

Summary
Este protocolo detalha as etapas, custos e equipamentos necessários para gerar Escherichia coli-com base em extratos de células e implementar em vitro reações de síntese de proteínas dentro de 4 dias ou menos. Para aproveitar a natureza flexível desta plataforma para aplicações de amplas, podemos discutir as condições de reação que podem ser adaptadas e otimizadas.
Abstract
Nos últimos 50 anos, a síntese de proteína sem célula (CFPS) emergiu como uma poderosa tecnologia para aproveitar a capacidade transcricional e translacional de células dentro de um tubo de ensaio. Por obviating a necessidade de manter a viabilidade da célula e eliminando a barreira celular, CFPS tem sido fundamental para aplicações emergentes em biomanufacturing de proteínas tradicionalmente desafiadores, bem como aplicações em prototipagem rápida para engenharia metabólica e genômica funcional. Nossos métodos para implementar um Escherichia coli-baseado em plataforma CFPS permitir que novos usuários acessar muitos desses aplicativos. Aqui, descrevemos os métodos para preparar o extrato com o uso de meios enriquecidos, frascos perplexos e um método reprodutível de lise celular baseado em sonication ajustáveis. Este extrato pode ser usado na expressão de proteínas capaz de produzir 900 µ g/mL ou mais de proteína fluorescente verde super pasta (sfGFP) em apenas 5 h de instalação experimental para análise de dados, dado que os estoques de reagentes apropriados foram preparados antecipadamente. O custo estimado de inicialização de obtenção de reagentes é $4.500 que sustentarão milhares de reações em um custo estimado de US $0,021 por µ g de proteína produzida ou $0,019 por µ l de reação. Além disso, os métodos de expressão de proteínas espelham a facilidade da instalação do reação vista em sistemas comercialmente disponíveis devido à otimização de misturas prévias de reagente, em uma fração do custo. A fim de habilitar o usuário alavancar a natureza flexível da plataforma para aplicações de amplas CFPS, nós identificamos uma variedade de aspectos da plataforma que pode ser ajustado e otimizado dependendo dos recursos disponíveis e os resultados da expressão da proteína desejado.
Introduction
Síntese de proteína sem célula (CFPS) surgiu como uma tecnologia que desbloqueou uma série de novas oportunidades para a produção de proteína, genómica funcional, engenharia metabólica e muito mais no âmbito do último 50 anos1,2. Comparado ao padrão na vivo plataformas de expressão de proteínas, CFPS fornece três vantagens principais: 1) a natureza sem célula da plataforma permite a produção de proteínas que seriam potencialmente tóxicos ou estrangeiros para a célula3,4 ,5,6; 2) inativação do DNA genômico e a introdução de um modelo de DNA os genes de interesse de codificação canalizar toda a energia sistêmica dentro da reação para a produção das somáticas de interesse; e 3) a natureza aberta da plataforma permite ao usuário modificar e monitorar as condições da reação e a composição em tempo real7,8. Esse acesso direto para a reação suporta o aumento dos sistemas biológicos com químicos expandidos e redox as condições para a produção de novas proteínas e a otimização de processos metabólicos2,9, 10. direto acesso também permite ao usuário combinar a reação CFPS com ensaios de atividade em um sistema de single-pote para teste de compilação de projeto mais rápido ciclos de11. A capacidade de realizar a reação de CFPS em gotas de pequeno volume ou em dispositivos baseados em papel mais suporta os esforços de descoberta do elevado-throughput e prototipagem rápida12,13,14,15 ,16. Como resultado destas vantagens e da natureza de plug and play do sistema, CFPS excepcionalmente permitiu uma variedade de aplicações da biotecnologia tais como a produção de proteínas que são difíceis de solubly expresso na vivo17, 18,19,20, detecção de22,doença21,23, na demanda biomanufacturing18,24 ,25,26,27e educação28,29, os quais mostram a flexibilidade e a utilidade da plataforma livre de célula.
Sistemas CFPS podem ser gerados a partir de uma variedade de bruto lysates de ambas as linhas de células procariotas e eucariotas. Isto permite diversas opções no sistema de escolha, cada uma delas tem vantagens e desvantagens, dependendo da aplicação de juros. Sistemas CFPS também variam muito em produtividade, custo e tempo de preparação. Os mais comumente utilizaram célula extratos são produzidos a partir de células de Escherichia coli , sendo o último mais cost-effective para data ao produzir no mais alto rendimento volumétrico da proteína30, Reticulócito coelho, células de inseto e germe de trigo . Enquanto outros sistemas CFPS podem ser vantajosos para suas máquinas de modificação pós-traducional inata, emergentes aplicações usando o Escherichia coli-baseado em máquinas são capazes de preencher a lacuna, gerando site-specifically fosforilada e glicosilados proteínas na demanda31,32,33,34,35.
Reações de CFPS podem ser executadas em qualquer lote, troca contínua sem célula (CECF) ou fluxo contínuo sem célula (CFCF) formatos. O formato do lote é um sistema fechado, cujo tempo de vida de reação é limitado devido à diminuição de quantidades de reagentes e o acúmulo de subprodutos inibitórios da reação. CECF CFCF métodos aumentam o tempo de vida da reação e, assim, resultam em aumento volumétrico proteína rendimentos em comparação com a reação do lote. Isso é realizado, permitindo que os subprodutos da síntese de proteínas deve ser retirado da embarcação da reação, enquanto novos reagentes são fornecidos durante todo o curso da reação2. No caso de CFCF, a proteína de interesse também pode ser removida da câmara de reação, enquanto no CECF, a proteína de interesse permanece na câmara de reação, composto de uma membrana semi-permeável36,37. Esses métodos são especialmente valiosos na superação pobre rendimento volumétrico de difícil-para-expressam proteínas de interesse38,39,40,41,42, 43. Os desafios na implementação das abordagens CECF e CFCF são que 1) enquanto eles resultam em um uso mais eficiente da maquinaria bio responsável pela transcrição e tradução, necessitam de ser notavelmente maiores quantidades de reagentes que aumenta o custo geral e 2) Eles exigem mais complexa reação de configurações e equipamentos especializados em comparação com o formato de lote44. Para assegurar a acessibilidade para usuários novos, os protocolos descritos foco sobre o formato do lote em volumes de reação de 15 µ l com recomendações específicas para aumentar o volume de reação para a escala de mililitro.
