

Summary
Ce protocole détaille les étapes, les coûts et équipements nécessaires pour générer des e. coli-extraits cellulaires de base et mettre en œuvre en vitro réactions de synthèse de protéines dans les 4 jours ou moins. Pour tirer parti de la souplesse de cette plate-forme pour les applications larges, nous discutons des conditions de réaction qui peuvent être adaptées et optimisées.
Abstract
Au cours des 50 dernières années, la synthèse de protéine acellulaire (CFPS) est devenue une technologie puissante pour exploiter la capacité transcriptionnelle et translationnelle des cellules dans un tube à essai. En évitant la nécessité de maintenir la viabilité de la cellule et en éliminant la barrière cellulaire, PFC a été fondamentale aux applications émergentes en biofabrication de protéines traditionnellement difficiles, ainsi que des applications de prototypage rapide pour génie métabolique et la génomique fonctionnelle. Nos méthodes de mise en œuvre d’un e. coli-plateforme PFC permettre à nouveau aux utilisateurs d’accéder à bon nombre de ces applications. Nous décrivons ici les méthodes pour préparer l’extrait par l’utilisation de milieux enrichis, flacons déroutés et une méthode reproductible de la lyse cellulaire de base sonication accordable. Cet extrait peut ensuite servir pour l’expression de la protéine capable de produire 900 µg/mL ou plus de la protéine fluorescente de super dossier vert (sfGFP) dans seulement 5 h de montage expérimental à l’analyse des données, étant donné que les stocks de réactifs appropriés ont été préparés préalablement. Le coût estimatif de démarrage d’obtenir des réactifs est $ 4 500 qui soutiendront des milliers de réactions à un coût estimatif de 0,021 $ par µg de protéines produites ou 0,019 $ par µL de réaction. En outre, les méthodes d’expression de protéine en miroir la facilité de l’installation de réaction vue dans les systèmes disponibles dans le commerce grâce à l’optimisation des mélanges réactifs, à une fraction du coût. Afin de permettre à l’utilisateur de tirer parti de la souplesse de la plate-forme de produits forestiers certifiés pour des applications larges, nous avons identifié les divers aspects de la plate-forme qui peut être à l’écoute et optimisé selon les ressources disponibles et des résultats d’expression de protéine vous le souhaitez.
Introduction
Synthèse de protéine acellulaire (CFPS) a émergé comme une technologie qui a débloqué un certain nombre de nouvelles opportunités pour la production de protéines, génomique fonctionnelle, génie métabolique et plus encore dans les derniers 50 ans1,2. Par rapport à la norme en vivo plates-formes expression de protéine, CFPS offre trois avantages principaux : 1) la nature acellulaire de la plate-forme permet la production de protéines qui seraient potentiellement toxiques ou étrangers à la cellule3,4 ,5,6; 2) inactivation de l’ADN génomique et la mise en place d’un modèle d’ADN codant le gène d’intérêt canaliser toute l’énergie systémique au sein de la réaction à la production des ou des protéines d’intérêt ; et 3) la nature ouverte de la plate-forme permet à l’utilisateur de modifier et de surveiller les conditions de la réaction et la composition en temps réel7,8. Cet accès direct à la réaction prend en charge l’augmentation des systèmes biologiques avec des chimies élargis et conditions redox pour la production de nouvelles protéines et l’accordage des processus métaboliques2,9, 10. direct access permet également l’utilisateur de combiner la réaction de PFC avec tests d’activité dans un système single-pot des cycles de conception-construction-test plus rapide11. La capacité à effectuer la réaction de PFC dans les gouttelettes de faible volume ou sur appareils sur papier plus soutient les efforts de découverte de haut débit et de prototypage rapide12,13,14,15 ,,16. À la suite de ces avantages et de la nature de plug-and-play du système, CFPS unique a permis à une variété d’applications de biotechnologie telles que la production de protéines qui sont difficilement soluble express en vivo17, 18,19,20, détection des maladies21,22,23, sur demande la biofabrication18,24 ,25,26,27et l’éducation28,29, qui montrent la flexibilité et l’utilité de la plate-forme acellulaire.
Systèmes de PFC peuvent provenir d’une variété de lysats bruts de deux lignées de cellules procaryotes et eucaryotes. Cela permet diverses options dans le système de choix, chacun d'entre eux ont des avantages et inconvénients selon l’application des intérêts. Systèmes de CFPS varient également considérablement en productivité, coût et temps de préparation. Le plus couramment utilisé cellule extraits sont produites à partir de germe de blé, réticulocytes de lapin, des cellules d’insecte et cellules d’Escherichia coli , cette dernière étant la plus efficace à ce jour tout en produisant le meilleur rendement volumétrique de la protéine30 . Alors que les autres systèmes de PFC peuvent être avantageux pour leurs machines de modification post-traductionnelle innée, nouvelles applications à l’aide de l' e. coli-machines basé sont en mesure de combler le fossé en générant relativement phosphorylée et protéines glycosylées sur demande31,32,33,34,35.
