

Summary
Dieses Protokoll beschreibt die Schritte, die Kosten und die notwendige Ausrüstung für E. Colizu generieren-basierte Zellextrakte und in-vitro- Protein Synthesereaktionen innerhalb von 4 Tagen oder weniger zu implementieren. Um die flexiblen Charakter dieser Plattform für breite Anwendungen nutzen zu können, diskutieren wir Reaktionsbedingungen, die angepasst und optimiert werden können.
Abstract
In den letzten 50 Jahren entstanden zellfreien Protein Synthese (GFP) als eine leistungsstarke Technologie, die transkriptionelle und translationale Kapazität von Zellen in einem Reagenzglas nutzbar zu machen. Durch die Notwendigkeit, die Lebensfähigkeit der Zelle vermieden und durch den Wegfall der zellulären Barriere wurde GFP grundlegend für neue Anwendungen in Biomanufacturing traditionell anspruchsvolle Proteine sowie Anwendungen im rapid Prototyping für Metabolic Engineering und funktionelle Genomik. Unsere Methoden für die Umsetzung einer E. Coli-basierten GFP-Plattform ermöglichen neue viele dieser Anwendungen Zugriff auf. Hier beschreiben wir Methoden um Extrakt durch angereicherte Medien, verblüfft Fläschchen und eine reproduzierbare Methode der einstellbaren Ultraschall-basierte Zelle Lysis vorzubereiten. Dieser Extrakt kann dann für Protein-Expression in der Lage 900 µg/mL oder mehr super Ordner grün fluoreszierenden Proteins (SfGFP) in nur 5 h vom Versuchsaufbau zur Datenanalyse, verwendet werden, angesichts der Tatsache, dass entsprechende Reagenz Bestände im Voraus vorbereitet worden sind. Voraussichtliche Inbetriebnahme kostet Reagenzien zu erhalten $4.500, die Tausende von Reaktionen mit geschätzten Kosten von $0,021 pro µg Protein produziert oder $0,019 pro Mikroliter Reaktion aufrechterhalten wird. Darüber hinaus spiegeln die Protein-Ausdruck-Methoden die Leichtigkeit der Reaktion Installation in marktübliche Systeme durch Optimierung des Reagenz Vormischungen, zu einem Bruchteil der Kosten gesehen. Damit den Benutzer die flexible Art der GFP-Plattform für breite Anwendungen nutzen kann, haben wir eine Vielzahl von Aspekten der Plattform identifiziert, die abgestimmt und je nach den verfügbaren Ressourcen und die Protein Ausdruck Ergebnisse optimiert werden können auf Wunsch.
Introduction
Zellfreien Protein Synthese (GFP) hat eine Technologie entwickelt, die eine Reihe von neuen Möglichkeiten für die Proteinproduktion, funktionelle Genomik, metabolische Technik und mehr innerhalb der letzten 50 Jahre1,2freigeschaltet hat. Im Vergleich zu standard in Vivo Protein Ausdruck Plattformen, GFP bietet drei entscheidende Vorteile: 1) die zellfreie Natur der Plattform ermöglicht die Produktion von Proteinen, die potenziell toxischen oder die Zelle3,4 fremd wäre ,5,6; (2) Inaktivierung von genomischer DNA und die Einführung einer Vorlage DNA Kodierung der DNA des Interesses Kanal aller die systemische Energie in die Reaktion auf die Produktion von der Aktivit‰tderzur von Interesse; und 3) der offene Charakter der Plattform ermöglicht dem Benutzer, ändern und überwachen die Reaktionsbedingungen und Zusammensetzung in Echtzeit7,8. Dieser direkte Zugang zur Reaktion unterstützt die Vermehrung von biologischen Systemen mit erweiterten Chemikalien und Redox-Bedingungen für die Produktion von neuartigen Proteinen und der Abstimmung des metabolischen Prozesse2,9, 10. direkter Zugriff erlaubt auch dem Benutzer, die GFP-Reaktion mit Aktivität Assays in einem Single-Topf-System zu verbinden, für mehr Design-Build-Schnelltest11Zyklen. Die Kapazität die GFP-Reaktion in kleinen Tröpfchen oder auf Papier-basierten Geräten weiter unterstützt Hochdurchsatz-Entdeckung Bemühungen und rapid-Prototyping12,13,14,15 ,16. Durch diese Vorteile und die Plug & Play-Natur des Systems ist GFP hat eindeutig ermöglicht eine Vielzahl von Biotechnologieanwendungen wie die Produktion von Proteinen, die schwer zu Schwermetallspuren express- in Vivo17, 18,19,20, Erkennung der Krankheit21,22,23, auf Nachfrage Biomanufacturing18,24 ,25,26,27und Bildung28,29, die zeigen die Flexibilität und den Nutzen der zellfreien Plattform.
GFP-Systeme können aus einer Vielzahl von Rohöl Lysates von beiden prokaryotischen und eukaryotischen Zelle Linien erzeugt werden. Dies ermöglicht vielfältige Möglichkeiten in das System der Wahl, die jeweils vor- und Nachteile je nach Anwendung von Interesse haben. GFP-Systeme sind sehr unterschiedlich in Vorbereitungszeit, Kosten und Produktivität. Am meisten verwendet häufig Zelle, die Extrakte aus Weizenkeimen, Kaninchen Retikulozytenzahl und Insektenzellen Escherichia coli Zellen, wobei letztere die kostengünstigste bisher beim produzieren höchste volumetrische Erträge des Proteins30 hergestellt werden . Während andere GFP-Systeme für ihre angeborene Post-translationale Modifikation Maschinen vorteilhaft sein können, neue Anwendungen mithilfe der E. Coli-basierte Maschinen sind in der Lage, die Lücke durch die Generierung von ortspezifisch phosphorylierten und glykosylierten Proteinen auf Nachfrage31,32,33,34,35.
GFP-Reaktionen können in entweder Charge, kontinuierlichen Austausch zellfreie (CECF) oder kontinuierlichen Fluss zellfreie (CFCF) Formate ausgeführt werden. Das Batch-Format ist ein geschlossenes System, deren Reaktion Lebensdauer aufgrund der abnehmenden Mengen der Reaktionspartner und die Anhäufung von hemmenden Nebenprodukte der Reaktion beschränkt. CECF und CFCF Methoden erhöhen die Lebensdauer der Reaktion und damit höhere volumetrische Protein Renditen im Vergleich zu den Batch-Reaktion führen. Dies geschieht dadurch, dass die Nebenprodukte der Proteinsynthese den Reaktionsbehälter entfernt werden, während neue Reaktionspartner im Laufe der Reaktion2geliefert werden. Bei CFCF kann das Protein des Interesses auch aus der Reaktionskammer, während in CECF, das Protein des Interesses bleibt in der Reaktionskammer, bestehend aus einer halbdurchlässigen Membran36,37entfernt werden. Diese Methoden sind besonders wertvoll bei der Überwindung der schlechten volumetrische Erträge schwer Express Proteine des Interesses38,39,40,41,42, 43. Die Herausforderungen bei der Umsetzung der CECF und CFCF Ansätze sind, dass (1) während sie eine effizientere Nutzung der Bio-Maschinen verantwortlich für Transkription und Translation führen, benötigen sie vor allem größere Mengen an Reagenzien, die Gesamtkosten und 2 erhöht) Sie erfordern komplexere Reaktion Setups und spezialisierte Ausrüstung im Vergleich zu den Batch-Format-44. Um Barrierefreiheit für neue Benutzer zu gewährleisten, beschriebenen die Protokolle hier Fokus auf dem Batch-Format bei Reaktion Volumen von 15 µL mit konkreten Empfehlungen für die Erhöhung des Volumens der Reaktion auf die Milliliter-Skala.
Die hier vorgestellten Methoden ermöglichen Laien mit grundlegenden Labor Fähigkeiten (z. B. Studenten) implementieren Zellwachstum, Vorbereitung zu extrahieren und batch-Format Reaktion Setup für eine E. Coli-basiertes GFP-System. Dieser Ansatz ist kostengünstig im Vergleich zu handelsüblichen Kits ohne Einbußen bei der Einfachheit der Installation Kit-basierte Reaktion. Darüber hinaus ermöglicht dieser Ansatz Anwendungen im Labor und im Feld. Bei der Entscheidung zur Umsetzung der GFP, sollten neue Benutzer die Wirksamkeit der konventionellen Protein Expressionssysteme für Start-up-Investitionen, gründlich auswerten, wie GFP möglicherweise nicht in jedem Fall überlegen. Die hier beschriebenen Methoden GFP ermöglichen dem Benutzer direkt implementieren eine Reihe von Anwendungen, einschließlich der funktionellen Genomik, Hochdurchsatz-Tests, die Produktion von Proteinen, die unlösbar für in Vivo Ausdruck sowie Feld Anwendungen, einschließlich Biosensoren und Bildungs-Kits für synthetische Biologie. Zusätzliche Anwendungen wie z. B. metabolische Technik, Abstimmung von Protein Ausdruck Bedingungen, Krankheit Erkennung, und skalieren mit CECF oder CFCF Methoden sind immer noch möglich, aber erfordern Erfahrung mit der GFP-Plattform für weitere Modifikation der Reaktion Bedingungen. Unsere Methoden kombinieren Wachstum in angereicherten Medien und ratlos Fläschchen mit relativ schnelle und reproduzierbare Methoden der Lyse der Zelle durch Beschallung, gefolgt von einem vereinfachten GFP Reaktion-Setup, das optimierte Vormischungen45nutzt. Während die Zellwachstum Methoden etwas in diesem Bereich standardisiert werden, unterschiedlich Methoden zur Lyse der Zelle. Gemeinsamen Lyse Methoden gehören neben Beschallung Auslastung der französischen Presse, ein Homogenisator, Wulst Schläger, oder Lysozym und anderen Störungen der biochemischen und physikalischen Methoden46,47,48, 49. mit unserer Methode, ca. 2 mL Rohöl Zelle Extrakt pro 1 L Zellen stammen. Diese Menge von Zelle-Extrakt unterstützen vierhundert 15 µL GFP Reaktionen, jedes produzierenden ~ 900 µg/mL Reporter SfGFP Eiweiß aus der Vorlage Plasmid pJL1-SfGFP. Diese Methode kostet $ 0,021/µg von SfGFP hergestellt ($.019/µL Reaktion), ausgenommen die Kosten für die Arbeit und Ausrüstung (ergänzende Abbildung1). Ausgehend von der Pike auf, diese Methode kann von einer einzelnen Person in 4 Tagen umgesetzt werden und wiederholen Sie GFP Reaktionen innerhalb von Stunden (Abbildung 1) abgeschlossen werden können. Darüber hinaus kann das Protokoll in Volumen für größere Chargen von Reagenz Vorbereitung entsprechend den Bedürfnissen des Benutzers skaliert. Wichtig ist, kann die hier vorgestellten Protokoll von Labor ausgebildete Laien wie Studenten, implementiert werden, wie es nur grundlegende Labor Fähigkeiten erfordert. Der nachstehend beschriebenen Verfahren und das dazugehörige Video wurden speziell entwickelt, um die Zugänglichkeit der E. Coli -GFP-Plattform für den breiten Einsatz zu verbessern.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
(1) Media-Vorbereitung und Zellwachstum
-
Tag1
- Streifen BL21*(DE3) E. Coli Zellen aus einem Glycerin auf eine LB-Agar-Platte auf Lager und mindestens 18 h bei 37 ° c inkubieren
- Bereiten Sie 50 mL LB Medien und Autoklaven die Lösung auf einem flüssigen Zyklus für 30 min bei 121 ° C. Bei Raumtemperatur lagern.