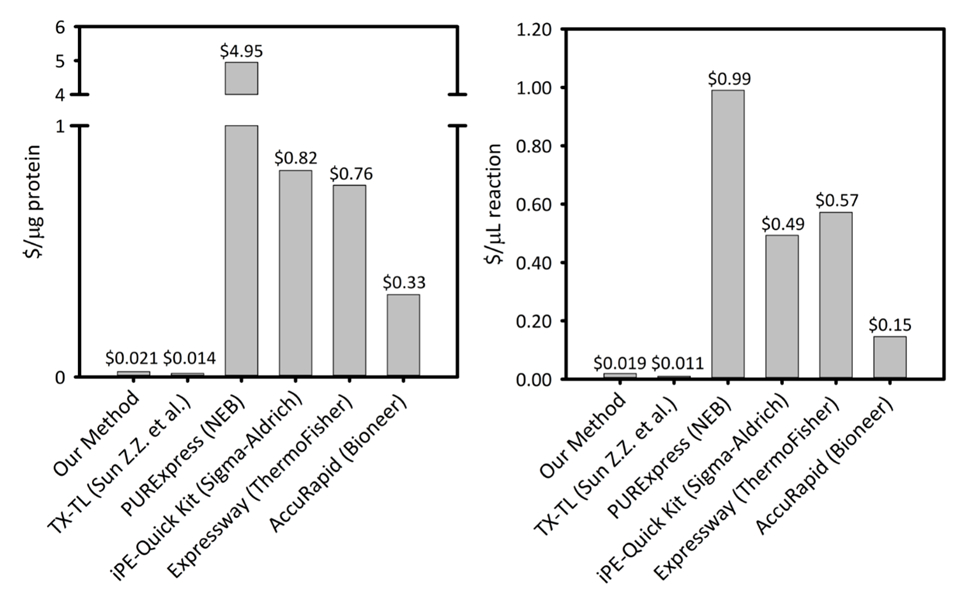
Os métodos aqui apresentados permitem que não-especialistas com habilidades básicas de laboratório (tais como estudantes de graduação) para implementar o crescimento celular, extrair a preparação e instalação de reação de formato de lote para uma Escherichia coli-CFPS sistema baseado. Esta abordagem é rentável em comparação aos kits comercialmente disponíveis sem sacrificar a facilidade de instalação baseado no kit de reação. Além disso, essa abordagem permite que os aplicativos no laboratório e no campo. Ao decidir implementar CFPS, novos usuários devem avaliar cuidadosamente a eficácia dos sistemas de expressão da proteína convencional para investimento de inicialização, como CFPS não pode ser superior em todos os casos. Os métodos CFPS descritos aqui permitem ao usuário diretamente implementar uma variedade de aplicações, incluindo genômica funcional, alta produtividade, a produção de proteínas que são intratáveis por expressão em vivo , bem como campo de teste aplicações, incluindo biosensores e kits educativos para biologia sintética. Aplicações adicionais, tais como a engenharia metabólica, ajuste das condições de expressão de proteínas, doença detecção e aumentar usando métodos CECF ou CFCF ainda são possíveis, mas podem exigir a experiência com a plataforma CFPS de modificações adicionais de reação condições. Nossos métodos combinam crescimento em meios enriquecidos e frascos perplexos, com métodos relativamente rápidos e reprodutíveis de lise celular através de sonication, seguido de uma configuração simplificada de reação CFPS que utiliza premixes otimizado45. Enquanto os métodos de crescimento celular tem tornar-se um pouco padronizados dentro deste campo, métodos para lise celular variam amplamente. Além de sonication, métodos comuns de Lise incluem a utilização de uma imprensa francesa, um homogenizador, batedores do grânulo, ou lisozima e outras perturbações físicas e bioquímicas métodos46,,47,48, 49. usando nossos métodos, cerca de 2 mL de extrato bruto de celular são obtidos por 1 L de células. Esta quantidade de extrato celular pode oferecer suporte a quatrocentos 15 reações de CFPS µ l, cada produção ~ 900 µ g/mL de proteína de sfGFP repórter do modelo do plasmídeo pJL1-sfGFP. Esse método custa $0,021 / µ g de sfGFP produzido ($.019 / µ l de reação), excluindo o custo de mão de obra e equipamentos (Supplemental Figura 1). A partir do zero, este método pode ser implementado em 4 dias por uma única pessoa e repita as reações CFPS podem ser concluídas dentro de horas (Figura 1). Além disso, o protocolo pode ser escalado em volume para lotes maiores de preparação dos reagentes para atender às necessidades do usuário. Importante, o protocolo aqui apresentado pode ser implementado pelo laboratório treinado não-especialistas como alunos de graduação, pois só requer habilidades básicas de laboratório. Os procedimentos descritos abaixo e o de acompanhamento foram especificamente desenvolvidos para melhorar a acessibilidade da plataforma CFPS Escherichia coli para uso amplo.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. meios preparação e crescimento celular
-
Dia 1
- Células de Escherichia coli BL21*(DE3) raia de glicerol um estoque em uma placa de ágar LB e incubam durante pelo menos 18 h a 37 ° C.
- Prepara-se 50 mL de LB mídia e autoclave a solução em um ciclo de líquido por 30 min a 121 ° C. Armazenar em temperatura ambiente.
-
Dia 2
- Prepare 750 mL de 2 x YTP mídia e 250 mL de solução de 0,4 M D-glicose como descrita na informação suplementar.
- Despeje a mídia x YTP 2 em um balão de perplexo autoclavado 2,5 L e a solução de D-glicose em um frasco de vidro 500ml autoclavados. Autoclave ambas as soluções em um ciclo de líquido por 30 min a 121 ° C.
- Certifique-se de que ambas as soluções estéreis são armazenadas a 37 ° C, se o crescimento celular está sendo executado no dia seguinte, para maximizar as taxas de crescimento após a inoculação. Não combine soluções até a inoculação.
Nota: Soluções podem ser armazenadas a 4 ° C, durante 1-2 d, se necessário, embora a mídia x YTP 2 é altamente propenso à contaminação. - Começa uma cultura durante a noite de BL21(DE3) por inocular 50 mL de LB mídia com uma única colônia de BL21(DE3) usando um loop esterilizada e técnica asséptica para evitar contaminação.
- Coloque a 50 mL de cultura BL21*(DE3) LB a 37 ° C 250 rpm tremendo incubadora e crescer durante a noite por 15-18 h.
- Preparar e esterilizar todos os materiais necessários para os dias 3 e 4, incluindo: duas garrafas de centrífuga de 1 L, 4 x frio cónico tubos de 50 mL (pesagem e registros massas de três) e muitos tubos de microcentrífuga de 1,5 mL.
-
Dia 3
- Remova a cultura da noite 50 mL de BL21*(DE3) de LB a incubadora tremendo e medir o OD600 em um espectrofotômetro usando um 01:10 diluição com LB mídia. Calcular o volume da noite cultura necessário adicionar a 1 L de mídia para uma partida de OD600 de 0.1 (por exemplo, se uma OD600 de um 01:10 diluição é lido como 0.4, inocular 25 mL de OD não diluído600 = 4.0 cultura durante a noite para 1 L de 2 x YTP G).
- Remova aquecido 2 x YTP meios de comunicação e soluções de D-Glucose a incubadora 37 ° C, juntamente com os 50 mL de cultura LB. Utilizando uma técnica estéril, com cuidado, despeje a solução de D-Glucose na mídia x YTP 2 (evitando os lados do balão perplexo).
Nota: A adição de D-Glucose completa a receita para 1 L de 2 x YTPG. - Mantendo uma técnica estéril, inocule a 1L de solução 2 de x YTPG com a quantidade adequada de cultura 50ml para iniciar a cultura de 1L em um 0.1 OD600. Coloca imediatamente a cultura 1L inoculados em um 37 ° C, agitando a incubadora a 200 rpm.
- Pegue o primeiro OD600 leitura após a primeira hora de crescimento (fase de retardo típico leva 1 h). Não dilua a cultura. Continue a tomar medidas de OD600 aproximadamente cada 20-30 min até OD600 atinge 0,6.
- Ao atingir OD600 = 0,6, adicionar 1 mL de 1 M IPTG (concentração final na cultura 1 L = 1 mM) para a cultura de x YTPG 2.
Nota: Indução Ideal OD600 é 0,6; no entanto, uma gama de 0.6-0.8 é aceitável. Indução por IPTG é para a produção endógena de T7 RNA polimerase (T7RNAP). - Após a indução, medir o OD600 aproximadamente a cada 20-30 minutos até que ele atinja o 3.0.
Nota: Refrigerar para baixo o centrifugador a 4 ° C durante este tempo. Prepare o buffer a frio S30 conforme detalhado nas informações complementares. Se o buffer de S30 é preparado com antecedência, certifique-se de que a TDT não é adicionada até o dia de uso. - Uma vez que o OD600 atinge 3.0 (Figura 2A), despeje a cultura em uma garrafa de centrífuga 1L frio num banho de água gelada. Prepare uma garrafa cheio de água centrífuga de 1 L de igual peso para ser usado como um equilíbrio na centrífuga.
Nota: Os valores de absorvância variam de instrumento-à-instrumento. Enquanto o OD600 da colheita de BL21(DE3) não é uma variável sensível, é recomendável que o usuário avaliar e otimizar essa variável como uma medida de resolução de problemas. Espectrofotômetros maiores podem resultar em leituras de600 OD relativamente baixas em comparação com menores baseados na cubeta espectrofotômetros. - Centrifugar as garrafas de 1 L para 10 min a 5.000 x g e 10 ° C a células de Pelotas.
- Lentamente, decantar o sobrenadante e elimine-os em conformidade com procedimentos de resíduos biológicos da instituição. Coloque a pelota no gelo.
- Usando uma espátula estéril, raspar o centrifugado da garrafa centrífuga e transferi-lo para um tubo cônico de frio 50 mL.
- Adicionar 30 mL de tampão de S30 frio ao tubo cónico e ressuspender as células vortexing com rajadas curtas (20-30 s) e períodos de repouso (1 min) no gelo até totalmente resuspended com nenhum pedaços.
- Uma vez que o sedimento é totalmente resuspended, use outro tubo cónico de 50 mL com água como um equilíbrio e centrifugar por 10 min em 5000 x g e 10 ° C (pre-resfriado a 4 ° C).
Nota: Esta completa 1st de 3 lavagens necessárias ao colher as células. - Despeje o líquido sobrenadante e elimine-os em conformidade com procedimentos de resíduos biológicos da instituição. Resuspenda o pellet com 20-25 mL de frio S30 buffer e centrifugação por 10min a 5000 x g e 10 ° C (pre-resfriado a 4 ° C).
