al. A call for transparent reporting to optimize the predictive value of preclinical research. Nature. 490, 187-191 (2012).
- Gervásio, O. L., Phillips, W. D. Increased ratio of rapsyn to ACh receptor stabilizes postsynaptic receptors at the mouse neuromuscular synapse. J Physiol. 562, 673-685 (2005).
- Gervásio, O. L., Armson, P. F., Phillips, W. D. Developmental increase in the amount of rapsyn per acetylcholine receptor promotes postsynaptic receptor packing and stability. Dev Biol. 305, 262-275 (2007).
- Brockhausen, J., Cole, R. N., Gervásio, O. L., Ngo, S. T., Noakes, P. G., Phillips, W. D. Neural agrin increases postsynaptic ACh receptor packing by elevating rapsyn protein at the mouse neuromuscular synapse. Dev Neurobiol. 68, 1153-1169 (2008).
- Cole, R. N., Reddel, S. W., Gervásio, O. L., Phillips, W. D. Anti-MuSK patient antibodies disrupt the mouse neuromuscular junction. Ann Neurol. 63, 782-789 (2008).
- Morsch, M., Reddel, S. W., Ghazanfari, N., Toyka, K. V., Phillips, W. D. Muscle Specific Kinase autoantibodies cause synaptic failure through progressive wastage of postsynaptic acetylcholine receptors. Exp Neurol. 237, 237-286 (2012).
- Cole, R. N., Ghazanfari, N., Ngo, S. T., Gervasio, O. L., Reddel, S. W., Phillips, W. D. Patient autoantibodies deplete postsynaptic Muscle Specific Kinase leading to disassembly of the ACh receptor scaffold and myasthenia gravis in mice. J Physiol. 588, 3217-3229 (2010).
- Viegas, S., et al. Passive and active immunization models of MuSK-Ab positive myasthenia: Electrophysiological evidence for pre and postsynaptic defects. Exp Neurol. 234, 506-512 (2012).
- Morsch, M., Reddel, S. W., Ghazanfari, N., Toyka, K. V., Phillips, W. D. Pyridostigmine but not 3,4-diaminopyridine exacerbates ACh receptor loss and myasthenia induced in mice by Muscle Specific Kinase autoantibody. J Physiol. 591, 2747-2762 (2013).
- Ghazanfari, N., Morsch, M., Reddel, S. W., Liang, S. X., Phillips, W. D. Muscle Specific Kinase autoantibodies suppress the MuSK pathway and ACh receptor retention at the mouse neuromuscular junction. J Physiol. 592, 2881-2897 (2014).
- Cheng, A., Morsch, M., Murata, Y., Ghazanfari, N., Reddel, S. W., Phillips, W. D. Sequence of age-associated changes to the mouse neuromuscular junction and the protective effects of voluntary exercise. PLoS One. 8, e67970 (2013).
- Schaefer, A. M., Sanes, J. R., Lichtman, J. W. A compensatory subpopulation of motor neurons in a mouse model of amyotrophic lateral sclerosis. J Comp Neurol. 490, 209-219 (2005).
- Kilkenny, C., Browne, W. J., Cuthill, I. C., Emerson, M., Altman, D. G. Improving bioscience research reporting: the ARRIVE guidelines for reporting animal research. PLos Biol. 8, e1000412 (2010).
- Shimizu, S. Routes of Administration. The Laboratory Mouse. Hedrich, H. J., Bullock, G. , Elsevier. (2004).
- Chiasson, R. B. Laboratory anatomy of the white rat. , Brown. Dubuque, Iowa. (1988).
- Gage, G. J., Kipke, D. R., Shain, W. Whole Animal Perfusion Fixation for Rodents. J. Vis. Exp. (65), e3564 (2012).
- Mitra, A. K., Stroud McCarthy, M. P., M, R. Three-dimensional structure of the nicotinic acetylcholine receptor and location of the major associated 43-kD cytoskeletal protein, determined at 22A by low dose electron microscopy and x-ray diffraction to 12.5A. J Cell Biol. 109, 755-774 (1989).
- Paas, Y., et al. Electron microscopic evidence for nucleation and growth of 3D acetylcholine receptor microcrystals in structured lipid-detergent matrices. Proc. Natl Acad. Sci. (USA). 100, 11309-11314 (2003).
- Samson, A. O., Scherf, T., Eisenstein, M., Chill, J. H., Anglister, J. The mechanism for acetylhcoline receptor inhibition by α-neurotoxins and species-specific resistance to α-bungarotoxin revealed by NMR). Neuron. 35, 319-332 (2002).
- Ghazanfari, N., et al. Muscle Specific Kinase: Organiser of synaptic membrane domains. Int J Biochem Cell Biol. 43, 295-298 (2011).
- Ghazanfari, N., Morsch, M., Tse, N., Reddel, S. W., Phillips, W. D. Effects of the β2-adrenoceptor agonist, albuterol, in a mouse model of anti-MuSK myasthenia gravis. PLoS ONE. 9, e87840 (2014).
- Prakash, Y. S., Miller, S. M., Huang, M., Sieck, G. C. Morphology of diaphragm neuromuscular junctions on different fibre types. J Neurocytol. 25, 88-100 (1996).
- Salpeter, M. M., Harris, R. Distribution and turnover rate of acetylcholine receptors throughout the junction folds at a vertebrate neuromuscular junction. J Cell Biol. 96, 1781-1785 (1983).
- Soper, S. A., Nutter, H. L., Keller, R. A., Davis, L. M., Shera, E. B. The photophysical constants of several fluorescent dyes pertaining to ultrasensitive fluorescence spectroscopy. Photochem Photobiol. 57, 972-977 (1993).
- Panchuk-Voloshina, N., et al. Alexa dyes, a series of new fluorescent dyes that yield exceptionally bright, photostable conjugates. J Histochem Cytochem. 47, 1179-1188 (1999).