-
Tag2
- Bereiten Sie 750 mL 2 X YTP Medien und 250 mL 0,4 M D-Glucose-Lösung, wie in die ergänzenden Informationen beschrieben.
- Gießen Sie 2 X YTP Medien in eine autoklaviert ratlos 2,5 L-Flasche und die D-Glucose-Lösung in einem Autoklaven 500 mL-Glasflasche. Autoklaven beide Lösungen auf einem flüssigen Zyklus für 30 min bei 121 ° C.
- Stellen Sie sicher, dass beide sterilen Lösungen bei 37 ° C gelagert werden, wenn am nächsten Tag, Wachstumsraten bei der Impfung zu maximieren Zellwachstum durchgeführt wird. Kombinieren Sie Lösungen nicht bis Inokulation.
Hinweis: Lösungen können bei 4 ° C für 1-2-d bei Bedarf gespeichert werden, obwohl die 2 X YTP Medien sind sehr anfällig für Verschmutzung. - Starten Sie eine Nacht Kultur des BL21(DE3) durch impfen 50 mL LB Medien mit einer einzigen Kolonie von BL21(DE3) mit einer sterilisierten Schleife und steriler Technik zur Vermeidung von Kontaminationen.
- Die 50 mL BL21*(DE3) LB Kultur in einem 37 ° C 250 u/min schütteln Inkubator und wachsen über Nacht für 15-18 Uhr.
- Vorbereiten und sterilisieren alle Tage 3 und 4, einschließlich der benötigten Materialien: zwei 1 L Zentrifuge Flaschen, 4 X kalt 50 mL konische Rohre (wiegen und Rekord Massen von drei) und viele 1,5 mL Microfuge Röhren.
-
Tag3
- Entfernen Sie die 50 mL über Nacht Kultur der BL21*(DE3) in LB aus dem Schütteln Inkubator und messen die OD600 auf einem Spektrophotometer mit einer 01:10 Verdünnung mit LB Medien. Berechnen Sie das Volumen der Nacht Kultur notwendig, 1 L von Medien für einen Start OD600 von 0,1 hinzuzufügen (z. B. wenn ein OD600 eine 01:10 Verdünnung als 0,4 gelesen wird, impfen 25 mL der unverdünnten OD600 = 4.0 über Nacht Kultur in 1 L 2 X YTP (G).
- Entfernen Sie die erwärmten 2 X YTP Medien und D-Glucose-Lösungen aus dem Inkubator 37 ° C zusammen mit den 50 mL LB-Kultur. Gießen Sie mit steriler Technik, vorsichtig die D-Glucose-Lösung in den 2 X YTP Medien (Vermeidung von den Seiten des Kolbens ratlos).
Hinweis: Zugabe von D-Glucose rundet das Rezept für 1 L 2 X YTPG. - Beibehaltung sterilen Technik, impfen Sie die 1 L 2 X YTPG-Lösung mit der entsprechenden Menge an die 50 mL-Kultur um die 1 L-Kultur bei einer 0.1 OD600zu beginnen. Sofort setzen Sie die beimpften 1 L Kultur in einem 37 ° C Inkubator bei 200 u/min schütteln.
- Nehmen Sie die erste OD600 lesen nach der ersten Stunde des Wachstums (Lag-Phase typische dauert 1 h). Verdünnen Sie die Kultur nicht. Die Einnahme OD600 Messungen etwa alle 20-30 min bis OD600 0,6 erreicht.
- Beim erreichen von OD600 = 0,6, fügen Sie 1 mL 1 M IPTG (Endkonzentration in 1 L Kultur = 1 mM), 2 X YTPG-Kultur.
Hinweis: Ideal Induktion OD600 ist 0,6; eine Reihe von 0,6-0,8 ist jedoch akzeptabel. Induktion mit IPTG ist für die körpereigene Produktion der T7 RNA-Polymerase (T7RNAP). - Messen Sie nach Induktion die OD600 ca. alle 20-30 min bis 3.0 erreicht.
Hinweis: Abkühlen der Zentrifuge auf 4 ° C während dieser Zeit. Bereiten Sie kalt S30 Puffer wie in den Zusatzinformationen. Wenn der S30-Puffer im Voraus vorbereitet ist, sicherzustellen Sie, dass DVB-t nicht bis zum Tag der Anwendung hinzugefügt wird. - Sobald die OD600 3.0 (Abbildung 2A) erreicht, Gießen Sie die Kultur in eine kalte Flasche 1 L Zentrifuge in einer Eis-Wasserbad. Bereiten Sie eine wassergefüllte 1 L Zentrifuge Flasche gleich Gewicht als ein Gleichgewicht in der Zentrifuge verwendet werden.
Hinweis: Absorption Werte variieren von Instrument zu Instrument. Während die OD600 der Ernte des BL21(DE3) keine sensible Variable ist, empfiehlt es sich, dass der Benutzer bewerten und optimieren diese Variable als Maßnahme zur Fehlerbehebung. Größere Spektralphotometer können in relativ niedriger OD600 Lesungen im Vergleich zu kleineren Küvette-basiertes Spektralphotometer führen. - Zentrifugieren der 1 L-Flaschen für 10 min bei 5.000 x g und 10 ° C, pellet-Zellen.
- Gießen Sie langsam aus dem überstand und entsorgen es nach biologischen Abfällen Verfahren des Instituts. Legen Sie das Pellet auf Eis.
- Mit einem sterilen Spatel, kratzen Sie die Zelle Pellet aus der Zentrifuge Flasche und auf eine kalte 50 mL konische Rohr übertragen.
- Das konische Rohr 30 mL kalte S30 Puffer hinzu und Aufschwemmen der Zelle Pellet durch Vortexen mit kurzen Stößen (20-30 s) und Ruhezeiten (1 min) auf dem Eis bis voll mit keine Klumpen Nukleinsäuretablette.
- Sobald das Pellet vollständig Nukleinsäuretablette ist, verwenden Sie ein weiteres 50 mL konische Rohr mit Wasser als Ausgleich und Zentrifuge für 10 min bei 5000 x g und 10 ° C (bis 4 ° C vorgekühlt).
Hinweis: Dies schließt die 1St 3 Waschgänge erforderlich, wenn die Zellen zu ernten. - Gießen Sie den Überstand und entsorgen Sie es entsprechend der Institution biologischen Abfällen Verfahren. Aufzuwirbeln Sie das Pellet mit 20-25 mL kalte S30 Puffer und Zentrifuge für 10 min bei 5000 X g und 10 ° C (bis 4 ° C vorgekühlt).
Hinweis: Dies schließt die 2Nd 3 Wäschen. - Wieder, Gießen Sie überstand und entsorgen Sie es entsprechend der Institution biologischen Abfällen Verfahren. Hinzugeben Sie genau 30 mL S30 Puffer und Wirbel wieder an das Pellet Aufschwemmen.
- Mit Hilfe der 3 vorgewogene, kalten 50 mL konische Röhrchen und des Füllstoffes serologische Pipette mit einer sterilen Pipette, aliquoten 10 mL der resuspendierte Pellet/S30 Puffer Mischung in jedes der 3 konische Rohre.
Hinweis: Die Teilung der Zellen in 3 Röhren ist nicht erforderlich, aber dieser Schritt führt zu kleineren Zelle Pellets (~ 1 g) für mehr Komfort in späteren Schritten. - Zentrifugieren Sie alle Röhren, mit entsprechenden Salden Bedarf für 10 min bei 5000 x g und 10 ° C (vorgekühlt auf 4 ° C).
Hinweis: Damit ist der letzten Waschschritt abgeschlossen. - Gießen Sie den Überstand und entsorgen Sie es entsprechend der Institution biologischen Abfällen Verfahren. Entfernen Sie die überschüssige S30-Puffer durch das Innere der konische Rohr und Verschluss mit einem sauberen Tuch sorgfältig abwischen; Berühren Sie das Pellet.