Nota: Isto completa a 2nd de 3 lavagens. - Novamente, despeje o líquido sobrenadante e elimine-os em conformidade com procedimentos de resíduos biológicos da instituição. Adicione exatamente a 30 mL de tampão S30 e vórtice novamente para Ressuspender o precipitado.
- Usando os 3 tubos cónicos 50ml pré-pesados, frio e um enchedor de pipeta sorológica com uma pipeta esterilizada, alíquota 10 mL da mistura de tampão de ressuspensão de sedimento/S30 em cada um dos 3 tubos cónicos.
Nota: Dividir as células em 3 tubos não é necessária, mas esta etapa resulta em menor célula Pelotas (~ 1 g) para maior conveniência em etapas posteriores. - Todos os tubos de centrifuga, usando apropriado saldos conforme necessário, para 10 min a 5000 x g e 10 ° C (pre-resfriado a 4 ° C).
Nota: Isto é concluída a etapa de lavagem final. - Despeje o líquido sobrenadante e elimine-os em conformidade com procedimentos de resíduos biológicos da instituição. Remover o excesso S30 buffer limpando cuidadosamente o interior do tubo cônico e tampa com um tecido limpo; Evite tocar a pelota.
- Pesar de novo os tubos em uma balança analítica e anotar o peso final da pelota em cada tubo.
Nota: O protocolo pode ser interrompido neste ponto. Os pellets podem ser flash congelado em nitrogênio líquido e armazenado a-80 ° C por até um ano, até que seja necessário para a preparação do extrato.
2. preparação de extrato celular bruto - dia 4
- Para preparação do extrato, não deixar as células no gelo durante cada etapa. Adicione 1 mL de buffer a frio S30 1 g de massa celular da pelota. Certifique-se de que ditiotreitol (DTT) foi completado para o buffer de S30 para uma concentração final de 2 mM.
Nota: Refrigerar para baixo a microcentrifuga a 4 ° C durante este tempo. - Resuspenda o pellet celular vortexing com rajadas curtas (20-30 s) e períodos de repouso (1 min) no gelo até totalmente resuspended. Se ressuspensão é difícil, embora os pellets no gelo durante 30 minutos descongelar.
- Transferi 1,4 mL de ressuspensa células para um tubo de microcentrífuga de 1,5 mL.
- Coloque um tubo de 1,5 mL contendo 1,4 mL de ressuspensa células em um banho de água gelada num copo. Proceda à sonicação durante 45 s em seguida por 59 s fora por 3 ciclos totais, com amplitude fixada em 50%. Feche e inverter os tubos a misture suavemente durante os períodos de folga. No total, entrega 800-900 J de energia para cada tubo de microcentrífuga de 1,5 mL contendo 1,4 mL de ressuspensa células (Figura 3A e 3B).
Nota: Este passo é sensível ao tipo de sonicador e modelo usado e deve ser otimizado se o equipamento é diferente do listado para este procedimento. Duas abordagens complementares podem ser usadas para aumentar a quantidade de extrato preparado durante esta etapa: 1) vários tubos de microcentrífuga de 1,5 mL podem ser sonicated em paralelo e/ou volumes 2) maiores podem ser sonicated em tubos cónicos (até 15 mL de ressuspensão de célula por tubo) , dimensionar a quantidade de energia entregada conforme descrito anteriormente 29,,45. - Imediatamente depois da sonication, adicionar 4,5 µ l de TDT (completando um adicional 2 mM DTT) de 1 M a 1,4 ml de lisado e inverter várias vezes para misturar. Coloca o tubo no gelo. Repita as etapas de 2.4 e 2.5 para tubos de adicionais de ressuspensão células antes de proceder a centrifugação.
- Microcentrifuga amostras em 18.000 x g e 4 ° C por 10 min (Figura 3).
- Pipete o sobrenadante para um novo tubo de microcentrífuga de 1,5 mL. Não perturbe a pelota; é preferível deixar alguns sobrenadante para trás para manter a pureza do que to perturbar a pelota em esforços para maximizar o rendimento.
- Incube o sobrenadante da etapa anterior a 250 rpm e 37 ° C por 60 min gravando os tubos para a plataforma de agitação de incubadora (esta é a reação de escoamento).
- Microcentrifuga amostras em 10.000 x g e 4 ° C por 10 min.
- Remover o sobrenadante sem perturbar o sedimento e transferi-lo para um novo tubo. Crie muitos 100 alíquotas de µ l do extrato para o armazenamento.
Nota: O protocolo pode ser pausado aqui, e o extrato pode ser flash congelado em nitrogênio líquido e armazenado a-80 ° C por até um ano, até que seja necessário para reações de CFPS. Pelo menos 5 ciclos de gelo-degelo pode ser submetido sem detrimento para extrair produtividade (Figura 4).
3. célula livre proteína reações da síntese lote formato
- Degelo soluções A e B, DNA modelo, BL21(DE3) extrato (se congelado), T7RNAP e uma alíquota de água molecular da classe.
Nota: Modelo de reação CFPS pode ser encontrado nas Informações complementares. Receitas de soluções A e B são fornecidas nas Informações complementares e correspondem às concentrações específicas para inúmeros reagentes apoiar o sistema de energia de PANOx-SP com base para CFPS. O papel de cada reagente e a variação aceitável nestas concentrações de reagente que possa suportar CFPS ter sido determinado50. Um protocolo de purificação de T7RNAP pode ser encontrado na informações complementares51. T7RNAP suplementar pode aumentar o rendimento volumétrico, mas não é necessário se T7RNAP é induzida durante o crescimento celular. Modelo de DNA de plasmídeo (pJL1-sfGFP) pode ser preparado usando um kit de maxiprep com duas lavagens usando o tampão de lavagem no kit, seguido de uma limpeza de pós-processamento de DNA usando um kit de purificação de PCR (Figura 2B). Modelos de DNA lineares também podem ser usados em reações de CFPS. - Rotule a quantidade necessária de tubos de microcentrífuga necessário para reações de CFPS.
Nota: Reações podem ser executadas em vários tamanhos de navio, mas uma nave menor pode diminuir o rendimento volumétrico da proteína (Figura 2). Ampliação de uma reação no mesmo vaso tamanho também pode reduzir o rendimento volumétrico, em função de diminuir a troca de oxigênio, devido a uma diminuição da área de superfície à relação do volume. Quando aumentar o volume de reação acima de 100 μL, recomenda-se usar o fundo plano bem placas 31,37,52. - Adicionar 2,2 µ l da solução A, extrato de 2,1 µ l da solução B, 5 µ l de BL21*(DE3), 0,24 μg de T7RNAP (concentração final μg/mL 16), 0,24 ng de DNA modelo (concentração final de 16 ng/mL) e água para trazer o volume final de 15 µ l.
Nota: Vortex soluções A e B com frequência durante a configuração de reação para evitar a sedimentação de componentes e garantir que cada reação recebe uma homogênea aliquota de cada solução. Evitar a utilização do Vortex do extracto, em vez disso, inverta o tubo para misturar. - Depois que todos os reagentes foram adicionados para a reação, misture cada tubo pipetagem para cima e para baixo ou suavemente num Vortex, garantindo simultaneamente que a mistura de reação final é combinada em um único talão de 15 µ l no fundo do tubo de microcentrífuga de 1,5 mL.
- Coloque cada reação em incubadora a 37 ° C sem tremer por 4h, ou 30 ° C durante a noite.
Nota: Reações bem sucedidas podem ser qualitativamente avaliadas visualmente com base na cor verde do produto sfGFP dentro a mistura de reação CFPS (Figura 3D). Expressão da proteína de interesse também pode ser confirmada por SDS-PAGE (Supplemental Figura 2).
4. quantificação da proteína repórter, [sfGFP]
- Carrega 48 µ l de 0,05 M HEPES, pH 8, para cada um bem necessário para quantificação (geralmente realizada em triplicata por tubo de reação).
- Retire as reações da incubadora. Pipetar para cima e para baixo para misturar cada reação e, em seguida, transferir 2 µ l de reação para o 48 µ l de 0,05 M HEPES, pH 8. Pipetar acima e para baixo outra vez no poço para misturar.
- Uma vez que todas as reações são carregadas e misturadas, coloque a placa bem 96 para os dados e medir a fluorescência de ponto de extremidade sfGFP.
Nota: Comprimentos de onda de excitação e emissão para quantificação de fluorescência sfGFP são 485 nm e 510 nm, respectivamente. - Utilizando uma curva padrão gerada anteriormente, determine a [sfGFP] desde as leituras de fluorescência obtidos.