Réactions de PFC peuvent être exécutées dans chaque lot, échange continu acellulaire (CECF) ou acellulaire (CFCF) formats de flux continu. Le format de lot est un système fermé, dont vie de réaction est limitée en raison de la diminution des quantités de réactifs et l’accumulation des sous-produits inhibiteurs de la réaction. Méthodes CECF et CFCF augmentent la durée de vie de la réaction et ainsi entraînent des rendements de protéine volumétrique accrue par rapport à la réaction de lot. Ceci est accompli en autorisant les sous-produits de la synthèse protéique à être retiré de la cuve de réaction alors que nouveaux réactifs sont fournis tout au long de la réaction2. Dans le cas de la CFCF, la protéine d’intérêt peut également être retirée de la chambre de réaction, tandis que dans la CECF, la protéine d’intérêt demeure dans la chambre de réaction, composé d’une membrane semi-perméable36,37. Ces méthodes sont particulièrement utiles pour surmonter les piètres rendements volumétriques de difficile-à-express des protéines d’intérêt38,39,40,41,42, 43. Les difficultés à mettre en œuvre les approches CECF et CFCF sont que 1) alors qu’ils se traduire par une utilisation plus efficace des machines bio responsable de la transcription et la traduction, ils exigent notamment de plus grandes quantités de réactifs, ce qui augmente le coût global et 2) ils nécessitent des configurations plus complexes de réaction et d’équipements spécialisés par rapport à la lot format44. Afin d’assurer l’accessibilité pour les nouveaux utilisateurs, les protocoles décrits se concentrer sur le format de traitement par lots à des volumes de réaction de 15 µL avec des recommandations précises pour augmenter le volume de réaction à l’échelle de millilitre.
Les méthodes présentées ici permettent profanes avec des compétences de laboratoire de base (telles que les étudiants de premier cycle) à mettre en œuvre la croissance cellulaire, extrait préparation et format réaction configuration de lot pour un e. coli-système de PFC. Cette approche est rentable par rapport aux kits disponibles dans le commerce sans sacrifier la facilité d’installation basé sur kit de réaction. En outre, cette approche permet aux applications en laboratoire et sur le terrain. Lorsque vous décidez de mettre en œuvre des produits forestiers certifiés, nouveaux utilisateurs devraient évaluer soigneusement l’efficacité des systèmes d’expression de protéine conventionnelle pour l’investissement de démarrage, comme le PFC n’est peut-être pas supérieure dans tous les cas. Les méthodes CFPS décrites ici permettent à l’utilisateur d’appliquer directement une variété d’applications, y compris la génomique fonctionnelle, haut débit stable, la production de protéines qui sont insolubles pour in vivo de l’expression, mais aussi de champ applications, y compris les biocapteurs et trousses pédagogiques pour la biologie synthétique. Des applications supplémentaires, tels que l’ingénierie métabolique, tuning des conditions d’expression de protéine, détection de la maladie et mesurent-vers le haut à l’aide des méthodes CECF ou CFCF sont toujours possibles mais peuvent exiger l’expérience avec la plateforme PFC pour une modification supplémentaire de la réaction conditions. Nos méthodes de conjuguer croissance en milieux enrichis et flacons déroutés, et des méthodes relativement rapides et reproductibles de la lyse cellulaire par le biais de la sonication, suivie d’une configuration de réaction CFPS simplifiée qui utilise des prémélanges optimisé45. Même si les méthodes de croissance cellulaire ont devenu quelque peu normalisées dans ce domaine, procédés de lyse cellulaire varient considérablement. En plus de la sonication, les méthodes de lyse courantes sont utilisation d’une presse Français, un homogénéisateur, batteurs de perle, ou lysozyme et autres perturbations biochimiques et physiques méthodes46,47,48, 49. à l’aide de nos méthodes, environ 2 mL d’extrait cellulaire brut sont obtenus par 1 L de cellules. Cette quantité d’extrait cellulaire peut prendre en charge quatre cents réactions de CFPS 15 µL, chaque producteur ~ 900 µg/mL de protéine sfGFP journaliste du plasmide de modèle pJL1-sfGFP. Cette méthode coûte $ 0,021/µg de sfGFP produit ($.019/µL de réaction), à l’exclusion du coût du travail et des équipements (la Figure 1). À partir de l’éraflure, cette méthode peut être implémentée en 4 jours par une seule personne et répétez les réactions PFC peuvent être complétées en heures (Figure 1). En outre, le protocole peut être transposé en volume pour les plus gros lots de préparation du réactif aux besoins de l’utilisateur. Ce qui est important, le protocole présenté ici peut être implémenté par laboratoire formé profanes tels que les étudiants de premier cycle, comme elle exige seulement des compétences du laboratoire de base. Les procédures décrites ci-dessous et la vidéo qui l’accompagne ont été spécifiquement développés pour améliorer l’accessibilité de la plate-forme de PFC d’e. coli pour une utilisation large.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. médias préparation et croissance cellulaire
-
Jour 1
- Cellules d’Escherichia coli BL21*(DE3) Streak d’un glycérol stock sur une gélose LB et incuber pendant au moins 18 h à 37 ° C.