- Die Röhren auf einer Analysenwaage Vakuumtrockenschrank und nehmen die endgültige Pellet Gewicht auf jedes Rohr.
Hinweis: Das Protokoll kann an dieser Stelle angehalten werden. Die Pellets können Flash-werden in flüssigem Stickstoff eingefroren und bei-80 ° C bis zu einem Jahr bis für Extrakt Vorbereitung erforderlich gespeichert.
(2) grobe Zelle extrahieren Vorbereitung - Tag 4
- Vorbereitung der Extrakt halten Sie Zellen kalt auf dem Eis bei jedem Schritt. Fügen Sie 1 mL kalte S30 Puffer pro 1 g Zellmasse des Pelletofens. Sicherzustellen Sie, dass diese Dithiothreitol (DTT) auf den Puffer, S30, eine Endkonzentration von 2 mM ergänzt wurde.
Hinweis: Abkühlen der Microcentrifuge bis 4 ° C während dieser Zeit. - Aufschwemmen der Zelle Pellet durch Vortexen mit kurzen Stößen (20-30 s) und Ruhezeiten (1 min) auf dem Eis bis voll Nukleinsäuretablette. Wenn Wiederfreisetzung schwierig ist, lassen Sie die Pellets für 30 min zum Auftauen auf Eis.
- Übertragen Sie 1,4 mL der resuspendierte Zellen in einem 1,5 mL Microfuge Rohr.
- Legen Sie eine 1,5 mL Tube mit 1,4 mL der resuspendierte Zellen in ein Eis-Wasserbad in ein Becherglas. Beschallen für 45 s auf gefolgt von 59 s aus 3 insgesamt Zyklen mit Amplitude auf 50 % eingestellt. Schließen und die Röhrchen vorsichtig mischen aus Zeiten zu invertieren. Insgesamt liefern Sie 800-900 J Energie an jede 1,5 mL Microfuge Röhre mit 1,4 mL der resuspendierte Zellen (Abb. 3A und 3 b).
Hinweis: Dieser Schritt ist empfindlich gegenüber der Sonikator Typ und Modell verwendet und sollten optimiert werden, wenn Geräte anders als für dieses Verfahren aufgeführt ist. Zwei sich ergänzende Ansätze können verwendet werden, um Scale-up-die Menge an Extrakt bei diesem Schritt vorbereitet: (1) mehrere 1,5 mL Microfuge Rohre parallel beschallt werden können und/oder (2) größere Mengen in konischen Rohren beschallt werden können (bis zu 15 mL Zelle Wiederfreisetzung pro Rohr) , wie oben beschrieben Skalierung der gelieferten Energiemenge, 29,45. - Unmittelbar nach Beschallung abgeschlossen ist, fügen Sie 4,5 µL 1 m (zur Ergänzung einer zusätzliche 2 mM DVB-t) DVB-t in der 1,4 mL lysate und invertieren Sie mehrmals zu mischen. Platzieren Sie das Rohr auf dem Eis. Wiederholen Sie die Schritte 2.4 und 2.5 für jede zusätzliche Rohre der resuspendierte Zellen vor der Zentrifugation.
- Microcentrifuge Proben bei 18.000 x g und 4 ° C für 10 min (Abbildung 3).
- Pipette des Überstands in ein neues 1,5 mL Microfuge Rohr. Stören Sie das Pellet nicht; Es empfiehlt sich, einige überstand hinterlassen weiterhin Reinheit als zu stören das Pellet in Bemühungen um die Rendite zu maximieren.
- Inkubieren Sie überstand aus dem vorherigen Schritt auf 250 u/min und 37 ° C für 60 min durch Abkleben der Rohre, die schütteln Plattform des Inkubators (Dies ist der Abfluss-Reaktion).
- Microcentrifuge Proben bei 10.000 x g und 4 ° C für 10 Minuten.
- Den Überstand zu entfernen, ohne zu stören das Pellet und überträgt es auf einen neuen Schlauch. Viele 100 µL-Aliquots der Extrakt für die Lagerung zu schaffen.
Hinweis: Das Protokoll kann hier angehalten kann, und des Extrakts flash, in flüssigem Stickstoff eingefroren und bei-80 ° C bis zu einem Jahr bis zur GFP Reaktionen benötigt gespeichert. Mindestens 5 Gefrier-Tau-Zyklen können ohne Beeinträchtigung der Produktivität (Abbildung 4) extrahieren absolviert werden.
(3) zellfreien Protein Synthesereaktionen Batch-Format
- Tauen Sie Lösungen A und B, DNA-Vorlage BL21(DE3) Extrakt (wenn gefroren), T7RNAP und ein Aliquot der molekularen Grade Wasser auf.
Hinweis: GFP Reaktion Vorlage finden Sie in den Zusatzinformationen. Lösungen A und B Rezepte werden in den Zusatzinformationen zur Verfügung gestellt und entsprechen bestimmten Konzentrationen für zahlreiche Reagenzien, PANOx-SP-basierte Energiesystem für GFP zu unterstützen. Die Rolle der einzelnen Reagenz und akzeptable Variation in diesen Reagenz-Konzentrationen, die GFP unterstützen kann schon bestimmt50. Ein T7RNAP-Reinigung-Protokoll finden Sie in den Zusatzinformationen51. Ergänzende T7RNAP volumetrische Erträge zu steigern, aber ist nicht erforderlich, wenn T7RNAP beim Zellwachstum induziert wird. Plasmid DNA Schablone (pJL1-SfGFP) kann mit einem Maxiprep-Kit mit zwei Wäschen mit dem Waschpuffer im Kit, gefolgt von einer Nachbearbeitung DNA-Bereinigung mit einem PCR-Reinigung-Kit (Abb. 2 b) vorbereitet werden. Lineare DNA-Vorlagen können auch im GFP Reaktionen verwendet werden. - Beschriften Sie die notwendige Menge an Microfuge Rohre für GFP Reaktionen benötigt.
Hinweis: Reaktionen in verschiedenen Schiffsgrößen durchgeführt werden können, aber ein kleineres Schiff kann volumetrische Protein Erträge (Abbildung 2) verringern. Skalierung bis eine Reaktion in der gleichen Größe Gefäß kann auch volumetrische Erträge als Funktion der Verringerung der Sauerstoffaustausch auf einen Rückgang der Fläche zum Volumenverhältnis reduzieren. Wenn über 100 μl Reaktionsvolumen zu erhöhen, empfiehlt es sich, flachen Boden well-Platten 31,37,52zu verwenden. - Fügen Sie 2.2 µL der Lösung A, 2.1 µL Lösung B, 5 µL BL21*(DE3) extrahieren, 0,24 μg des T7RNAP (16 μg/mL Endkonzentration), 0,24 ng DNA Schablone (16 ng/mL Endkonzentration) und Wasser 15 µL Endvolumen bringen.
Hinweis: Vortex Lösungen A und B häufig während des Setups Reaktion zur Sedimentation der Bauteile zu vermeiden und sicherzustellen, dass jede Reaktion eine homogene Aliquot der jeweiligen Lösung erhält. Vermeiden Sie Aufschütteln der Extrakt, stattdessen Invertieren der Röhre zu mischen. - Nachdem alle Reagenzien zur Reaktion hinzugefügt wurden, mischen Sie jedes Rohr durch Pipettieren rauf und runter oder sanft aufschütteln gleichzeitig sicherzustellen, dass die endgültige Reaktionsgemisch in einen einzigen 15 µL Wulst am unteren Rand der 1,5 mL Microfuge Tube kombiniert wird.
- Stellen Sie jede Reaktion in den Inkubator 37 ° C, ohne Zittern für 4 h oder 30 ° C über Nacht.
Hinweis: Erfolgreiche Reaktionen können qualitativ optisch basierend auf die grüne Farbe des SfGFP Produkts innerhalb der GFP Reaktionsgemisch (Abbildung 3D) beurteilt werden. Expression des Proteins des Interesses kann auch durch SDS-PAGE (ergänzende Abbildung2) bestätigt werden.
(4) Quantifizierung der Reporter Protein, [SfGFP]
- Laden Sie 48 µL von 0,05 M HEPES, pH 8, in jeder gut zur Quantifizierung (in der Regel in dreifacher Ausfertigung pro Reaktionsgefäß durchgeführt).
- Reaktionen aus Inkubator zu entfernen. Pipette rauf und runter um jede Reaktion zu mischen, dann 2 µL der Reaktion in den 48 µL von 0,05 M HEPES, pH 8 übertragen. Pipettieren oben und unten wieder in den Brunnen zu mischen.
- Sobald alle Reaktionen geladen und vermischt sind, legen Sie die 96-well-Platte in den Fluorometer und Messen Sie die SfGFP Endpunkt Fluoreszenz.
Hinweis: Anregung und Emission Wellenlängen für SfGFP Fluoreszenz Quantifizierung sind 485 nm und 510 nm, beziehungsweise. - Anhand einer zuvor generierten Standardkurve fest [SfGFP] aus den erhaltenen Fluoreszenz-Lesungen.