Nota: As instruções para gerar uma curva padrão de concentração de sfGFP contra a intensidade da fluorescência constam informações suplementares (complementar a Figura 3). Os usuários precisarão estabelecer uma curva padrão para seu instrumento, desde que a sensibilidade do instrumento pode variar.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Apresentamos uma base sonication Escherichia coli extrato de protocolo de preparação que pode ser concluído em um período de quatro dias, com a Figura 1 , demonstrando o esgotamento processual ao longo de cada dia. Há maleabilidade para as etapas que podem ser concluídas em cada dia com vários pontos de pausando, mas encontrámos este fluxo de trabalho para ser o mais eficaz executar. Além disso, ambas as pelotas de célula (etapa 1.3.18) e totalmente preparado extrato (passo 2.10) são estáveis a-80 ° C durante pelo menos um ano, permitindo ao usuário criar maiores estoques de cada um para salvar para uso em uma posterior tempo17. Não só foi o extrato estável durante longos períodos de tempo, mas o extrato também pode sofrer ciclos de degelo congelar pelo menos cinco sem uma perda significativa de produtividade (Figura 4). Isto permite maiores alíquotas do extrato para ser armazenado para usos múltiplos, se o espaço de armazenamento do congelador é limitado. No entanto, recomendamos várias alíquotas menores (~ 100 μL) de extrato, sempre que possível.
Com cada nova preparação do extrato, recomendamos que o usuário executa uma titulação do magnésio a fim de determinar a quantidade ideal de magnésio para aquele lote de extrato. Os usuários podem quantificar lotes a variabilidade na concentração de proteínas totais do extrato celular pelo ensaio de Bradford. Para maior desempenho extratos, vemos normalmente as concentrações de proteína total de 30-50 mg/mL, e dentro desta escala, não há nenhuma correlação óbvia entre as concentrações de proteína total e desempenho de extrato celular. Portanto, recomendamos que os usuários sintonizar as concentrações de magnésio em conformidade para garantir que a funcionalidade de ácido nucleico e proteína são maximizados para cada lote de extrato. Os níveis de magnésio são importantes para a adequada replicação do DNA, transcrição e tradução, mas os níveis excessivos podem ser prejudiciais a estes processos53. Para demonstrar essa dependência, podemos ter realizado uma titulação co de magnésio e extrair o volume para determinar a melhor combinação que minimiza a quantidade de extrato necessária, mantendo uma reação produtiva (Figura 5). Neste experimento, recomendamos 5 µ l de extracto e 10mm Mg2 + de extrato com um teor de proteínas totais de 30 mg/mL, a fim de obter mais 1.000 μg/mL de sfGFP.
Nossa experiência com CFPS também nos permitiu determinar etapas dentro do protocolo que pode ser variado sem prejuízo para a produtividade global do sistema e outros que são parte integrantes de um sistema CFPS alto desempenho. Mais notavelmente, o OD final600 de colheita da pilha não afeta significativamente o resultado final da reação CFPS, e células viável podem ser colhidas em qualquer lugar de 2.7-4.0 OD600. Isto representa a primeira fase exponencial de crescimento onde Ribossoma concentração por célula é a mais alta e a maquinaria de translação é o mais ativo para apoiar o crescimento rápido. Esta observação permite aos usuários a flexibilidade otimizar seus próprios procedimentos. Recomendamos que a colheita em aproximadamente 3.0 OD600 a fim de capturar as células em um OD600 mais perto de 3.3 pelo tempo da colheita é completa (Figura 2A). Variáveis que impactam rendimentos CFPS incluem modelo DNA de qualidade, tamanho de vaso de reação e as quantidades relativas de celular extrato e magnésio íon presente na reação. Encontramos qualidade DNA ter notável variação de lotes. Para resolver esse problema, recomendamos que os usuários purificam DNA através de uma preparação midi ou maxi, seguida por uma etapa de limpeza adicional do DNA ou na coluna de purificação de DNA usado no maxiprep, ou pós-purificação usando um kit de limpeza de DNA adicional. Isso melhora a reprodutibilidade na qualidade do DNA para reações CFPS e resulta na produção de proteína mais robusta (Figura 2B). A embarcação da reação também impacta o rendimento volumétrico, tal que a produção de proteína de configurações de reação idêntica em diferentes volumes de navio pode diferir até 40%. Tem sido teorizado que o aumento no rendimento volumétrico observado em vasos maiores é devido a um aumento da área superficial da mistura de reação, permitindo a troca de oxigênio melhor (Figura 2), e outros mais têm impulsionado o rendimento volumétrico, executando Reações de CFPS em grandes placas de fundo plano, que recomendamos para reações com 100 µ l17,31,37,52.

Figura 1 : Linha do tempo para o crescimento da cultura, produção de extrato celular, configuração e quantificação de reações CFPS. O usuário pode implementar a plataforma CFPS para suas aplicações de pesquisa através deste fluxo de trabalho de quatro dias. Preparação do reagente representa o principal tempo e custo de investimento para a primeira rodada desta experiência e diminui substancialmente depois estoques de reagentes forem estabelecidas. Adicionalmente, célula pelotas e extrato celular preparado podem ser armazenados por mais de um ano a-80 ° C, permitindo que o usuário começar a linha do tempo em várias etapas para resultados mais rápidos. O usuário também pode pausar em várias etapas para modificar o cronograma de fluxo de trabalho. Clique aqui para ver uma versão maior desta figura.

Figura 2 : Condições modificáveis para CFPS e os efeitos sobre o rendimento volumétrico reação. R. extrair comparação de produtividade com base na colheita de células BL21(DE3) em várias leituras de600 OD. Baseado nesta trama, recomendamos colheita em um OD de600 de 3.3 para produzir pelo menos 1000 µ g/mL de proteína do alvo. As reações foram realizadas em escala 15 μL em tubos de microcentrífuga de 1,5 mL. B. comparação de dois DNA maxiprep lavagem protocolos de com e sem pós-purificação DNA-limpezas. pJL1-sfGFP plasmídeos submeteu-se a um maxiprep com uma ou duas lavagens, seguidas de uma limpeza pós-purificação pelo kit de purificação de PCR. Para conseguir ~ 900 µ g/mL de expressão da proteína, sugerimos realizar uma limpeza de DNA de pós-purificação independentemente do número de lavagens de maxiprep. As reações foram realizadas em escala 15 μL em tubos de microcentrífuga de 1,5 mL. C. reações de CFPS 15 µ l executada em várias embarcações que variam de 2 mL a tubos de microcentrífuga de 0,6 mL. "Neg" representa um controlo negativo onde nenhum modelo de DNA foi adicionado para a reação. Todas as barras de erro representam 1 desvio-padrão de três reações independentes para cada condição, cada um dos quais foi quantificado em triplicado. Clique aqui para ver uma versão maior desta figura.

Figura 3 : Extraem o chave processuais configurações e resultados para a criação produtiva. R. adequada configuração do banho de água gelada sonication para garantir o resfriamento da amostra, enquanto o calor é gerado durante sonication. B. tubo de microcentrífuga de 1,5 mL contendo ressuspensão celular pelota pre (à esquerda) e sonication post (à direita). O lisado resultante deve exibir uma tonalidade mais escura em comparação a ressuspensão centrifugado. C. adequada separação do sobrenadante e sedimento de lisado depois de 18.000 x centrifugação g celular. M. reações de CFPS após 4 h de incubação a 37 ° C. tubo de microcentrífuga de 1,5 mL no lado direito (reação bem sucedida) mostra fluorescência visível da proteína repórter sfGFP ~ 900 μg/ml. O tubo controle negativo à esquerda, falta modelo DNA e simulando uma reação malsucedida, exibe uma solução clara com nenhuma fluorescência. Clique aqui para ver uma versão maior desta figura.

Figura 4 : Mudança na expressão de proteínas por 5 ciclos de congelamento e descongelamento de CFPS extrair. Extrato preparado a partir do crescimento mesmo passou por cinco congelar ciclos de descongelação através de nitrogênio líquido flash congelamento seguido de descongelamento no gelo. Nenhuma mudança significativa na produtividade de extrato para expressar sfGFP foram vista sobre os cinco ciclos de congelamento e descongelamento. As reações foram realizadas em escala 15 μL em tubos de microcentrífuga de 1,5 mL. "Neg" representa um controlo negativo onde nenhum modelo de DNA foi adicionado para a reação. Todas as barras de erro representam 1 desvio-padrão de três reações independentes para cada condição, cada um dos quais foi quantificado em triplicado. Clique aqui para ver uma versão maior desta figura.