- Préparer 50 mL de médias LB et stériliser la solution sur un cycle de liquid pendant 30 minutes à 121 ° C. Conserver à température ambiante.
-
Jour 2
- Préparer 750 mL de 2 x YTP médias et 250 mL de 0,4 M de D-Glucose solution comme indiqué dans les informations supplémentaires.
- Versez les 2 médias de la x YTP dans une fiole dérouté stérilisés à l’autoclave de 2,5 L et la solution de D-Glucose dans une bouteille en verre stérilisés à l’autoclave de 500 mL. Autoclave les deux solutions selon un cycle de liquide pendant 30 minutes à 121 ° C.
- Veiller à ce que les deux solutions stériles sont stockées à 37 ° C si la croissance cellulaire est effectuée le jour suivant, afin de maximiser les taux de croissance après l’inoculation. Ne pas combiner des solutions avant l’inoculation.
Remarque : Les Solutions peuvent être stockées à 4 ° C pendant 1-2 d si nécessaire, bien que les médias x YTP 2 est très sujette à la contamination. - Démarrer une culture nuitée de BL21 (de3) en inoculant 50 mL de médias LB avec une seule colonie de BL21 (de3) à l’aide d’une boucle stérilisé et stérile pour éviter toute contamination.
- Placer les 50 mL de la culture BL21*(DE3) LB dans un 37 ° C 250 tr/min, secouant incubateur et croître durant la nuit pendant 15 à 18 h.
- Préparer et stériliser tout le matériel nécessaire pour les jours 3 et 4, y compris : deux bouteilles de centrifugeuse 1 L, 4 tubes de coniques de froid 50 mL x (poids et masses records de trois) et nombreux microtubes de 1,5 mL.
-
Jour 3
- Retirer la culture nuit 50 mL de BL21*(DE3) dans des livres de l’incubateur agitateur et mesurer l' OD600 sur un spectrophotomètre à l’aide d’un 01:10 dilution avec LB media. Calculer le volume d’une culture nécessaire d’ajouter à 1 L de médias pour un départ de l’OD600 de 0,1 (par exemple, si un OD600 d’un 01:10 dilution est lu comme 0,4, inoculer 25 mL de la non dilué de l’OD600 = 4,0 culture nuit dans 1 L de 2 x YTP G).
- Sortir les chauffée 2 x YTP les médias et les solutions de D-Glucose de l’incubateur de 37 ° C ainsi que les 50 mL de culture LB. Utilisant une technique stérile, verser délicatement la solution de D-Glucose dans les médias x YTP 2 (en évitant les côtés du ballon dérouté).
Remarque : L’Addition de D-Glucose termine la recette pour 1 L de 2 x YTPG. - Maintien technique stérile, ensemencer le 1 L de solution de 2 x YTPG avec la quantité appropriée de la culture de 50 mL pour commencer la culture de 1 L à un 0.1 OD600. Placer immédiatement la culture 1 L inoculés dans un 37 ° C, agitation incubateur à 200 tr/min.
- Prendre la première OD600 lecture après la première heure de croissance (phase de latence typique prend 1 h). Ne pas diluer la culture. Continuer à prendre des mesures OD600 environ toutes les 20-30 min OD600 jusqu'à 0,6.
- En arrivant à OD600 = 0,6, ajouter 1 mL de 1 M IPTG (concentration finale dans la culture de 1 L = 1 mM) à la culture de x YTPG 2.
NOTE : Induction idéal OD600 est 0,6 ; Toutefois, une fourchette de 0,6 à 0,8 est acceptable. L’induction de l’IPTG est pour la production endogène de T7 RNA polymérase (T7RNAP). - Après l’induction, mesurer l' OD600 environ toutes les 20-30 min jusqu'à ce qu’il atteigne 3.0.
Remarque : Le refroidissement le centrifuger à 4 ° C pendant cette période. Préparer le tampon S30 froid comme il est indiqué dans les informations complémentaires. Si la mémoire tampon S30 est préparée à l’avance, veiller à ce que la TNT n’est pas ajoutée jusqu’au jour d’utilisation. - Une fois que l' OD600 atteint 3,0 (Figure 2 a), versez la culture dans une bouteille de centrifugeuse 1 L froid dans un bain d’eau glacée. Préparer une bouteille de centrifugeuse 1 L rempli d’eau d’un poids égal à utiliser comme un équilibre dans la centrifugeuse.
NOTE : Valeurs d’Absorbance varient de l’instrument-à-instrument. Alors que l' OD600 de récolte de BL21 (de3) n’est pas une variable sensible, il est recommandé que l’utilisateur évaluer et optimiser cette variable mesure de dépannage. Spectrophotomètres plus susceptible lectures de600 OD relativement faibles par rapport aux plus petits spectrophotomètres axée sur la cuvette. - Centrifuger les bouteilles de 1 L pendant 10 min à 5 000 x g et 10 ° C à granules des cellules.