Hinweis: Anweisungen zum Generieren einer Standardkurve SfGFP Konzentration im Vergleich zu Fluoreszenzintensität sind in Zusatzinformationen (ergänzende Abbildung 3) ausgestattet. Benutzer müssen eine Standardkurve für ihr Instrument zu etablieren, da Instrument Sensibilität variieren.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Wir haben ein Ultraschall-basierte E. Coli Extrakt präsentiert Vorbereitung-Protokoll, das über einen Zeitraum von vier Tagen mit Abbildung 1 zeigt die verfahrensrechtliche Aufschlüsselung über jeden Tag abgeschlossen werden kann. Gibt es Formbarkeit zu den Schritten, die jeden Tag mit verschiedenen anhalten Punkten abgeschlossen werden können, aber wir haben festgestellt, dass dieses Workflows, die effektivste ausgeführt werden. Darüber hinaus sind die Zelle Pellets (Schritt 1.3.18) und gut vorbereitet-Extrakt (Schritt 2.10) stabil bei-80 ° C für mindestens ein Jahr, ermöglicht dem Benutzer, größere Bestände der einzelnen für den Einsatz bei einer späteren Zeit17speichern zu erstellen. Nicht nur der Extrakt über lange Zeiträume stabil ist, sondern der Extrakt kann auch mindestens fünf Einfrieren Auftauen Zyklen ohne einen erheblichen Verlust an Produktivität (Abbildung 4) unterziehen. Dies ermöglicht größere Aliquote Extrakt für verschiedene Zwecke gespeichert werden, wenn der Gefrierschrank Stauraum begrenzt ist. Wir empfehlen jedoch mehrere kleinere Aliquote (~ 100 μL) Extrakt, wann immer möglich.
Mit jeder neuen Auszug-Vorbereitung empfehlen wir, dass der Benutzer eine Magnesium-Titrierung durchführt, um die optimale Menge an Magnesium für diesen Batch-Extrakt zu bestimmen. Benutzer können Variabilität von Charge zu Charge in Gesamt-Protein-Konzentration des Extraktes Zelle von Bradford Assay quantifizieren. Für leistungsfähigere Extrakte, wir in der Regel insgesamt Proteinkonzentrationen von 30-50 mg/mL sehen, und innerhalb dieses Bereichs gibt es keine offensichtliche Korrelation zwischen Gesamt-Protein-Konzentrationen und Extrakt Zellleistung. Daher wird empfohlen, dass Benutzer Magnesium-Konzentrationen entsprechend einstellen, um sicherzustellen, dass Protein und Nukleinsäure-Funktionalität für jede Charge Extrakt maximiert werden. Magnesiumspiegel sind wichtig für die korrekte DNA-Replikation, Transkription und Translation können, übermässigem jedoch nachteilig für diese Prozesse53. Um diese Abhängigkeit zu zeigen, wir haben eine Co-Titrierung von Magnesium durchgeführt und extrahieren Volumen um die optimale Kombination zu ermitteln, die die Menge an Extrakt erforderlich, wobei eine produktive Reaktion (Abbildung 5) minimiert. Aus diesem Experiment empfehlen wir 5 µL-Extrakt und 10 mM Mg2 + für Extrakt mit einem Gesamtproteingehalt von 30 mg/mL, um mehr als 1.000 μg/mL des SfGFP zu erhalten.
Unsere Erfahrung mit GFP hat uns Schritte innerhalb des Protokolls, die ohne Nachteil für die Gesamtproduktivität des Systems variiert werden kann, und andere zu bestimmen, die für eine hohe Leistung GFP-System sind auch erlaubt. Vor allem die letzte OD600 der Zellernte wirkt sich nicht deutlich die endgültige Ausgabe der GFP-Reaktion, und Zellen können praktisch überall von 2,7-4,0 OD600geerntet werden. Dies entspricht die frühen exponentielle Wachstumsphase wo Ribosom Konzentration pro Zelle ist die höchste und die translationale Maschinen ist am aktivsten, schnelles Wachstum zu unterstützen. Diese Beobachtung ermöglicht die Flexibilität, ihren eigenen Verfahren zu optimieren. Wir empfehlen Ernte bei ca. 3,0 OD600 um die Zellen bei einer OD600 näher auf 3.3 durch die Zeit ernten zu erfassen ist abgeschlossen (Abb. 2A). Variablen, die GFP Erträge Auswirkungen haben gehören Vorlage DNA-Qualität, Reaktion Behältergröße und die relativen Mengen der Zelle-Extrakt und Magnesium-Ionen in die Reaktion vorhanden. Wir haben DNA-Qualität haben bemerkenswerte Variation von Charge zu Charge gefunden. Um dieses Problem zu beheben, empfehlen wir, dass Benutzer reinigen DNA über Midi oder Maxi Prep, gefolgt von einer zusätzlichen DNA Aufräumsegment entweder auf der DNA-Reinigung-Spalte in der Maxiprep verwendet, oder Post-Reinigung mit einer zusätzlichen DNA-Bereinigung Kit. Dies verbessert die Reproduzierbarkeit in der DNA-Qualität für GFP Reaktionen und Ergebnisse in robuster Protein-Produktion (Abbildung 2 b). Der Reaktionsbehälter wirkt sich auch volumetrische Erträge, so dass die Proteinproduktion identische Reaktion Setups in unterschiedliche Behälter-Volumen bis zu 40 % abweichen kann. Es hat die Theorie, dass die Steigerung der volumetrischen Ertrag in größeren Gefäßen beobachtet ist durch eine vergrößerte Oberfläche der Reaktionsmischung, zulassend bessere Sauerstoffaustausch (Abbildung 2), und andere weiter volumetrische Erträge durch Ausführen steigerte haben GFP-Reaktionen in großen flach-Boden-Platten, die wir für Reaktionen über 100 µL17,31,37,52empfehlen.

Abbildung 1 : Zeitplan für Kultur Wachstum, Herstellung von Zelle-Extrakt, Setup und Quantifizierung der GFP Reaktionen. Der Benutzer kann die GFP-Plattform für ihre Anwendungen in der Forschung durch dieses viertägigen Workflow implementieren. Reagenz Vorbereitung steht für die primäre Zeit und Kosten Investitionen für die erste Runde dieses Experiments und vermindert erheblich nachdem Reagenzien Bestände festgelegt sind. Darüber hinaus können Zelle Pellets und vorbereitete Zelle Extrakt für über ein Jahr bei-80 ° C, so dass der Benutzer das Ablaufprogramm an verschiedenen Schritte für schnellere Ergebnisse beginnen gespeichert werden. Die Benutzer kann auch in verschiedenen Schritten zum Ändern der Zeitachse dieses Workflows anhalten. Bitte klicken Sie hier für eine größere Version dieser Figur.

Abbildung 2 : Veränderbaren Bedingungen für GFP und die Auswirkungen auf die Renditen der volumetrischen Reaktion. A. Auszug Produktivitätsvergleich je nach Ernte BL21(DE3) Zellen bei verschiedenen OD600 Lesungen. Basierend auf diesem Grundstück, empfehlen wir die Ernte bei einer OD600 von 3,3, mindestens 1000 µg/mL Zielprotein zu produzieren. Reaktionen wurden in einem 15 μl Maßstab in 1,5 mL Microfuge Rohre durchgeführt. B. Vergleich der beiden DNA-Maxiprep waschen Protokolle mit und ohne Post-Reinigung DNA-Aufräumarbeiten. pJL1-SfGFP Plasmide wurde ein Maxiprep mit ein oder zwei Wäschen, gefolgt von einer Post-Reinigung-Bereinigung durch PCR-Reinigung-Kit. Um ~ 900 µg/mL Protein-Expression zu erreichen, empfiehlt es sich, eine Post-Reinigung DNA Bereinigung unabhängig von der Anzahl der Maxiprep wäscht. Reaktionen wurden in einem 15 μl Maßstab in 1,5 mL Microfuge Rohre durchgeführt. C. 15 µL GFP Reaktionen durchgeführt in verschiedenen Gefäßen von 2 mL bis 0,6 mL Microfuge Röhren. "Neg" stellt eine Negativkontrolle, wo die Reaktion keine DNA-Vorlage hinzugefügt wurde. Alle Fehlerbalken darzustellen 1 Standardabweichung von drei unabhängigen Reaktionen für jede Bedingung, von denen jede in dreifacher Ausfertigung quantifiziert wurde. Bitte klicken Sie hier für eine größere Version dieser Figur.

Abbildung 3 : Schlüssel prozedurale Setups und Ergebnisse für die Schaffung produktiver zu extrahieren. A. korrekte Einrichtung der Beschallung Eiswasser Bad sorgt für Kühlung der Probe, während Wärme während der Anwendung von Ultraschall erzeugt wird. B. 1,5 mL Microfuge Röhrchen mit resuspendierte Zelle Pellet vor (links) und Post (rechts) Beschallung. Die daraus resultierenden lysate sollte einen dunkleren Farbton im Vergleich zu Pellet resuspendierte Zelle angezeigt werden. C. ordnungsgemäße Trennung des Überstandes und Pellet Zelle lysate nach 18.000 x g zentrifugieren. D. GFP Reaktionen nach 4 h Inkubation bei 37 ° C. 1,5 mL Microfuge Schlauch auf der rechten Seite (erfolgreiche Reaktion) zeigt sichtbare Fluoreszenz des SfGFP Reporter Proteins bei ~ 900 μg/mL. Die Negativkontrolle Röhre auf der linken Seite, fehlt Vorlage DNA und simuliert eine erfolglose Reaktion zeigt eine klare Lösung mit keine Fluoreszenz. Bitte klicken Sie hier für eine größere Version dieser Figur.

Abbildung 4 : Veränderung der Proteinexpression über 5 Gefrier-Tau-Zyklen für GFP extrahieren. Auszug aus der gleichen Wachstum vorbereitet unterzog sich fünf Auftau-Zyklen über Flüssigstickstoff Blitz Einfrieren gefolgt von Auftauen auf Eis einfrieren. Über die fünf Gefrier-Tau-Zyklen wurden keine wesentlichen Änderungen im Extrakt Produktivität für den Ausdruck von SfGFP gesehen. Reaktionen wurden in einem 15 μl Maßstab in 1,5 mL Microfuge Rohre durchgeführt. "Neg" stellt eine Negativkontrolle, wo die Reaktion keine DNA-Vorlage hinzugefügt wurde. Alle Fehlerbalken darzustellen 1 Standardabweichung von drei unabhängigen Reaktionen für jede Bedingung, von denen jede in dreifacher Ausfertigung quantifiziert wurde. Bitte klicken Sie hier für eine größere Version dieser Figur.