Figura 5 : CFPS para reações com diferentes [Mg2+] e extrair volumes contra [sfGFP]. [Mg2 +] variou de 8 mM a 14 mM com incrementos de 2 mM e extrair volumes variou de 3 µ l a 7 µ l com incrementos de 1 µ l. A cor código representa [sfGFP] produzida a partir de alta (vermelho) para baixo (roxo). Para maximizar a eficiência de reagente, mantendo a produção de proteína de alta, recomendamos a utilização de 5 µ l de extracto e 10mm Mg2+ de extratos que tem um teor de proteína total de ~ 30 mg/mL, conforme determinado pelo ensaio de Bradford. Pontos originais para gerar o contorno enredo basearam-se fora da fluorescência de ponto de extremidade de três reações independentes para cada condição, cada um dos quais foi medido em triplicado. As reações foram realizadas em escala 15 μL em tubos de microcentrífuga de 1,5 mL. Clique aqui para ver uma versão maior desta figura.
Complementar a Figura 1: custo por micrograma de proteína produzida e por microlitro de reação através de plataformas de síntese de proteína sem célula seis. Nossa plataforma é comparada com cinco diferentes células proteína livre síntese kits/plataformas com variando de produtividade e de preços. Nossa plataforma CFPS sonication baseado é mais rentável em ambos $/ µ g de proteína e $/ µ l de reação do que a maioria dos comerciais kits de e fornece a facilidade de um kit para instalação de reação, enquanto o restante custo comparável a outras plataformas CFPS acadêmicas. Clique aqui para ver uma versão maior desta figura.
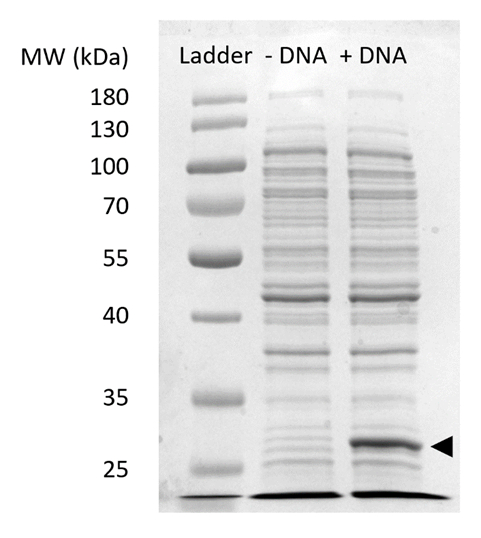
Complementar Figura 2: SDS-PAGE de expressão sfGFP no CFPS. Sem célula reações de síntese de proteína com (+ DNA) e sem (-DNA) modelo de DNA para sfGFP foram executados em um 12% gel de SDSPAGE para demonstrar a expressão de sfGFP observado em 27 kDa (seta preta). Foram utilizadas técnicas tradicionais de SDS-PAGE. Cada amostra carregados o gel extrato incluído µ 18 g de proteína total, com base na quantificação de ensaio de Bradford da proteína total na célula. Com base em medições de intensidade de fluorescência e nossa curva padrão, estimamos que o "+ DNA" pista contém 0,42 µ g de sfGFP. Para obter estas amostras, reações CFPS foram executadas em uma escala de 15 µ l em tubos de microcentrífuga de 1,5 mL produzindo rendimento volumétrico consistente com a Figura 3. Clique aqui para ver uma versão maior desta figura.
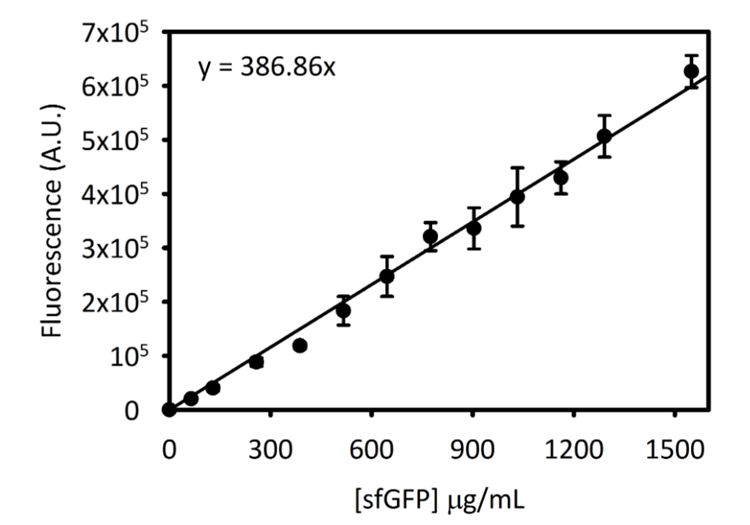
Complementar Figura 3: curva padrão para sfGFP na Cytation 5. Esta curva foi determinada utilizando os métodos descritos acima. Todos os dados coletados para este manuscrito foi convertido a partir de leituras de fluorescência de ponto de extremidade para [sfGFP] em µ g/mL, usando esta curva padrão. Clique aqui para ver uma versão maior desta figura.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Síntese de proteína sem célula tem emergido como uma tecnologia poderosa e propício para uma variedade de aplicações que vão desde biomanufacturing para prototipagem rápida de sistemas bioquímicos. A gama de aplicativos é compatível com a capacidade de monitorar, manipular e aumentar a maquinaria celular em tempo real. Apesar do impacto em expansão desta tecnologia de plataforma, manteve-se ampla adaptação lento devido a nuances técnicas na implementação dos métodos. Através desse esforço, pretendemos proporcionar simplicidade e clareza para o estabelecimento desta tecnologia em novos laboratórios. Para este fim, nosso protocolo para uma Escherichia coli-plataforma de síntese de proteína sem célula com base pode ser alcançado dentro de um tempo de inicialização de quatro dias pelo laboratório treinado não-especialistas, tais como estudantes de graduação (Figura 1). Além disso, uma vez que um estoque de reagentes e extrato são produzidos, subsequentes CFPS lote reações podem ser configurar, incubadas e quantificadas em apenas 5 h. Um único, o crescimento de células 1L pode resultar em suficiente extrato para reações de CFPS quatrocentos 15 µ l, enquanto preparações único lote dos outros reagentes sem célula podem suportar milhares de reações. Preparações de reagente também podem ser escaladas, se um estoque ainda maior é necessária. As reações de CFPS podem ser configurado de forma elevado-throughput, usando uma placa de 96 poços ou tubos PCR para teste de uma variedade de condições em paralelo. Rendimento volumétrico diminuirá quando usando embarcações menores, como visto na Figura 2. Reações CFPS podem também ser ampliadas de microlitros de dezenas de mililitros de volume total reação a fim de aumentar o rendimento de proteína total para uma única condição. Ao dimensionar o volume, a consideração principal é que o rendimento volumétrico de reação com a diminuição o rácio superfície de área e o volume da reação37,52. A fim de aumentar, mantendo o rendimento volumétrico semelhante da expressão da proteína, usuários devem dividir o volume de reação em numerosos vasos de reação e/ou aumentar o tamanho do navio. Para escalas de reação, variando de 15 µ l - 100 µ l de volume, numerosos 15 reações de µ l em paralelo são recomendadas. Para reações superior a 100 µ l de volume, recomendam-se placas de 24 poços de fundo plano e placas 12 boas são recomendadas para volumes de reação superior a 600 µ l. Tais pares de volumes de reação e navios fornecem consistência do rendimento volumétrico reação após aumentar17,31,37,,52. Ampliação do além desses volumes pode ser realizado utilizando múltiplas cavidades da placa em paralelo. Usando este formato, a reação pode ser escalada para o volume total de mais de 10 mL. Otimizando a combinação de navio de volume-reação de reação pode oferecer suporte a aplicativos de biomanufacturing sem sacrificar a produtividade da reação.
