In order to choose a method to deplete thymic lobes of endogenous thymocytes allowing the best development of colonizing progenitors, we compared the levels of T cell reconstitution in thymic lobes colonized after irradiation or a 5-day deoxy-guanosine (d-Gua) treatment. The results show that while there is no difference at day 9 of culture, irradiated lobes contained more T cells than those treated with d-Gua, at day 12. Thus, irradiation is more appropriate than d-Gua treatment to obtain T cell development after thymic colonization (Figure 1). To study the developmental potential of E13 and E18 TSP, we colonized E14 irradiated thymic lobes with a mixture of equal numbers of the two types of progenitors. The results show that E13 TSPs give rise to less thymocytes and less DP than E18 TSP but, in contrast, E13 TSP generate DETC and higher frequencies of CD3+ mature cells (Figure 2). To analyze the in vivo potential of E13 and E18 TSP, thymic lobes colonized with each type of progenitors were grafted under the kidney capsule of CD3-/- recipients. The results show that, consistent with the FTOC, E13 TSP gave rise to T cells faster than E18 progenitors but the number of circulating T cells was significantly lower (Figure 3).

Figure 1: T cell development is more efficient in irradiated than in deoxyguanosine treated, colonized thymic lobes. E14 thymic lobes (CD45.2) were either irradiated with 30 Gy or treated for 5 days with deoxy-guanosine (d-Gua). Thymic lobes were then colonized with 1000 Lin–CD117+Sca-1+ (LSK) E14 FL cells isolated from CD45.1/2 embryos, in hanging drop for 48 hr and cultured on a filter. No differences were observed in the efficiency of endogenous thymocyte depletion between the two groups of thymic lobes. Developing thymocytes stained with antibodies against CD45.1, CD45.2, CD3, Vγ5, Vδ1 and CD4 were analyzed at day 9 and 12 by FACS. Please click here to view a larger version of this figure.

Figure 2: In contrast to E18, E13 TSPs generate Vγ5Vδ1 DETC. CD45.2 thymic lobes were irradiated and colonized for 48 hr with a mixed cohort of 500 E13 TSP from CD45.1/2 embryos and 500 E18 TSP from CD45.1 embryos. Under these experimental conditions, E13 and E18 TSP develop in the same environment and differences in the rate of differentiation and mature T cell subsets observed after culture can