Abbildung 5 : GFP für Reaktionen mit unterschiedlichen [Mg2+] und extrahieren Volumen im Vergleich zu [SfGFP]. [Mg2 +] reichte von 8 mM bis 14 mM mit 2 mM-Schritten und Volumina reichte von 3 µL bis 7 µL mit 1 µL Schritten zu extrahieren. Die Farbe Code stellt [SfGFP] hergestellt aus hoch (rot) zu niedrig (lila). Um Reagenz Effizienz zu maximieren und gleichzeitig hohen Protein-Produktion, wir empfehlen 5 µL des Extraktes und 10 mM Mg2+ für Extrakte, die ein Gesamtproteingehalt von ~ 30 mg/mL, wie von Bradford-Test bestimmt. Ursprünglichen Punkte um die Konturdiagramm zu generieren wurden Endpunkt Fluoreszenz von drei unabhängigen Reaktionen für jede Bedingung basiert auf die jeweils in dreifacher Ausfertigung gemessen wurde. Reaktionen wurden in einem 15 μl Maßstab in 1,5 mL Microfuge Rohre durchgeführt. Bitte klicken Sie hier für eine größere Version dieser Figur.
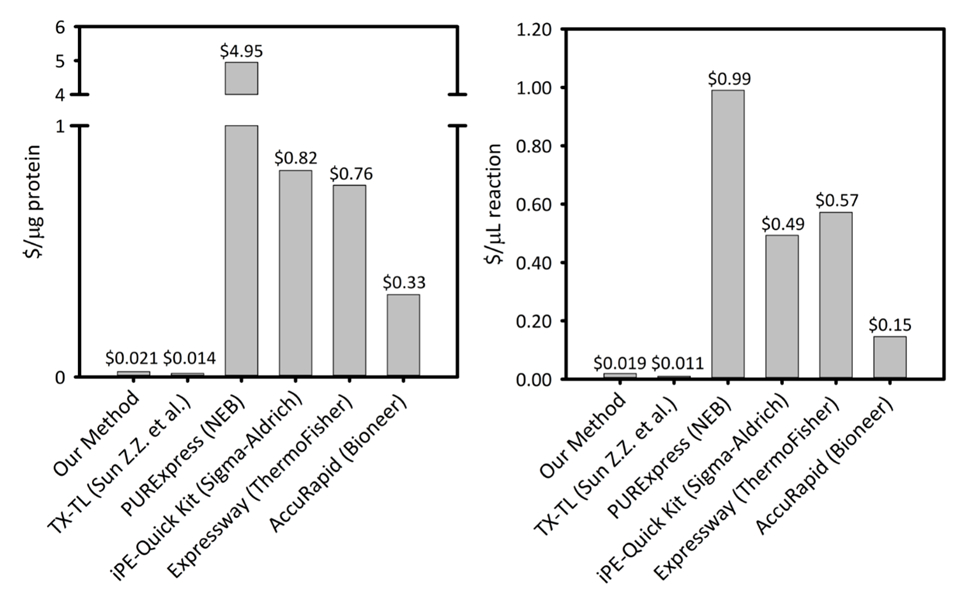
Zusätzliche Abbildung 1: Kosten pro Mikroliter der Reaktion und Mikrogramm Protein produziert sechs zellfreien Protein Synthese plattformübergreifend. Unsere Plattform wird auf fünf verschiedenen Zelle freie Protein Synthese Kits/Plattformen mit unterschiedlichen Produktivität und Preise verglichen. Unsere Beschallung basierte GFP Plattform ist kostengünstiger als die meisten kommerziellen kits und die Leichtigkeit eines Bausatzes für Reaktion Setup während der verbleibenden Kosten vergleichbar mit anderen akademischen GFP-Plattformen bietet in beiden $/ µg Protein und $/ µL der Reaktion. Bitte klicken Sie hier für eine größere Version dieser Figur.
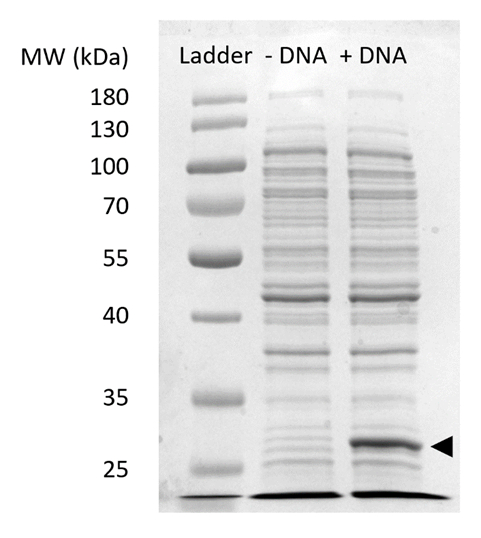
Zusätzliche Abbildung 2: SDS-PAGE des SfGFP Ausdrucks im GFP. Zellfreien Protein Synthesereaktionen mit (+ DNA) und ohne (-DNA) DNA Schablone für SfGFP wurden laufen auf 12 % SDSPAGE Gel zu zeigen, die Expression von SfGFP bei 27 kDa (schwarzer Pfeil) beobachtet. Traditionelle Techniken der SDS-PAGE verwendet wurden. Jede Probe verladen des Gels enthalten 18 µg von Gesamtprotein basierend auf Bradford Assay Quantifizierung der Gesamt-Protein in der Zelle zu extrahieren. Basierend auf Fluoreszenz Intensität Messungen und unsere Standardkurve, wir schätzen, dass die "+ DNA" Spur enthält 0,42 µg des SfGFP. Um diese Proben zu erhalten, wurden in einem 15 µL Maßstab in 1,5 mL Microfuge Röhren volumetrische Erträge gemäß Abbildung 3GFP Reaktionen ablaufen. Bitte klicken Sie hier für eine größere Version dieser Figur.
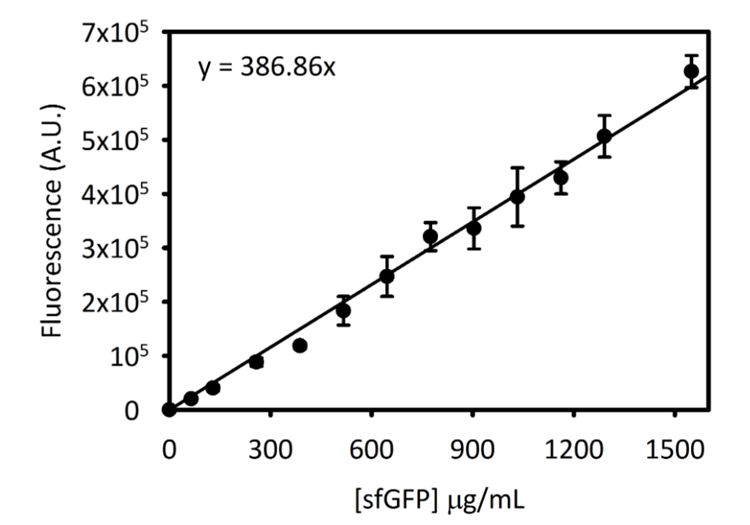
Ergänzende Abbildung 3: Standardkurve für SfGFP auf Cytation 5. Diese Kurve wurde über die oben beschriebenen Methoden ermittelt. Alle Daten für diese Handschrift von Endpunkt Fluoreszenz Lesungen in [SfGFP] umgewandelt wurde in µg/mL mit diesem Standardkurve gesammelt. Bitte klicken Sie hier für eine größere Version dieser Figur.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Zellfreie Proteinsynthese ist eine leistungsfähige und förderlichen Technologie für eine Vielzahl von Anwendungen von Biomanufacturing bis hin zu rapid Prototyping von biochemischen Systemen entstanden. Die Bandbreite der Anwendungen stützt sich auf die Fähigkeit zu überwachen, zu manipulieren und zellulären Maschinerie in Echtzeit zu erweitern. Trotz der wachsenden Auswirkungen dieser Plattform-Technologie, breiten Anpassung blieb langsam durch technische Feinheiten bei der Umsetzung der Methoden. Durch diese Bemühungen wollen wir Einfachheit und Klarheit für die Festlegung dieser Technologie in neue Labs bieten. Zu diesem Zweck unser Protokoll für ein E. Coli-basierte zellfreien Protein Synthese Plattform innerhalb einer Startzeit von vier Tage durch Labor ausgebildete Laien, wie Studierende (Abbildung 1) erreicht werden kann. Darüber hinaus einmal ein Bestand von Reagenzien und Extrakt hergestellt werden, nachfolgende GFP Batch Reaktionen können werden eingerichtet, inkubiert und in nur 5 h quantifiziert. Einem einzigen kann 1 L Zellwachstum führen genügend Extrakt für vierhundert 15 µL GFP Reaktionen, während einzelne Charge Vorbereitungen der anderen zellfreie Reagenzien Tausende von Reaktionen unterstützen können. Reagenz Vorbereitungen können auch skaliert, wenn eine noch größere Lager benötigt wird. Die GFP-Reaktionen können in gewissem Hochdurchsatz-werden mithilfe einer 96-Well-Platte oder PCR-Röhrchen für die Prüfung von einer Vielzahl von Bedingungen parallel. Volumetrische Erträge sinken, wenn mit kleineren Schiffen wie in Abbildung 2zu sehen. GFP Reaktionen können auch von Mikroliter bis zu zehn Milliliter total Reaktionsvolumen skaliert werden um die Gesamt-Protein-Ausbeute für eine einzelne Bedingung zu erhöhen. Bei der Skalierung Lautstärke ist der vorrangig zu berücksichtigen, dass volumetrische Reaktion Erträge sinken, abnehmender Oberfläche Bereich-zu-Volumen-Verhältnis der Reaktion37,52. Um Scale-Up und gleichzeitig ähnliche volumetrische Erträge der Proteinexpression Benutzer sollten das Reaktionsvolumen in zahlreichen Reaktionsgefäßen aufgeteilt und/oder Behältergröße zu erhöhen. Für Reaktion Skalen von 15 µL - 100 µL Volumen, sind zahlreiche 15 µL Reaktionen parallel empfohlen. Für Reaktionen von mehr als 100 µL Volumen flach-Boden-24-Well-Platten empfohlen, und 12-Well-Platten sind für die Reaktion Volumen von mehr als 600 µL empfohlen. Solche Paarungen der Reaktion Volumen und Schiffe bieten Konsistenz der volumetrischen Reaktion Renditen auf Scale-up-17,31,37,52. Jenseits dieser Bände Skalierung kann durch die Verwendung mehrere Vertiefungen der Platte parallel erfolgen. Dieses Format verwenden, kann die Reaktion auf mehr als 10 mL Gesamtvolumen skaliert werden. Optimierung der Reaktion Volumen-Reaktion Schiff Kombination kann Anwendungen von Biomanufacturing unterstützen, ohne Einbußen bei der Produktivität der Reaktion.