- Lentement de décanter le liquide surnageant et la jeter conformément aux procédures des déchets biologiques de l’institution. Placez la pastille sur la glace.
- À l’aide d’une spatule stérile, gratter le culot de la bouteille de la centrifugeuse et le transférer dans un tube conique froid 50 mL.
- Ajouter 30 mL de tampon de S30 froid dans le tube conique et resuspendre le culot cellulaire au vortex avec saccades (20-30 s) et de périodes de repos (1 min) sur la glace jusqu'à ce que complètement remises en suspension avec aucun morceaux.
- Une fois que le culot est resuspendu pleinement, utiliser un autre tube conique de 50 mL avec de l’eau comme un équilibre et une centrifugation pendant 10 min à 5000 x g et 10 ° C (préalablement refroidi à 4 ° C).
Remarque : Ceci termine le 1st 3 lavages nécessaires lorsqu’ils pêchent les cellules. - Versez le liquide surnageant et la jeter conformément aux procédures des déchets biologiques de l’institution. Resuspendre le culot avec 20-25 mL de froid S30 mémoire tampon et centrifuger pendant 10 min à 5 000 x g et 10 ° C (préalablement refroidi à 4 ° C).
Remarque : Ceci termine 2ème de 3 lavages. - Encore une fois, versez le liquide surnageant et la jeter conformément aux procédures des déchets biologiques de l’institution. Ajouter exactement 30 mL de tampon de S30 et vortex à nouveau à resuspendre le culot.
- En utilisant les 3 tubes coniques pré-pesés, froid 50 mL et une pipette sérologique avec une pipette stérile, aliquote 10 mL du mélange tampon pellet/S30 resuspendues dans chacun des 3 tubes coniques.
NOTE : Fractionner les cellules en 3 tubes n’est pas nécessaire, mais cette étape se traduit par petites boulettes de cellule (~ 1 g) pour une commodité accrue à des étapes ultérieures. - Centrifuger tous les tubes, à l’aide de balances appropriées selon les besoins, pendant 10 min à 5000 x g et 10 ° C (préalablement refroidi à 4 ° C).
Remarque : Ceci termine l’étape de lavage final. - Versez le liquide surnageant et la jeter conformément aux procédures des déchets biologiques de l’institution. Enlever l’excès de tampon S30 en essuyant soigneusement l’intérieur du tube à fond conique et boucher avec un tissu propre ; Évitez de toucher le culot.
- Réévaluer les tubes sur une balance analytique et noter le poids de granule final sur chaque tube.
Remarque : Le protocole peut être suspendu à ce stade. Les pellets peuvent être flash congelés dans l’azote liquide et stockées à-80 ° C pendant un an jusqu’au moment de la préparation de l’extrait.
2. préparation d’extrait cellulaire brut - jour 4
- Pour la préparation de l’extrait, garder les cellules froid sur la glace au cours de chaque étape. Ajouter 1 mL de tampon de S30 froid par 1 g de masse cellulaire de la pastille. Veiller à ce que le dithiothréitol (DTT) a été complété au tampon S30 à une concentration finale de 2 mM.
Remarque : Le refroidissement la microcentrifugeuse à 4 ° C pendant cette période. - Resuspendre le culot cellulaire au vortex avec saccades (20-30 s) et de périodes de repos (1 min) sur la glace jusqu'à ce que complètement remises en suspension. Si remise en suspension est difficile, laisser les boulettes sur la glace pendant 30 min décongeler.
- Transfert de 1,4 mL de cellules resuspendues dans un tube de microtubes de 1,5 mL.
- Placer un tube de 1,5 mL contenant 1,4 mL de cellules resuspendues dans un bain d’eau glacée dans un bécher. Laisser agir pendant 45 s sur suivie de 59 s éteint pendant 3 cycles totales, avec une amplitude fixée à 50 %. Fermer et retourner les tubes à mélanger doucement pendant les périodes d’arrêt. Au total, livrer 800-900 J d’énergie dans chaque tube de microtubes de 1,5 mL contenant 1,4 mL de cellules resuspendues (Figure 3 a et 3 b).
Remarque : Cette étape est sensible au type sonicateur et le modèle utilisé et doit être optimisé si le matériel est différent de celui indiqué pour cette procédure. Deux approches complémentaires peuvent être utilisés pour intensifier la quantité d’extrait préparé au cours de cette étape : 1) multiples microtubes de 1,5 mL peuvent être sonication en parallèle, et/ou 2) plus gros volumes peuvent être sonication en tubes coniques (jusqu'à 15 mL de cellule remise en suspension par tube) , mise à l’échelle de la quantité d’énergie fournie comme décrit précédemment, 29,45. - Immédiatement après que sonication est terminée, ajouter 4,5 µL de 1 M DTT (complétant un TNT supplémentaires de 2 mM) dans le 1,4 mL de lysat et inverser plusieurs fois pour mélanger. Placer le tube sur la glace. Répétez les étapes 2.4 et 2.5 pour les tubes supplémentaires de cellules resuspendues avant de procéder à une centrifugation.