Ao realizar este protocolo, existem algumas considerações-chave que afetam o rendimento volumétrico de reação, bem como indicadores associados mal realizando extrato. Para garantir a adequada Lise e para evitar a desnaturação da maquinaria de transcrição/tradução funcional, é importante atenuar o calor produzido durante a Lise. Mergulhe a ressuspensão de células em um banho de água gelada durante sonication para dissipar o calor rapidamente durante sonication (Figura 3A). Um indicador de lise celular eficaz é o surgimento de uma aparência mais escura da célula lisada em comparação com as amostras pré-lisados (Figura 3B). Para flexibilidade do usuário, o sonicador e sonda mostrado na Figura 3A é adaptável a uma série de volumes de 100 µ l a 15 mL de células ressuspensa. Para fazer isso, o usuário pode ajustar o número de joules entregado para lise do volume desejado de células. Além disso, volumes maiores de extrato podem ser preparados através de duas abordagens complementares. Os usuários podem proceda à sonicação vários tubos em paralelo, e/ou proceda à sonicação volumes maiores de ressuspensão de célula, escala a quantidade de energia proporcionalmente com o volume como descrito anteriormente29,,45. Mais um passo que indica a qualidade do extrato é a etapa de centrifugação após a lise celular. Post lysis da pilha, nós recomendamos a centrifugação a 18.000 x g, para fornecer uma divisão clara entre o sobrenadante (maquinaria de transcrição/tradução, DNA genômico fragmentado que não funciona mais para modelo de transcrição e tradução) e o (pellet indesejada de componentes celulares como a membrana celular e precipitado de proteínas) (Figura 3). Encontramos essa centrifugação a 18.000 x g melhora a separação, resultando em melhor reprodutibilidade em comparação com rotações em velocidades mais baixas como a 12.000 x g. Para sua conveniência, nós recomendamos usar uma centrífuga refrigerada de mesa, capaz de atingir um mínimo de 12.000 x g. Esta etapa é também comumente realizada a 30.000 x g, que deve ser considerada se o equipamento apropriado está disponível54,55,56,57,58, 59 , 60. extrato de desempenho não é afetado pelas velocidades de centrifugação a este passo dado que separação adequada é alcançada. Ao remover o sobrenadante desejado, é melhor evitar qualquer nublados materiais que existem na fronteira entre o sobrenadante e sedimento já esta contaminação reduz a produtividade do extrato. Visando a pureza do líquido sobrenadante resulta em extractos mais produtivos e vale a pena a reduzida quantidade de extracto obtido para novos usuários.
É importante a nota que enquanto os métodos que apresentamos são reprodutíveis e podem ser executados por cientistas com experiência mínima, pode haver a lotes e variação de reação-para-reação. Isto pode ser atribuído a variação na composição proteomic do lisado pós-sonication61. A variabilidade de lotes que temos observado é geralmente diminuída após suplementação com T7RNAP e otimização das concentrações de magnésio. Adição exógena de T7RNAP é comum entre reações CFPS para apoiar a expressão da proteína ideal, e achamos que ter duas fontes de T7RNAP - expressão endógena em BL21*(DE3) e o T7RNAP suplementar a uma concentração final de 16 µ g/mL - melhora reprodutibilidade de lotes para novos usuários45,46. Com a experiência, os usuários podem modificar seus experimentos para utilizar apenas uma única fonte de T7RNAP, se desejado. Quantificação do teor de proteínas totais de um novo lote de extrato e ajustamento adequado da concentração de Mg2 + também pode ajudar a diminuir a variação de lotes em rendimentos de expressão de proteínas volumétrico. Variações na expressão de proteínas também podem ser devido a diferenças no tamanho e estrutura da proteína de interesse, o uso de códon do gene e sua correspondente sítio de ligação do ribossomo do gene de interesse, bem como o tipo de vetor de expressão usado62 ,,63. Por estas razões, algumas proteínas não podem expressar assim como o modelo sfGFP de proteína, resultando em reduzido rendimento volumétrico de reações CFPS.
Limitações da técnica CFPS apresentada incluem que pode não ser diretamente apropriado para todas as aplicações de célula-livre, tais como a engenharia metabólica e otimização das condições de expressão, sem modificações adicionais para os protocolos. No entanto, acreditamos que este protocolo irá fornecer uma base para o estabelecimento da plataforma CFPS em laboratórios novos e fornecer não especialistas com a capacidade de implementar reações sem célula introdutórias em seus laboratórios. Após a implementação inicial, os pesquisadores podem experimentar com a plataforma para fazer suas próprias modificações para aplicações mais específicas, com base em outra literatura no campo.
A proteína CFPS plataforma custos $0,021 / µ g (excluindo o custo de mão de obra e equipamentos), tornando nosso sistema a preços competitivos com kits comerciais sem comprometer a facilidade de instalação de reação. Avaliações de custos comparativos por µ l de reação mostram tendências similares (suplementar Figura 1). Estimamos que os custos de inicialização para ser ~ US $4.500 para todos os reagentes e um adicional de US $3.200 de equipamento especializado, como um sonicador. Horas de pessoa para concluir este procedimento são estimadas em ~ 26 h para a preparação do reagente todos da terra acima. No entanto, uma vez que grandes estoques de reagentes foram preparadas, demandas no trabalho diminuem substancialmente. Além disso, como a experiência com a plataforma é adquirida, nós recomendo dimensionamento até o tamanho do crescimento da célula, extrato de preparação e preparação dos reagentes para maximizar a eficiência de tempo. Tendo em conta os custos de arranque, recomendamos a plataforma CFPS para aplicações em biologia sintética, os esforços de alta produtividade, e condições de expressão de proteínas que são incompatíveis com plataformas de expressão tradicional da proteína devido a um conflito com a célula restrições de Bioquímica e viabilidade. Nestes casos especializados, onde a técnica desejada é ativada pela plataforma CFPS, justifica-se o maior custo de CFPS sobre expressão na vivo .
Desenvolvimento contínuo da plataforma CFPS é susceptível de fornecer a mais ampla utilidade para os esforços de biotecnologia, como a engenharia metabólica de vias enzimáticas, produção e caracterização de proteínas tradicionalmente intratáveis, fora do padrão de aminoácidos incorporação e expressão da proteína natural, fabricação de medicamento estratificada e expansão além do laboratório em sala de aula para educação de tronco64,,65,66. Estes esforços serão ainda mais apoiados pelos esforços em curso para a caracterização detalhada da plataforma CFPS. Um melhor entendimento da composição do extrato celular vai levar ao refinamento contínuo em direção a reação maior rendimento e flexibilidade nas condições de reação a61,67,68.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Os autores declaram que eles têm não concorrentes interesses financeiros ou outros conflitos de interesse.
Acknowledgments
Autores gostaria de reconhecer o Dr. Jennifer VanderKelen, Andrea Laubscher e Tony Turretto para suporte técnico, Wesley Kao, Layne Williams e Christopher Hight para debates úteis. Os autores reconhecem também apoio financeiro do Bill e fundo de Frost Linda, centro para aplicações em Chevron biotecnologia aplicada pesquisa Endowment Grant da biotecnologia, Cal Poly pesquisa, publicações e programa de concessão de actividades criativas (2017 RSCA), e a National Science Foundation (NSF-1708919). MZL reconhece a Califórnia estado Universidade pós-graduação Grant. MCJ reconhece o escritório da pesquisa da exército W911NF-16-1-0372, National Science Foundation concede MCB-1413563 e MCB-1716766, a força aérea pesquisa laboratório centro de excelência Grant FA8650-15-2-5518, a concessão de agência de redução de ameaça de defesa HDTRA1-15-10052/P00001, o David e Lucile Packard Foundation, o programa de professor erudito Camille Dreyfus, o departamento de energia BER Grant DE-SC0018249, o programa de ciência humana fronteiras (RGP0015/2017), a concessão ETOP DOE Joint Genome Institute, e o consórcio biomédica de Chicago com o apoio dos fundos Searle no Chicago comunidade Trust para apoio.