Bei der Durchführung dieses Protokolls gibt es ein paar wichtige Überlegungen, die volumetrische Reaktion Erträge sowie Indikatoren zugeordnet Extrakt mit schlechter Leistung auswirken. Zur Sicherstellung der ordnungsgemäßen lyse und Denaturierung der funktionalen Transkription/Übersetzung Maschinen zu verhindern, ist es wichtig, die Wärmeentwicklung während Lyse zu mildern. Tauchen Sie die Zelle Wiederfreisetzung in ein Eis-Wasserbad während der Beschallung, schnell Wärme während der Beschallung (Abb. 3A). Ein Indikator für die effektive Zelle Lyse ist die Entstehung von ein dunkleres Aussehen der Zelle lysate im Vergleich zum Pre-sonorisiert Proben (Abb. 3 b). Für Flexibilität für Benutzer ist die Sonikator und Sonde, dargestellt in Abbildung 3A anpassungsfähig zu einer Reihe von Bänden von 100 µL bis 15 mL resuspendierte Zellen. Um dies zu erreichen, kann der Benutzer die Anzahl der Joule zur Lyse der Zellen die gewünschte Lautstärke anpassen. Darüber hinaus können größere Mengen an Extrakt durch zwei sich ergänzende Ansätze vorbereitet werden. Benutzer können mehrere Rohre parallel beschallen bzw. beschallen größere Volumen der Zelle Wiederfreisetzung Skalierung die Menge an Energie proportional mit dem Volumen als zuvor beschriebenen29,45. Ein weiterer Schritt, der angibt, Extraktqualität ist die Zentrifugationsschritt nach lyse der Zelle. Post Zelle Lysis, wir empfehlen Zentrifugation bei 18.000 x g, eine klare Trennung zwischen der Überstand (Transkription/Übersetzung Maschinen, fragmentierte genomischer DNA die Funktionen nicht mehr zur Vorlage Transkription/Übersetzung) bereitzustellen und das Pellet ( unerwünschte zelluläre Komponenten wie die Zellmembran und die ausgefällten Proteine) (Abbildung 3). Wir haben festgestellt, dass Zentrifugation bei 18.000 x g verbessert Trennung, wodurch verbesserte Reproduzierbarkeit im Vergleich zu Drehungen mit niedrigeren Geschwindigkeiten wie 12.000 x g. Der Einfachheit halber empfehlen wir eine Tischplatte gekühlte Zentrifuge, mindestens 12.000 x g erreichen kann. Dieser Schritt wird auch häufig bei 30.000 x g ausgeführt, die berücksichtigt werden sollten, ist die geeignete Ausrüstung zur Verfügung54,55,56,57,58, 59 , 60. Extrakt Leistung ist durch Zentrifugation Geschwindigkeiten bei diesem Schritt nicht beeinträchtigt, da die ordnungsgemäße Trennung erreicht wird. Wenn Sie den gewünschten überstand entfernen, empfiehlt es sich, keine trüben Materialien zu vermeiden, die an der Grenze zwischen dem überstand und Pellet vorhanden sind, da diese Kontamination wird die Produktivität des Extraktes reduzieren. Streben nach Reinheit des Überstands führt zu produktiver Extrakte und lohnt sich die reduzierte Menge an Extrakt für neue Benutzer.
Es ist wichtig zu beachten, die während der Methoden, die wir vorgestellt haben reproduzierbar sind und von Wissenschaftlern mit minimalen Kenntnissen ausgeführt werden können Charge zu Charge werden kann und Reaktion zu Reaktion Variation. Dies kann zu Schwankungen in der Proteomik-Zusammensetzung der lysate Post-Beschallung61zurückführen. Die Variabilität von Charge zu Charge, die wir beobachtet haben ist in der Regel nach Supplementierung mit T7RNAP und Optimierung der Magnesium-Konzentration vermindert. Exogene Zugabe von T7RNAP ist häufig bei GFP Reaktionen auf optimale Protein-Expression zu unterstützen, und wir finden, dass mit zwei Quellen der T7RNAP - endogenen Ausdruck in BL21*(DE3) und die zusätzliche T7RNAP, eine Endkonzentration von 16 µg/mL - verbessert Charge zu Charge Reproduzierbarkeit für neue Benutzer45,46. Mit Erfahrung können Benutzer ihre Experimente um nur eine einzige Quelle von T7RNAP nutzen zu können, falls gewünscht ändern. Quantifizierung der Gesamtproteingehalt eine neue Charge von Extrakt und entsprechende Anpassung der Mg2 + Konzentration kann auch helfen, um Charge zu Charge Variation der volumetrischen Protein Ausdruck Renditen zu vermindern. Variationen in Protein-Expression kann auch aufgrund von Unterschieden in der Größe und Struktur des Proteins des Interesses, das Codon Usage des Gens und seine entsprechenden Ribosom Bindungsstelle des Gens von Interesse, sowie die Art der Expressionsvektor verwendet62 ,63. Aus diesen Gründen können einige Proteine nicht sowie das Modell Protein SfGFP, wodurch volumetrische Ertragseinbußen von GFP Reaktionen äußern.
Einschränkungen der vorgestellten GFP Technik beinhalten, dass es möglicherweise nicht direkt geeignet für alle Anwendungen der zellfreien, z. B. metabolische Technik und tuning der Ausdruck Bedingungen, ohne weitere Änderungen zu den Protokollen. Wir glauben jedoch, dass dieses Protokoll wird eine Grundlage bieten für die Festlegung der GFP-Plattform in Laboratorien und Laien mit der Fähigkeit, einleitende zellfreie Reaktionen in ihren Laboren zu implementieren. Nach der anfänglichen Implementierung können die Plattform, um ihre eigenen Änderungen für spezielle Anwendungen basierend auf andere Literatur im Bereich vornehmen Forscher experimentieren.
Das GFP-Plattform Kosten $ 0,021/µg Protein (mit Ausnahme der Kosten der Arbeit und Ausrüstung), macht unser System preisgünstig mit kommerziellen Kits ohne einfache Reaktion Einrichtung. Bewertungen von Vergleichskosten pro Mikroliter Reaktion zeigen ähnliche Trends (Supplemental Abbildung 1). Wir schätzen die Anlaufkosten zu ~ $4.500 für alle Reagenzien und eine zusätzliche $3.200 für Spezialausrüstung, wie ein Sonikator. Person-Stunden für diesen Vorgang werden voraussichtlich ~ 26 h für alle Reagenz-Vorbereitung vom Boden auf. Jedoch sobald große Vorräte an Reagenzien vorbereitet haben, verringern Anforderungen an die Arbeit erheblich. Darüber, wie die Erfahrung mit der Plattform gewonnen wird, skalieren Sie die Größe der das Zellwachstum empfehlen wir, extrahieren, Vorbereitung und Reagenz Vorbereitung, zeitliche Effizienz zu maximieren. Angesichts der Startkosten, empfehlen wir die GFP-Plattform für Anwendungen in der synthetischen Biologie, Hochdurchsatz-Bemühungen, und Protein-Expression-Bedingungen, die unvereinbar mit traditionellen Protein Ausdruck Plattformen aufgrund zu Konflikten mit der Zelle Biochemie und Lebensfähigkeit Einschränkungen. In diese spezielle Fälle, wo die gewünschte Technik von der GFP-Plattform aktiviert ist, ist die höhere Kosten der GFP über in Vivo Ausdruck gerechtfertigt.
Weiterentwicklung der GFP-Plattform ist wahrscheinlich zu breiteren Dienstprogramm Biotechnologie Bemühungen wie die metabolic Engineering enzymatische Wege, Herstellung und Charakterisierung von traditionell hartnäckigen Proteinen, nicht dem Standard entsprechende Aminosäure Einarbeitung und unnatürliche Protein-Expression, geschichtete Medizin Fertigung und Expansion über das Labor hinaus in die Klassenzimmer für STEM Bildung64,65,66. Diese Anstrengungen werden weiter durch die laufenden Bemühungen zur detaillierten Charakterisierung der GFP-Plattform unterstützt werden. Ein besseres Verständnis der Zusammensetzung der Zelle Extrakt führt zur weiteren Verfeinerung in Richtung verbesserte Reaktion Erträge und Flexibilität bei der Reaktion Bedingungen61,67,68.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Die Autoren erklären, dass sie keine finanziellen Interessenkonflikte oder andere Interessenkonflikte.