- Microcentrifugeuse échantillons à 18 000 x g et 4 ° C pendant 10 min (Figure 3).
- Pipeter le surnageant dans un nouveau tube de microtubes de 1,5 mL. Ne pas déranger le culot ; Il est préférable de laisser certaines surnageant pour maintenir la pureté qu’afin de perturber le culot dans les efforts pour maximiser le rendement.
- Incuber le surnageant de l’étape précédente à 250 tr/min et 37 ° C pendant 60 min par du ruban adhésif les tubes sur la plateforme vibrante de l’incubateur (c’est la réaction de ruissellement).
- Microcentrifugeuse échantillons à 10 000 x g et 4 ° C pendant 10 min.
- Retirez le surnageant sans déranger le culot et le transférer dans un nouveau tube. Créer plusieurs 100 µL d’extraits d’extrait pour le stockage.
Remarque : Le protocole peut être suspendu ici, et l’extrait peut être flash congelés dans l’azote liquide et stockées à-80 ° C pendant jusqu'à une année jusqu'à ce que nécessaire pour les réactions de la PFC. Au moins 5 cycles de gel-dégel peuvent subir sans que cela nuise pour extraire la productivité (Figure 4).
3. synthèse des proteines acellulaires lot Format réactions
- Dégel Solutions une et B ADN modèle, BL21 (de3) extrait (si surgelés), T7RNAP et une partie aliquote d’eau moléculaire de catégorie.
Remarque : Modèle de réaction CFPS se trouvent dans les Informations complémentaires. Recettes des solutions A et B sont fournis dans les Informations complémentaires et correspondent à des concentrations spécifiques pour nombreux réactifs soutenir le système d’énergie de PANOx-SP basé de PFC. Le rôle de chaque réactif et la variation acceptable dans ces concentrations de réactif qui peut prendre en charge des produits forestiers certifiés ont été déterminés50. Un protocole de purification T7RNAP se trouvent dans les informations complémentaires51. T7RNAP supplémentaire peut augmenter le rendement volumétrique mais n’est pas nécessaire si la T7RNAP est induite au cours de la croissance cellulaire. Matrice d’ADN de plasmide (pJL1-sfGFP) peut être préparé à l’aide d’un kit maxiprep avec deux lavages à l’aide du tampon de lavage dans le kit, suivi d’un post-traitement ADN-nettoyage à l’aide d’un kit de purification de PCR (Figure 2 b). Matrices d’ADN linéaires utilisable également dans les réactions de la PFC. - L’étiquette la quantité nécessaire de microtubes nécessaire pour les réactions de la PFC.
Remarque : Les réactions peuvent être réalisées dans différentes tailles de bateau, mais un petit vaisseau peut diminuer les rendements volumétrique de protéine (Figure 2). Amplifier une réaction dans le même récipient de taille peut-être également réduire les rendements volumétriques, en fonction de la diminution de l’échange d’oxygène, due à une diminution du volume rapport surface. Lors de l’augmentation de volume de réaction au-dessus de 100 μL, il est recommandé d’utiliser le fond plat plats 31,37,52. - Ajouter 2,2 µL de la Solution A, 2,1 µL de Solution B, 5 µL de BL21*(DE3) extrait, 0,24 μg de T7RNAP (concentration finale 16 μg/mL), 0,24 ng de la matrice d’ADN (concentration finale de 16 ng/mL) et eau pour porter le volume final à 15 µL.
NOTE : Vortex Solutions A et B fréquemment lors de l’installation de réaction pour éviter la sédimentation des composants et faire en sorte que chaque réaction reçoit une homogène aliquote de chaque solution. Éviter l’agitation l’extrait, plutôt inverser le tube pour mélanger. - Après que tous les réactifs ont été ajoutées à la réaction, mélanger chaque tube en pipette verticalement ou doucement l’agitation tout en veillant à ce que le mélange réactionnel final est combiné en un seul cordon 15 µL au bas du tube de microtubes de 1,5 mL.
- Placer chaque réaction dans l’incubateur à 37 ° C sans agiter pendant 4 h ou 30 ° C durant la nuit.
NOTE : Réactions réussies peuvent être qualitativement évaluées visuellement basé sur la couleur verte du produit sfGFP dans le mélange réactionnel de PFC (Figure 3D). Expression de la protéine d’intérêt peut également être confirmée par SDS-PAGE (la Figure 2).
4. dosage de la protéine de journaliste, [sfGFP]
- Charger 48 µL de 0,05 M HEPES, pH 8, dans chaque bien nécessaire pour la quantification (effectuée en général en trois exemplaires par tube de réaction).