Materials
| Name | Company | Catalog Number | Comments |
| Luria Broth | ThermoFisher | 12795027 | |
| Tryptone | Fisher Bioreagents | 73049-73-7 | |
| Yeast Extract | Fisher Bioreagents | 1/2/8013 | |
| NaCl | Sigma-Aldrich | S3014-1KG | |
| Potassium Phosphate Dibasic | Sigma-Aldrich | 60353-250G | |
| Potassium Phosphate Monobasic | Sigma-Aldrich | P9791-500G | |
| D-Glucose | Sigma-Aldrich | G8270-1KG | |
| KOH | Sigma-Aldrich | P5958-500G | |
| IPTG | Sigma-Aldrich | I6758-1G | |
| Mg(OAc)2 | Sigma-Aldrich | M5661-250G | |
| K(OAc) | Sigma-Aldrich | P1190-1KG | |
| Tris(OAc) | Sigma-Aldrich | T6066-500G | |
| DTT | ThermoFisher | 15508013 | |
| tRNA | Sigma-Aldrich | 10109541001 | |
| Folinic Acid | Sigma-Aldrich | F7878-100MG | |
| NTPs | ThermoFisher | R0481 | |
| Oxalic Acid | Sigma-Aldrich | P0963-100G | |
| NAD | Sigma-Aldrich | N8535-15VL | |
| CoA | Sigma-Aldrich | C3144-25MG | |
| PEP | Sigma-Aldrich | 860077-250MG | |
| K(Glu) | Sigma-Aldrich | G1501-500G | |
| NH4(Glu) | MP Biomedicals | 02180595.1 | |
| Mg(Glu)2 | Sigma-Aldrich | 49605-250G | |
| Spermidine | Sigma-Aldrich | S0266-5G | |
| Putrescine | Sigma-Aldrich | D13208-25G | |
| HEPES | ThermoFisher | 11344041 | |
| Molecular Grade Water | Sigma-Aldrich | 7732-18-5 | |
| L-Aspartic Acid | Sigma-Aldrich | A7219-100G | |
| L-Valine | Sigma-Aldrich | V0500-25G | |
| L-Tryptophan | Sigma-Aldrich | T0254-25G | |
| L-Phenylalanine | Sigma-Aldrich | P2126-100G | |
| L-Isoleucine | Sigma-Aldrich | I2752-25G | |
| L-Leucine | Sigma-Aldrich | L8000-25G | |
| L-Cysteine | Sigma-Aldrich | C7352-25G | |
| L-Methionine | Sigma-Aldrich | M9625-25G | |
| L-Alanine | Sigma-Aldrich | A7627-100G | |
| L-Arginine | Sigma-Aldrich | A8094-25G | |
| L-Asparagine | Sigma-Aldrich | A0884-25G | |
| Glycine | Sigma-Aldrich | G7126-100G | |
| L-Glutamine | Sigma-Aldrich | G3126-250G | |
| L-Histadine | Sigma-Aldrich | H8000-25G | |
| L-Lysine | Sigma-Aldrich | L5501-25G | |
| L-Proline | Sigma-Aldrich | P0380-100G | |
| L-Serine | Sigma-Aldrich | S4500-100G | |
| L-Threonine | Sigma-Aldrich | T8625-25G | |
| L-Tyrosine | Sigma-Aldrich | T3754-100G | |
| Fisherbrand Premium Microcentrifuge Tubes: 2.0 mL | Fisher Scientific | 05-408-138 | |
| Fisherbrand Premium Microcentrifuge Tubes: 1.5 mL | Fisher Scientific | 05-408-129 | |
| Fisherbrand Premium Microcentrifuge Tubes: 0.6 mL | Fisher Scientific | 05-408-120 | |
| PureLink HiPure Plasmid Prep Kit | ThermoFisher | K210007 | |
| Ultrasonic Processor | QSonica | Q125-230V/50Hz | 3.175 mm diameter probe |
| Avanti J-E Centrifuge | Beckman Coulter | 369001 | |
| JLA-8.1000 Rotor | Beckman Coulter | 366754 | |
| 1L Centrifuge Tube | Beckman Coulter | A99028 | |
| Tunair 2.5L Baffeled Shake Flask | Sigma-Aldrich | Z710822 | |
| Microfuge 20 | Beckman Coulter | B30134 | |
| New Brunswick Innova 42/42R Incubator | Eppendorf | M1335-0000 | |
| Cytation 5 | BioTek | ||
| Strep-Tactin XT Starter Kit | IBA | 2-4998-000 | |
| pJL1-sfGFP | Addgene | 69496 | |
| BL21(DE3) | New England BioLabs |
References
- Jiang, L., Zhao, J., Lian, J., Xu, Z. Cell-free protein synthesis enabled rapid prototyping for metabolic engineering and synthetic biology. Synthetic and Systems Biotechnology. 3 (2), 90-96 (2018).
- Carlson, E. D., Gan, R., Hodgman, C. E., Jewett, M. C. Cell-free protein synthesis: Applications come of age. Biotechnology Advances. 30 (5), 1185-1194 (2012).
- Watanabe, M. Cell-Free Protein Synthesis for Structure Determination by X-ray Crystallography. Methods in molecular biology (Clifton, N.J). 607, 149-160 (2010).
- Martemyanov, K. A., Shirokov, V. A., Kurnasov, O. V., Gudkov, A. T., Spirin, A. S. Cell-Free Production of Biologically Active Polypeptides: Application to the Synthesis of Antibacterial Peptide Cecropin. Protein Expression and Purification. 21 (3), 456-461 (2001).
- Renesto, P., Raoult, D. From genes to proteins: in vitro expression of rickettsial proteins. Annals of the New York Academy of Sciences. 990, 642-652 (2003).
- Xu, Z., Chen, H., Yin, X., Xu, N., Cen, P. High-Level Expression of Soluble Human b-Defensin-2 Fused With Green Fluorescent Protein in Escherichia coli Cell-Free System. Applied Biochemistry and Biotechnology. 127 (1), 053-062 (2005).
- Baumann, A. In-situ observation of membrane protein folding during cell-free expression. PLoS ONE. 11 (3), 1-15 (2016).
- Wang, Y., Percival, Y. H. P. Cell-free protein synthesis energized by slowly-metabolized maltodextrin. BMC Biotechnology. 9, 1-8 (2009).
- Whittaker, J. W. Cell-free protein synthesis: the state of the art. Biotechnology Letters. 35 (2), 143-152 (2013).
- Martin, R. W. Cell-free protein synthesis from genomically recoded bacteria enables multisite incorporation of noncanonical amino acids. Nature Communications. 9 (1), 1203 (2018).
- Kwon, Y. C., Song, J. K., Kim, D. M. Cloning-Independent Expression and Screening of Enzymes Using Cell-Free Protein Synthesis Systems. Methods in Molecular Biology. 1118, 97-108 (2014).
- Chappell, J., Jensen, K., Freemont, P. S. Validation of an entirely in vitro approach for rapid prototyping of DNA regulatory elements for synthetic biology. Nucleic Acids Research. 41 (5), 3471-3481 (2013).
- Takahashi, M. K. Characterizing and prototyping genetic networks with cell-free transcription-translation reactions. Methods. 86, 60-72 (2015).
- Karim, A. S., Jewett, M. C. A cell-free framework for rapid biosynthetic pathway prototyping and enzyme discovery. Metabolic Engineering. 36, 116-126 (2016).
- Dudley, Q. M., Anderson, K. C., Jewett, M. C. Cell-Free Mixing of Escherichia coli Crude Extracts to Prototype and Rationally Engineer High-Titer Mevalonate Synthesis. ACS Synthetic Biology. 5 (12), 1578-1588 (2016).
- Pardee, K. Paper-based synthetic gene networks. Cell. 159 (4), 940-954 (2014).
- Zawada, J. F. Microscale to manufacturing scale-up of cell-free cytokine production-a new approach for shortening protein production development timelines. Biotechnology and Bioengineering. 108 (7), 1570-1578 (2011).
- Sullivan, C. J. A cell-free expression and purification process for rapid production of protein biologics. Biotechnology Journal. 11 (2), 238-248 (2016).
- Li, J. Cell-free protein synthesis enables high yielding synthesis of an active multicopper oxidase. Biotechnology Journal. 11 (2), 212-218 (2016).
- Heinzelman, P., Schoborg, J. A., Jewett, M. C. pH responsive granulocyte colony-stimulating factor variants with implications for treating Alzheimer's disease and other central nervous system disorders. Protein Engineering Design and Selection. 28 (10), 481-489 (2015).
- Pardee, K. Low-Cost Detection of Zika Virus Using Programmable Biomolecular Components. Cell. 165 (5), 1255-1266 (2016).
- Slomovic, S., Pardee, K., Collins, J. J. Synthetic biology devices for in vitro and in vivo diagnostics. Proceedings of the National Academy of Sciences. 112 (47), 14429-14435 (2015).
- Gootenberg, J. S. Nucleic acid detection with CRISPR-Cas13a/C2c2. Science. 356 (6336), 438-442 (2017).
- Pardee, K. Portable, On-Demand Biomolecular Manufacturing. Cell. 167 (1), 248-259 (2016).
- Karig, D. K., Bessling, S., Thielen, P., Zhang, S., Wolfe, J. Preservation of protein expression systems at elevated temperatures for portable therapeutic production. Journal of the Royal Society, Interface. 14 (129), (2017).
- Smith, M. T., Berkheimer, S. D., Werner, C. J., Bundy, B. C. Lyophilized Escherichia coli-based cell-free systems for robust, high-density, long-term storage. BioTechniques. 56 (4), 186-193 (2014).