Acknowledgments
Autoren möchten Dr. Jennifer VanderKelen, Andrea Laubscher, und Tony Turretto für den technischen Support, Wesley Kao, Layne Williams und Christopher Hight für hilfreiche Diskussionen anerkennen. Autoren erkennen auch finanzielle Unterstützung von Bill und Linda Frost Fonds, Zentrum für Anwendungen in der Biotechnologie Chevron Biotechnologie angewendet Endowment Forschungsstipendium, Cal Poly Forschung Scholarly und kreative Aktivitäten Grant Program (RSCA 2017), und die National Science Foundation (NSF-1708919). MZL erkennt an der California State University Graduate Grant. MCJ der Army Research Office W911NF-16-1-0372 anerkennt, National Science Foundation gewährt, MCB 1413563 MCB-1716766 und der Luftwaffe Forschung Labor Center of Excellence Grant FA8650-15-2-5518, die Verteidigung Bedrohung Reduktion Agentur Grant HDTRA1-15-10052/P00001, der David and Lucile Packard Foundation, die Camille Dreyfus Lehrer-Scholar Program, die Abteilung von Energie BER Grant DE-SC0018249, die menschliche Frontiers Science Program (RGP0015/2017), DOE Joint Genome Institute ETOP Grant, und das Chicago biomedizinische Konsortium mit Unterstützung der Searle-Fonds auf der Chicago Community Trust für Unterstützung.
Materials
| Name | Company | Catalog Number | Comments |
| Luria Broth | ThermoFisher | 12795027 | |
| Tryptone | Fisher Bioreagents | 73049-73-7 | |
| Yeast Extract | Fisher Bioreagents | 1/2/8013 | |
| NaCl | Sigma-Aldrich | S3014-1KG | |
| Potassium Phosphate Dibasic | Sigma-Aldrich | 60353-250G | |
| Potassium Phosphate Monobasic | Sigma-Aldrich | P9791-500G | |
| D-Glucose | Sigma-Aldrich | G8270-1KG | |
| KOH | Sigma-Aldrich | P5958-500G | |
| IPTG | Sigma-Aldrich | I6758-1G | |
| Mg(OAc)2 | Sigma-Aldrich | M5661-250G | |
| K(OAc) | Sigma-Aldrich | P1190-1KG | |
| Tris(OAc) | Sigma-Aldrich | T6066-500G | |
| DTT | ThermoFisher | 15508013 | |
| tRNA | Sigma-Aldrich | 10109541001 | |
| Folinic Acid | Sigma-Aldrich | F7878-100MG | |
| NTPs | ThermoFisher | R0481 | |
| Oxalic Acid | Sigma-Aldrich | P0963-100G | |
| NAD | Sigma-Aldrich | N8535-15VL | |
| CoA | Sigma-Aldrich | C3144-25MG | |
| PEP | Sigma-Aldrich | 860077-250MG | |
| K(Glu) | Sigma-Aldrich | G1501-500G | |
| NH4(Glu) | MP Biomedicals | 02180595.1 | |
| Mg(Glu)2 | Sigma-Aldrich | 49605-250G | |
| Spermidine | Sigma-Aldrich | S0266-5G | |
| Putrescine | Sigma-Aldrich | D13208-25G | |
| HEPES | ThermoFisher | 11344041 | |
| Molecular Grade Water | Sigma-Aldrich | 7732-18-5 | |
| L-Aspartic Acid | Sigma-Aldrich | A7219-100G | |
| L-Valine | Sigma-Aldrich | V0500-25G | |
| L-Tryptophan | Sigma-Aldrich | T0254-25G | |
| L-Phenylalanine | Sigma-Aldrich | P2126-100G | |
| L-Isoleucine | Sigma-Aldrich | I2752-25G | |
| L-Leucine | Sigma-Aldrich | L8000-25G | |
| L-Cysteine | Sigma-Aldrich | C7352-25G | |
| L-Methionine | Sigma-Aldrich | M9625-25G | |
| L-Alanine | Sigma-Aldrich | A7627-100G | |
| L-Arginine | Sigma-Aldrich | A8094-25G | |
| L-Asparagine | Sigma-Aldrich | A0884-25G | |
| Glycine | Sigma-Aldrich | G7126-100G | |
| L-Glutamine | Sigma-Aldrich | G3126-250G | |
| L-Histadine | Sigma-Aldrich | H8000-25G | |
| L-Lysine | Sigma-Aldrich | L5501-25G | |
| L-Proline | Sigma-Aldrich | P0380-100G | |
| L-Serine | Sigma-Aldrich | S4500-100G | |
| L-Threonine | Sigma-Aldrich | T8625-25G | |
| L-Tyrosine | Sigma-Aldrich | T3754-100G | |
| Fisherbrand Premium Microcentrifuge Tubes: 2.0 mL | Fisher Scientific | 05-408-138 | |
| Fisherbrand Premium Microcentrifuge Tubes: 1.5 mL | Fisher Scientific | 05-408-129 | |
| Fisherbrand Premium Microcentrifuge Tubes: 0.6 mL | Fisher Scientific | 05-408-120 | |
| PureLink HiPure Plasmid Prep Kit | ThermoFisher | K210007 | |
| Ultrasonic Processor | QSonica | Q125-230V/50Hz | 3.175 mm diameter probe |
| Avanti J-E Centrifuge | Beckman Coulter | 369001 | |
| JLA-8.1000 Rotor | Beckman Coulter | 366754 | |
| 1L Centrifuge Tube | Beckman Coulter | A99028 | |
| Tunair 2.5L Baffeled Shake Flask | Sigma-Aldrich | Z710822 | |
| Microfuge 20 | Beckman Coulter | B30134 | |
| New Brunswick Innova 42/42R Incubator | Eppendorf | M1335-0000 | |
| Cytation 5 | BioTek | ||
| Strep-Tactin XT Starter Kit | IBA | 2-4998-000 | |
| pJL1-sfGFP | Addgene | 69496 | |
| BL21(DE3) | New England BioLabs |
References
- Jiang, L., Zhao, J., Lian, J., Xu, Z. Cell-free protein synthesis enabled rapid prototyping for metabolic engineering and synthetic biology. Synthetic and Systems Biotechnology. 3 (2), 90-96 (2018).
- Carlson, E. D., Gan, R., Hodgman, C. E., Jewett, M. C. Cell-free protein synthesis: Applications come of age. Biotechnology Advances. 30 (5), 1185-1194 (2012).
- Watanabe, M. Cell-Free Protein Synthesis for Structure Determination by X-ray Crystallography. Methods in molecular biology (Clifton, N.J). 607, 149-160 (2010).
- Martemyanov, K. A., Shirokov, V. A., Kurnasov, O. V., Gudkov, A. T., Spirin, A. S. Cell-Free Production of Biologically Active Polypeptides: Application to the Synthesis of Antibacterial Peptide Cecropin. Protein Expression and Purification. 21 (3), 456-461 (2001).
- Renesto, P., Raoult, D. From genes to proteins: in vitro expression of rickettsial proteins. Annals of the New York Academy of Sciences. 990, 642-652 (2003).
- Xu, Z., Chen, H., Yin, X., Xu, N., Cen, P. High-Level Expression of Soluble Human b-Defensin-2 Fused With Green Fluorescent Protein in Escherichia coli Cell-Free System. Applied Biochemistry and Biotechnology. 127 (1), 053-062 (2005).
- Baumann, A. In-situ observation of membrane protein folding during cell-free expression. PLoS ONE. 11 (3), 1-15 (2016).
- Wang, Y., Percival, Y. H. P. Cell-free protein synthesis energized by slowly-metabolized maltodextrin. BMC Biotechnology. 9, 1-8 (2009).
- Whittaker, J. W. Cell-free protein synthesis: the state of the art. Biotechnology Letters. 35 (2), 143-152 (2013).
- Martin, R. W. Cell-free protein synthesis from genomically recoded bacteria enables multisite incorporation of noncanonical amino acids. Nature Communications. 9 (1), 1203 (2018).
- Kwon, Y. C., Song, J. K., Kim, D. M. Cloning-Independent Expression and Screening of Enzymes Using Cell-Free Protein Synthesis Systems. Methods in Molecular Biology. 1118, 97-108 (2014).
- Chappell, J., Jensen, K., Freemont, P. S. Validation of an entirely in vitro approach for rapid prototyping of DNA regulatory elements for synthetic biology. Nucleic Acids Research. 41 (5), 3471-3481 (2013).
- Takahashi, M. K. Characterizing and prototyping genetic networks with cell-free transcription-translation reactions. Methods. 86, 60-72 (2015).
- Karim, A. S., Jewett, M. C. A cell-free framework for rapid biosynthetic pathway prototyping and enzyme discovery. Metabolic Engineering. 36, 116-126 (2016).
- Dudley, Q. M., Anderson, K. C., Jewett, M. C. Cell-Free Mixing of Escherichia coli Crude Extracts to Prototype and Rationally Engineer High-Titer Mevalonate Synthesis. ACS Synthetic Biology. 5 (12), 1578-1588 (2016).
- Pardee, K. Paper-based synthetic gene networks. Cell. 159 (4), 940-954 (2014).
- Zawada, J. F. Microscale to manufacturing scale-up of cell-free cytokine production-a new approach for shortening protein production development timelines. Biotechnology and Bioengineering. 108 (7), 1570-1578 (2011).
- Sullivan, C. J. A cell-free expression and purification process for rapid production of protein biologics. Biotechnology Journal. 11 (2), 238-248 (2016).
- Li, J. Cell-free protein synthesis enables high yielding synthesis of an active multicopper oxidase. Biotechnology Journal. 11 (2), 212-218 (2016).