- Supprimer les réactions de l’incubateur. Pipette de haut en bas pour mélanger chaque réaction, puis transférer 2 µL de réaction dans les 48 µL de 0,05 M HEPES, pH 8. Pipette de haut en bas, à nouveau dans le puits pour mélanger.
- Une fois que toutes les réactions sont chargées et mélangées, placer la plaque 96 puits dans le fluorimètre et mesurer la fluorescence de point de terminaison sfGFP.
Nota : Les longueurs d’onde d’Excitation et d’émission pour la quantification de fluorescence sfGFP sont 485 nm et 510 nm, respectivement. - À l’aide d’une courbe d’étalonnage générée précédemment, déterminer le [sfGFP] d’après les relevés de fluorescence obtenue.
NOTE : Instructions pour générer la courbe d’étalonnage de concentration de sfGFP contre l’intensité de la fluorescence sont fournies dans les informations complémentaires (supplémentaire Figure 3). Les utilisateurs devront établir une courbe d’étalonnage pour leur instrument, étant donné que la sensibilité de l’instrument peut varier.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Nous avons présenté une sonication basée sur e. coli extrait protocole de préparation qui peut être effectué sur une période de quatre jours, avec la Figure 1 montrant la ventilation procédurale au cours de chaque journée. Il n’y a malléabilité aux étapes qui peut être accomplie chaque jour avec différents points de suspension, mais nous avons trouvé ce flux de travail la plus efficace d’exécuter. En outre, les granules cellulaires (étape 1.3.18) et extrait entièrement préparé (étape 2.10) sont stables à-80 ° C pendant au moins un an, permettant à l’utilisateur de créer de grands stocks de chacun à conserver pour l’utilisation à un temps ultérieur17. Non seulement l’extrait stable sur de longues périodes, mais l’extrait peut également subir au moins cinq gel dégel sans perte significative de la productivité (Figure 4). Cela permet plus grandes aliquotes d’extraits d’être stockées pour des utilisations multiples si l’espace de stockage du congélateur est limité. Cependant, nous recommandons plusieurs petites portions (~ 100 μL) d’extrait lorsque cela est possible.
Avec chaque nouvelle préparation d’extrait, nous recommandons que l’utilisateur effectue un titrage de magnésium afin de déterminer la quantité optimale de magnésium pour ce lot d’extrait. Les utilisateurs peuvent quantifier lot de variabilité de la concentration des protéines totales de l’extrait cellulaire par analyse de Bradford. Pour plus d’extraits, on voit généralement des concentrations de protéines totales de 30 à 50 mg/mL, et dans cet intervalle, il n’y a pas de corrélation évidente entre les concentrations de protéines totales et performance extrait des cellules. Par conséquent, nous recommandons qu’utilisateurs tune teneurs en magnésium en conséquence pour s’assurer que les protéines et acides nucléiques fonctionnalité sont maximisées pour chaque lot d’extrait. Le niveau de magnésium est important pour la bonne réplication, transcription et traduction de l’ADN, mais des niveaux excessifs peuvent nuire à ces processus53. Pour démontrer cette dépendance, nous ont interprété un titrage co de magnésium et extrait de volume afin de déterminer la combinaison optimale qui minimise la quantité d’extrait nécessaire, tout en conservant une réaction productive (Figure 5). De cette expérience, nous vous recommandons d’utiliser 5 µL de l’extrait et 10 mM Mg2 + extrait avec une teneur en protéines totales de 30 mg/mL, afin d’obtenir plus de 1 000 μg/mL de sfGFP.
Notre expérience avec PFC aussi nous a permis de déterminer les étapes dans le protocole qui peut être modifié sans que cela nuise à la productivité globale du système et d’autres qui font partie intégrante pour un système poussé de CFPS performant. Plus particulièrement, le final de l’OD600 de récolte de cellules n’affecte pas sensiblement le résultat final de la réaction de PFC et cellules peuvent facilement être récoltées n’importe où de 2,7 à 4,0 OD600. Cela représente la début phase exponentielle de croissance où concentration ribosome par cellule est la plus élevée et le mécanisme de translation est la plus active pour soutenir la croissance rapide. Cette observation permet aux utilisateurs de flexibilité afin d’optimiser leurs propres procédures. Nous vous recommandons la récolte environ 3,0 OD600 afin de capturer les cellules à une OD600 plus près à 3.3 de la récolte du temps est complet (Figure 2 a). Les variables qui ont d’impact sur les rendements CFPS incluent modèle ADN qualité, taille de bateau de réaction et les quantités relatives des piles ion extrait et le magnésium présente dans la réaction. Nous avons trouvé la qualité d’ADN d’avoir variation notable de lot à lot. Afin de résoudre ce problème, nous recommandons qu’utilisateurs purifient l’ADN via un midi ou maxi prep, suivi d’une étape de nettoyage ADN supplémentaire soit sur la colonne de purification de l’ADN utilisées dans le maxiprep, ou après la purification à l’aide d’un kit de nettoyage ADN supplémentaire. Cela améliore la reproductibilité en qualité d’ADN pour des réactions de PFC et aboutit à la production de protéines plus robuste (Figure 2 b). La cuve de réaction aussi des répercussions des rendements volumétriques, tels que la production de protéines de configurations de réaction identiques dans différents volumes de navire peut différer jusqu'à 40 %. Il a été théorisé que le coup de pouce en rendement volumétrique observée chez les plus gros bateaux est due à une augmentent la surface du mélange réactionnel, permettant d’améliorer l’échange d’oxygène (Figure 2), et d’autres ont encore amplifié les rendements volumétriques en exécutant Réactions de PFC en grandes plaques de fond plat, que nous recommandons pour les réactions sur 100 µL17,31,37,,52.