- Hunt, J. P., Yang, S. O., Wilding, K. M., Bundy, B. C. The growing impact of lyophilized cell-free protein expression systems. Bioengineered. 8 (4), 325-330 (2017).
- Stark, J. C. BioBitsTM Bright: A fluorescent synthetic biology education kit. Science Advances. 4 (8), (2018).
- Huang, A. BioBitsTM Explorer: A modular synthetic biology education kit. Science Advances. 4 (8), (2018).
- Zemella, A., Thoring, L., Hoffmeister, C., Kubick, S. Cell-Free Protein Synthesis: Pros and Cons of Prokaryotic and Eukaryotic Systems. ChemBioChem. 16 (17), 2420-2431 (2015).
- Oza, J. P. Robust production of recombinant phosphoproteins using cell-free protein synthesis. Nature Communications. 6 (1), 8168 (2015).
- Zemella, A. Cell-free protein synthesis as a novel tool for directed glycoengineering of active erythropoietin. Scientific Reports. 8 (1), 8514 (2018).
- Jaroentomeechai, T. Single-pot glycoprotein biosynthesis using a cell-free transcription-translation system enriched with glycosylation machinery. Nature Communications. 9 (1), 2686 (2018).
- Kightlinger, W. Design of glycosylation sites by rapid synthesis and analysis of glycosyltransferases. Nature Chemical Biology. 14 (6), 627-635 (2018).
- Schoborg, J. A. A cell-free platform for rapid synthesis and testing of active oligosaccharyltransferases. Biotechnology and Bioengineering. 115 (3), 739-750 (2018).
- Chekulayeva, M. N., Kurnasov, O. V., Shirokov, V. A., Spirin, A. S. Continuous-Exchange Cell-Free Protein-Synthesizing System: Synthesis of HIV-1 Antigen Nef. Biochemical and Biophysical Research Communications. 280 (3), 914-917 (2001).
- Hong, S. H. Improving Cell-Free Protein Synthesis through Genome Engineering of Escherichia coli Lacking Release Factor 1. ChemBioChem. 16 (5), 844-853 (2015).
- Endo, Y., Otsuzuki, S., Ito, K., Miura, K. Production of an enzymatic active protein using a continuous flow cell-free translation system. Journal of Biotechnology. 25 (3), 221-230 (1992).
- Volyanik, E. V., Dalley, A., Mckay, I. A., Leigh, I., Williams, N. S., Bustin, S. A. Synthesis of Preparative Amounts of Biologically Active Interleukin-6 Using a Continuous-Flow Cell-Free Translation System. Analytical Biochemistry. 214 (1), 289-294 (1993).
- Martin, G. A., Kawaguchi, R., Lam, Y., DeGiovanni, A., Fukushima, M., Mutter, W. High-yield, in vitro protein expression using a continuous-exchange, coupled transcription/ translation system. BioTechniques. 31 (4), 948-950 (2001).
- Stech, M., Quast, R. B., Sachse, R., Schulze, C., Wüstenhagen, D. A., Kubick, S. A Continuous-Exchange Cell-Free Protein Synthesis System Based on Extracts from Cultured Insect Cells. PLoS ONE. 9 (5), e96635 (2014).
- Quast, R. B., Sonnabend, A., Stech, M., Wüstenhagen, D. A., Kubick, S. High-yield cell-free synthesis of human EGFR by IRES-mediated protein translation in a continuous exchange cell-free reaction format. Scientific Reports. 6 (1), 30399 (2016).
- Thoring, L., Dondapati, S. K., Stech, M., Wüstenhagen, D. A., Kubick, S. High-yield production of "difficult-to-express" proteins in a continuous exchange cell-free system based on CHO cell lysates. Scientific Reports. 7 (1), 11710 (2017).
- Hoffmann, M., Nemetz, C., Madin, K., Buchberger, B. Rapid translation system: A novel cell-free way from gene to protein. Biotechnology annual review. 10, 1-30 (2004).
- Kwon, Y. C., Jewett, M. C. High-throughput preparation methods of crude extract for robust cell-free protein synthesis. Scientific Reports. 5 (1), 8663 (2015).
- Katsura, K. A reproducible and scalable procedure for preparing bacterial extracts for cell-free protein synthesis. Journal of Biochemistry. 162 (June), 357-369 (2017).
- Fujiwara, K., Doi, N. Biochemical preparation of cell extract for cell-free protein synthesis without physical disruption. PLoS ONE. 11 (4), 1-15 (2016).
- Shrestha, P., Holland, T. M., Bundy, B. C. Streamlined extract preparation for Escherichia coli-based cell-free protein synthesis by sonication or bead vortex mixing. BioTechniques. 53 (3), 163-174 (2012).
- Krinsky, N. A Simple and Rapid Method for Preparing a Cell-Free Bacterial Lysate for Protein Synthesis. PLoS ONE. 11 (10), (2016).
- Jewett, M. C., Swartz, J. R. Mimicking the Escherichia coli cytoplasmic environment activates long-lived and efficient cell-free protein synthesis. Biotechnology and Bioengineering. 86 (1), 19-26 (2004).
- Swartz, J. R., Jewett, M. C., Woodrow, K. A. Cell-Free Protein Synthesis With Prokaryotic Combined Transcription-Translation. Recombinant Gene Expression. (267), 169-182 (2004).
- Voloshin, A. M., Swartz, J. R. Efficient and scalable method for scaling up cell free protein synthesis in batch mode. Biotechnology and Bioengineering. 91 (4), 516-521 (2005).
- Vernon, W. B. The role of magnesium in nucleic-acid and protein metabolism. Magnesium. 7 (5-6), 234-248 (1988).
- Pratt, J. M. Transcription and Translation: A Practical Approach. , IRL Press. New York. (1984).
- Kim, D. M., Kigawa, T., Choi, C. Y., Yokoyama, S. A Highly Efficient Cell-Free Protein Synthesis System from Escherichia coli. European Journal of Biochemistry. 239 (3), 881-886 (1996).
- Shin, J., Noireaux, V. Efficient cell-free expression with the endogenous E. Coli RNA polymerase and sigma factor 70. Journal of Biological Engineering. 4 (1), 8 (2010).
- Zubay, G. In Vitro Synthesis of Protein in Microbial Systems. Annual Review of Genetics. 7 (1), 267-287 (1973).
- Kigawa, T. Preparation of Escherichia coli cell extract for highly productive cell-free protein expression. Journal of Structural and Functional Genomics. 5 (1/2), 63-68 (2004).
- Liu, D. V., Zawada, J. F., Swartz, J. R. Streamlining Escherichia Coli S30 Extract Preparation for Economical Cell-Free Protein Synthesis. Biotechnology Progress. 21 (2), 460-465 (2008).
- Yang, W. C., Patel, K. G., Wong, H. E., Swartz, J. R. Simplifying and streamlining Escherichia coli-based cell-free protein synthesis. Biotechnology Progress. 28 (2), 413-420 (2012).
- Foshag, D. The E. coli S30 lysate proteome: A prototype for cell-free protein production. New Biotechnology. 40 (Pt B), 245-260 (2018).
- Caschera, F. Bacterial cell-free expression technology to in vitro systems engineering and optimization. Synthetic and Systems Biotechnology. 2 (2), 97-104 (2017).
- Chizzolini, F., Forlin, M., Yeh Martín, N., Berloffa, G., Cecchi, D., Mansy, S. S. Cell-Free Translation Is More Variable than Transcription. ACS Synthetic Biology. 6 (4), 638-647 (2017).
- Hong, S. H., Kwon, Y. C., Jewett, M. C. Non-standard amino acid incorporation into proteins using Escherichia coli cell-free protein synthesis. Frontiers in Chemistry. 2, 34 (2014).
- Sawasaki, T., Ogasawara, T., Morishita, R., Endo, Y. A cell-free protein synthesis system for high-throughput proteomics. Proceedings of the National Academy of Sciences of the United States of America. 99 (23), 14652-14657 (2002).
- Dudley, Q. M., Karim, A. S., Jewett, M. C. Cell-free metabolic engineering: Biomanufacturing beyond the cell. Biotechnology Journal. 10 (1), 69-82 (2015).
- Garcia, D. C. Elucidating the potential of crude cell extracts for producing pyruvate from glucose. Synthetic Biology. 3 (1), 1-9 (2018).
- Hurst, G. B. Proteomics-Based Tools for Evaluation of Cell-Free Protein Synthesis. Analytical Chemistry. 89 (21), 11443-11451 (2017).

{kind=link}
{kind=link}
{kind=link}