- Heinzelman, P., Schoborg, J. A., Jewett, M. C. pH responsive granulocyte colony-stimulating factor variants with implications for treating Alzheimer's disease and other central nervous system disorders. Protein Engineering Design and Selection. 28 (10), 481-489 (2015).
- Pardee, K. Low-Cost Detection of Zika Virus Using Programmable Biomolecular Components. Cell. 165 (5), 1255-1266 (2016).
- Slomovic, S., Pardee, K., Collins, J. J. Synthetic biology devices for in vitro and in vivo diagnostics. Proceedings of the National Academy of Sciences. 112 (47), 14429-14435 (2015).
- Gootenberg, J. S. Nucleic acid detection with CRISPR-Cas13a/C2c2. Science. 356 (6336), 438-442 (2017).
- Pardee, K. Portable, On-Demand Biomolecular Manufacturing. Cell. 167 (1), 248-259 (2016).
- Karig, D. K., Bessling, S., Thielen, P., Zhang, S., Wolfe, J. Preservation of protein expression systems at elevated temperatures for portable therapeutic production. Journal of the Royal Society, Interface. 14 (129), (2017).
- Smith, M. T., Berkheimer, S. D., Werner, C. J., Bundy, B. C. Lyophilized Escherichia coli-based cell-free systems for robust, high-density, long-term storage. BioTechniques. 56 (4), 186-193 (2014).
- Hunt, J. P., Yang, S. O., Wilding, K. M., Bundy, B. C. The growing impact of lyophilized cell-free protein expression systems. Bioengineered. 8 (4), 325-330 (2017).
- Stark, J. C. BioBitsTM Bright: A fluorescent synthetic biology education kit. Science Advances. 4 (8), (2018).
- Huang, A. BioBitsTM Explorer: A modular synthetic biology education kit. Science Advances. 4 (8), (2018).
- Zemella, A., Thoring, L., Hoffmeister, C., Kubick, S. Cell-Free Protein Synthesis: Pros and Cons of Prokaryotic and Eukaryotic Systems. ChemBioChem. 16 (17), 2420-2431 (2015).
- Oza, J. P. Robust production of recombinant phosphoproteins using cell-free protein synthesis. Nature Communications. 6 (1), 8168 (2015).
- Zemella, A. Cell-free protein synthesis as a novel tool for directed glycoengineering of active erythropoietin. Scientific Reports. 8 (1), 8514 (2018).
- Jaroentomeechai, T. Single-pot glycoprotein biosynthesis using a cell-free transcription-translation system enriched with glycosylation machinery. Nature Communications. 9 (1), 2686 (2018).
- Kightlinger, W. Design of glycosylation sites by rapid synthesis and analysis of glycosyltransferases. Nature Chemical Biology. 14 (6), 627-635 (2018).
- Schoborg, J. A. A cell-free platform for rapid synthesis and testing of active oligosaccharyltransferases. Biotechnology and Bioengineering. 115 (3), 739-750 (2018).
- Chekulayeva, M. N., Kurnasov, O. V., Shirokov, V. A., Spirin, A. S. Continuous-Exchange Cell-Free Protein-Synthesizing System: Synthesis of HIV-1 Antigen Nef. Biochemical and Biophysical Research Communications. 280 (3), 914-917 (2001).
- Hong, S. H. Improving Cell-Free Protein Synthesis through Genome Engineering of Escherichia coli Lacking Release Factor 1. ChemBioChem. 16 (5), 844-853 (2015).
- Endo, Y., Otsuzuki, S., Ito, K., Miura, K. Production of an enzymatic active protein using a continuous flow cell-free translation system. Journal of Biotechnology. 25 (3), 221-230 (1992).
- Volyanik, E. V., Dalley, A., Mckay, I. A., Leigh, I., Williams, N. S., Bustin, S. A. Synthesis of Preparative Amounts of Biologically Active Interleukin-6 Using a Continuous-Flow Cell-Free Translation System. Analytical Biochemistry. 214 (1), 289-294 (1993).
- Martin, G. A., Kawaguchi, R., Lam, Y., DeGiovanni, A., Fukushima, M., Mutter, W. High-yield, in vitro protein expression using a continuous-exchange, coupled transcription/ translation system. BioTechniques. 31 (4), 948-950 (2001).
- Stech, M., Quast, R. B., Sachse, R., Schulze, C., Wüstenhagen, D. A., Kubick, S. A Continuous-Exchange Cell-Free Protein Synthesis System Based on Extracts from Cultured Insect Cells. PLoS ONE. 9 (5), e96635 (2014).
- Quast, R. B., Sonnabend, A., Stech, M., Wüstenhagen, D. A., Kubick, S. High-yield cell-free synthesis of human EGFR by IRES-mediated protein translation in a continuous exchange cell-free reaction format. Scientific Reports. 6 (1), 30399 (2016).
- Thoring, L., Dondapati, S. K., Stech, M., Wüstenhagen, D. A., Kubick, S. High-yield production of "difficult-to-express" proteins in a continuous exchange cell-free system based on CHO cell lysates. Scientific Reports. 7 (1), 11710 (2017).
- Hoffmann, M., Nemetz, C., Madin, K., Buchberger, B. Rapid translation system: A novel cell-free way from gene to protein. Biotechnology annual review. 10, 1-30 (2004).
- Kwon, Y. C., Jewett, M. C. High-throughput preparation methods of crude extract for robust cell-free protein synthesis. Scientific Reports. 5 (1), 8663 (2015).
- Katsura, K. A reproducible and scalable procedure for preparing bacterial extracts for cell-free protein synthesis. Journal of Biochemistry. 162 (June), 357-369 (2017).
- Fujiwara, K., Doi, N. Biochemical preparation of cell extract for cell-free protein synthesis without physical disruption. PLoS ONE. 11 (4), 1-15 (2016).
- Shrestha, P., Holland, T. M., Bundy, B. C. Streamlined extract preparation for Escherichia coli-based cell-free protein synthesis by sonication or bead vortex mixing. BioTechniques. 53 (3), 163-174 (2012).
- Krinsky, N. A Simple and Rapid Method for Preparing a Cell-Free Bacterial Lysate for Protein Synthesis. PLoS ONE. 11 (10), (2016).
- Jewett, M. C., Swartz, J. R. Mimicking the Escherichia coli cytoplasmic environment activates long-lived and efficient cell-free protein synthesis. Biotechnology and Bioengineering. 86 (1), 19-26 (2004).
- Swartz, J. R., Jewett, M. C., Woodrow, K. A. Cell-Free Protein Synthesis With Prokaryotic Combined Transcription-Translation. Recombinant Gene Expression. (267), 169-182 (2004).
- Voloshin, A. M., Swartz, J. R. Efficient and scalable method for scaling up cell free protein synthesis in batch mode. Biotechnology and Bioengineering. 91 (4), 516-521 (2005).
- Vernon, W. B. The role of magnesium in nucleic-acid and protein metabolism. Magnesium. 7 (5-6), 234-248 (1988).
- Pratt, J. M. Transcription and Translation: A Practical Approach. , IRL Press. New York. (1984).
- Kim, D. M., Kigawa, T., Choi, C. Y., Yokoyama, S. A Highly Efficient Cell-Free Protein Synthesis System from Escherichia coli. European Journal of Biochemistry. 239 (3), 881-886 (1996).
- Shin, J., Noireaux, V. Efficient cell-free expression with the endogenous E. Coli RNA polymerase and sigma factor 70. Journal of Biological Engineering. 4 (1), 8 (2010).
- Zubay, G. In Vitro Synthesis of Protein in Microbial Systems. Annual Review of Genetics. 7 (1), 267-287 (1973).
- Kigawa, T. Preparation of Escherichia coli cell extract for highly productive cell-free protein expression. Journal of Structural and Functional Genomics. 5 (1/2), 63-68 (2004).
- Liu, D. V., Zawada, J. F., Swartz, J. R. Streamlining Escherichia Coli S30 Extract Preparation for Economical Cell-Free Protein Synthesis. Biotechnology Progress. 21 (2), 460-465 (2008).
- Yang, W. C., Patel, K. G., Wong, H. E., Swartz, J. R. Simplifying and streamlining Escherichia coli-based cell-free protein synthesis. Biotechnology Progress. 28 (2), 413-420 (2012).
- Foshag, D. The E. coli S30 lysate proteome: A prototype for cell-free protein production. New Biotechnology. 40 (Pt B), 245-260 (2018).
- Caschera, F. Bacterial cell-free expression technology to in vitro systems engineering and optimization. Synthetic and Systems Biotechnology. 2 (2), 97-104 (2017).
- Chizzolini, F., Forlin, M., Yeh Martín, N., Berloffa, G., Cecchi, D., Mansy, S. S. Cell-Free Translation Is More Variable than Transcription. ACS Synthetic Biology. 6 (4), 638-647 (2017).
- Hong, S. H., Kwon, Y. C., Jewett, M. C. Non-standard amino acid incorporation into proteins using Escherichia coli cell-free protein synthesis. Frontiers in Chemistry. 2, 34 (2014).
- Sawasaki, T., Ogasawara, T., Morishita, R., Endo, Y. A cell-free protein synthesis system for high-throughput proteomics. Proceedings of the National Academy of Sciences of the United States of America. 99 (23), 14652-14657 (2002).
- Dudley, Q. M., Karim, A. S., Jewett, M. C. Cell-free metabolic engineering: Biomanufacturing beyond the cell. Biotechnology Journal. 10 (1), 69-82 (2015).
- Garcia, D. C. Elucidating the potential of crude cell extracts for producing pyruvate from glucose. Synthetic Biology. 3 (1), 1-9 (2018).
- Hurst, G. B. Proteomics-Based Tools for Evaluation of Cell-Free Protein Synthesis. Analytical Chemistry. 89 (21), 11443-11451 (2017).

{kind=link}
{kind=link}
{kind=link}