only reflect differences in cell intrinsic biologic properties. After 12 days in culture thymocytes from individual lobes were analyzed by flow cytometry after staining with antibodies recognizing CD45.1, CD45.2, CD4, CD8, CD3, Vγ5, Vδ1. (A) Panels show the numbers of cells recovered in each lobe. (B) Representative flow cytometry profiles. Please click here to view a larger version of this figure.
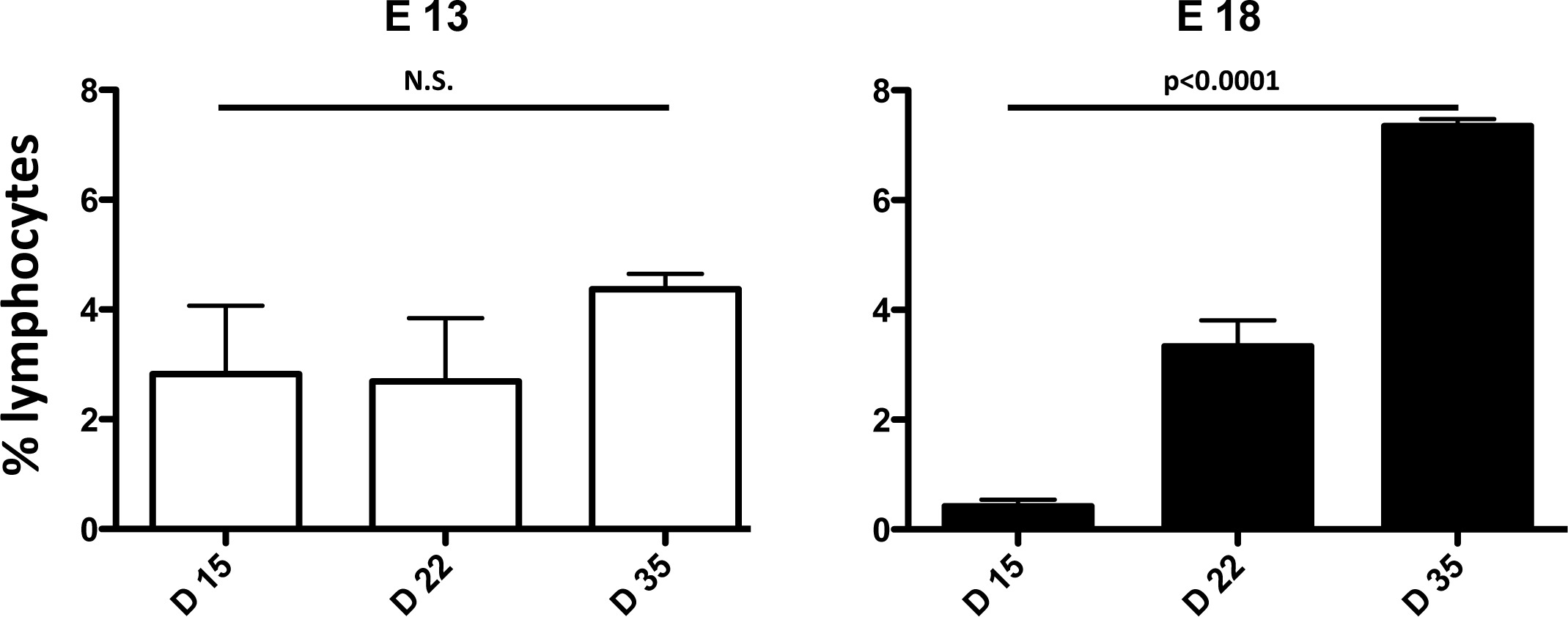
Figure 3: E13 TSPs develop faster than E18 TSPs. CD45.2 thymic lobes were irradiated and colonized for 48 hr with 500 E13 TSP or 500 E18 TSP from CD45.1 embryos. CD3-/- mice were grafted with 4 thymic lobes colonized with either E13 or E18 TSPs. Peripheral blood was collected at weekly intervals and analyzed by FACS for donor (CD45.1) CD3+ T cells. Please click here to view a larger version of this figure.
| step 1 | Mating mice to obtain E18 embryos (day -21); mating mice to obtain E14 embryos (day -17); mating mice to obtain E13 embryos (day -16) |
| step 2a | Dissection of the embryos, day -2 |
| step 2b | Irradiation of E14 thymic lobes, day -2 |
| step 2c | Preparing, staining, and sorting cells from E13 and E18 thymic lobes, day -2 |
| step 2d | Hanging drop culture, day -2 |
| step 3a | Fetal Thymic Organ Culture, day 0 |
| step 3b | Graft under the kidney capsule, day 0 |
| step 4 | FTOC: flow cytometry analysis, day 12 |
| step 5 | Graft: weekly flow cytometry analysis of circulating T cells, days 15, 22, 35 |
Table 1: The 5-step procedure followed in the experiment. Time-table of the experiment from the mating of the different mouse strains up to the analysis of the grafted mice. Grafts are performed in 7 week old mice. Taking the time of transplantation as day 0, day -21 is the mating of mice to obtain E18 embryos, day -17 to obtain E14 embryos, day -16 to obtain E13 embryos, and in day -2 the sorting of E13 and E18 TSP, irradiation thymic lobe and hanging drop technique.
| Antibody | Clone Number | Antibody | Clone Number | |
| CD25 | 7D4 | CD19 | 6D5 | |
| CD44 | IM7 | Ter 119 | TER-119 | |
| CD24 | M1/69 | NK1.1 | PK 136 | |
| CD117 | 2B8 | CD11c | HL3 | |
| CD3 | 145-2C11 | Gr1 | RB6-8C5 | |
| CD4 | RM4-5 | GD | eBioGL3 | |
| CD8 | 53-6.7 | Sca-1 | D7 | |
| CD135 | A2F10 | CD45.2 | 104 | |
| CD127 | A7R34 | Ly5.1 | A20 |
Table 2: Antibodies and clone numbers.