Figure 1 : Chronologie pour la croissance de la culture, la production d’extrait cellulaire, de montage et de quantification des réactions CFPS. L’utilisateur peut mettre en œuvre la plateforme de produits forestiers certifiés pour leurs applications de recherche par le biais de ce flux de travail de quatre jours. Préparation du réactif représente le principal de l’heure et le coût des investissements pour le premier tour de cette expérience et diminue considérablement après l’établissement de stocks de réactifs. En outre, les granulés de cellule et extrait cellulaire préparée peuvent être conservés pendant plus d’un an à-80 ° C, permettant à l’utilisateur de commencer la chronologie à différentes étapes pour des résultats plus rapides. L’utilisateur peut également mettre en pause à diverses mesures afin de modifier la chronologie de ce flux de travail. S’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.

Figure 2 : Conditions modifiables pour CFPS et les effets sur le rendement volumétrique réaction. A. extrait de comparaison de la productivité basé sur la récolte des cellules BL21 (de3) à diverses lectures de600 OD. Basé sur ce terrain, nous recommandons de récolte à une OD600 de 3,3 à produire au moins 1000 µg/mL de protéine cible. Les réactions ont été réalisées à l’échelle 15 μL dans des microtubes de 1,5 mL. B. comparaison de deux protocoles d’ADN maxiprep lavage avec et sans après la purification ADN-nettoyages. pJL1-sfGFP plasmides a subi une maxiprep avec un ou deux lavages, suivies par un nettoyage après la purification kit de purification de PCR. Pour atteindre environ 900 µg/mL d’expression de la protéine, nous vous suggérons d’effectuer un nettoyage après la purification de l’ADN indépendamment du nombre de lavages maxiprep. Les réactions ont été réalisées à l’échelle 15 μL dans des microtubes de 1,5 mL. C. réactions de CFPS 15 µL effectué dans des récipients divers, allant de 2 mL pour microtubes à 0,6 mL. « Neg » représente un contrôle négatif où aucun modèle d’ADN a été ajouté à la réaction. Toutes les barres d’erreur représentent 1 écart-type des trois réactions indépendantes pour chaque condition, dont chacun a été quantifiée en triple exemplaire. S’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.

Figure 3 : Extraient de clé procédurale parametrage et résultats pour la création de productif. A. configuration correcte du bain d’eau glacée sonication pour assurer un refroidissement de l’échantillon alors que la chaleur est générée au cours de la sonication. B. tube de microtubes de 1,5 mL contenant des cellules resuspendues pellet pré (à gauche) et la sonication post (à droite). Le lysat qui en résulte doit afficher une teinte plus foncée par rapport à culot cellulaire remises en suspension. C. une bonne séparation du surnageant et le culot de lysat après 18 000 x centrifugation g cellulaire. D. réactions de CFPS après 4 h d’incubation à 37 ° C. tube microtubes de 1,5 mL à droite (réaction réussie) montre une fluorescence visible de la protéine de journaliste sfGFP ~ 900 μg/ml. Le tube de contrôle négatif sur la gauche, manque de modèle ADN et simulant une réaction infructueuse, affiche une solution claire avec aucune fluorescence. S’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.

Figure 4 : Extrait de changement dans l’expression de la protéine plus de 5 cycles de gel-dégel de PFC. Extrait préparé à partir de la même croissance a subi cinq gel dégel par azote liquide flash gel suivi de décongélation sur la glace. Aucun changement important dans la productivité extrait pour exprimer des sfGFP ont été observés sur les cinq cycles de gel-dégel. Les réactions ont été réalisées à l’échelle 15 μL dans des microtubes de 1,5 mL. « Neg » représente un contrôle négatif où aucun modèle d’ADN a été ajouté à la réaction. Toutes les barres d’erreur représentent 1 écart-type des trois réactions indépendantes pour chaque condition, dont chacun a été quantifiée en triple exemplaire. S’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.

Figure 5 : Produits forestiers certifiés pour les réactions avec des variables [Mg2+] et extraire des volumes par rapport à [sfGFP]. [Mg2 +] variait de 8 mM à 14 mM par incréments de 2 mM et extraire les volumes variaient de 3 µL à 7 µL avec de1 µL. Le code représente couleur [sfGFP] produit de haut (rouge) en bas (violet) pour. Pour maximiser l’efficacité de réactif tout en maintenant la production de protéines élevé, nous recommandons l’utilisation de 5 µL de l’extrait et 10 mM Mg2+ pour les extraits qui ont une teneur en protéines totales d’environ 30 mg/mL, tel que déterminé par analyse de Bradford. Points d’origine pour générer le tracé d’isolignes reposaient au large de la fluorescence de point de terminaison des trois réactions indépendantes pour chaque condition, dont chacun a été mesuré en triple exemplaire. Les réactions ont été réalisées à l’échelle 15 μL dans des microtubes de 1,5 mL. S’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.
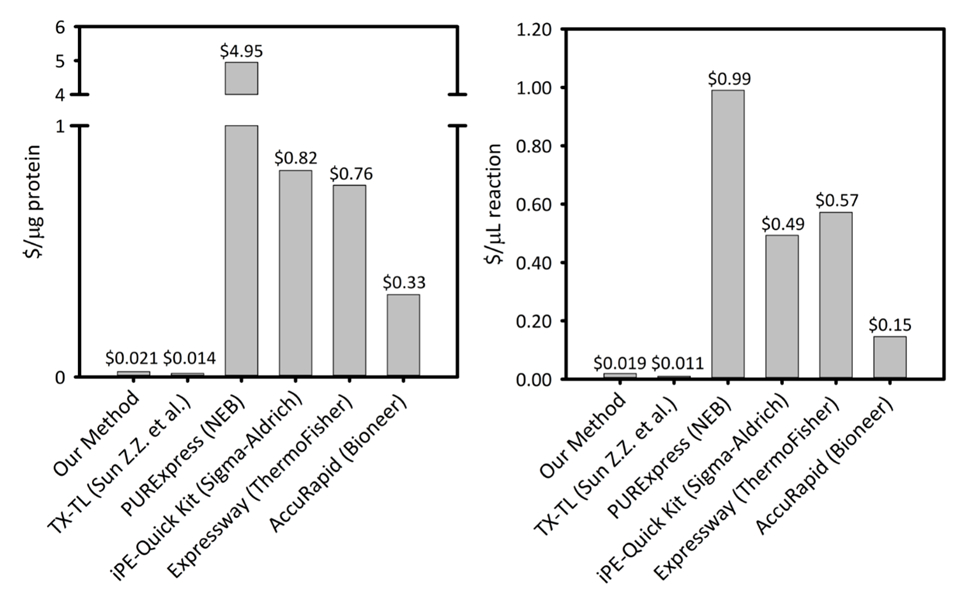
Supplémentaire Figure 1: coût par microgramme de protéine produite et par microlitre de réaction à travers six plateformes de synthèse protéique acellulaire. Notre plateforme est comparée aux cinq cellules différentes protéines libres synthèse kits/plates-formes variant de productivité et les prix. Notre plateforme CFPS sonication basée est plus rentable dans les deux $/ µg de protéines et $/ µL de réaction que commercial la plupart des kits et fournit la facilité d’un kit pour l’installation de la réaction, tout en restant coût comparable à d’autres plates-formes CFPS académiques. S’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.
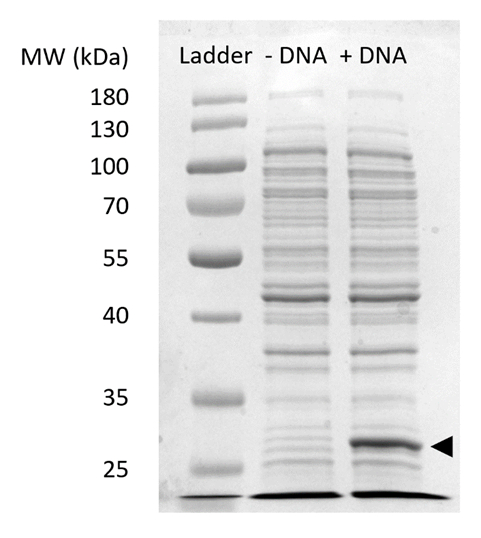
Supplémentaires Figure 2: SDS-PAGE de sfGFP expression dans CFPS. Acellulaire des réactions de synthèse de protéine avec ADN (+) et sans (-ADN) matrice d’ADN de sfGFP ont été exécutés sur un 12 % gel SDSPAGE pour démontrer l’expression des sfGFP observée à 27 kDa (flèche noire). Les techniques traditionnelles de SDS-PAGE ont été utilisées. Chaque échantillon chargées sur le gel extrait inclus 18 µg de protéine totale basée sur la quantification de dosage de Bradford de protéines dans la cellule. Basé sur les mesures d’intensité de fluorescence et notre courbe étalon, nous estimons que la « + ADN » lane contient 0,42 µg de sfGFP. Pour obtenir ces échantillons, réactions de PFC ont été exécutées à l’échelle 15 µL dans des microtubes de 1,5 mL rendements volumétriques conforme à la Figure 3. S’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.
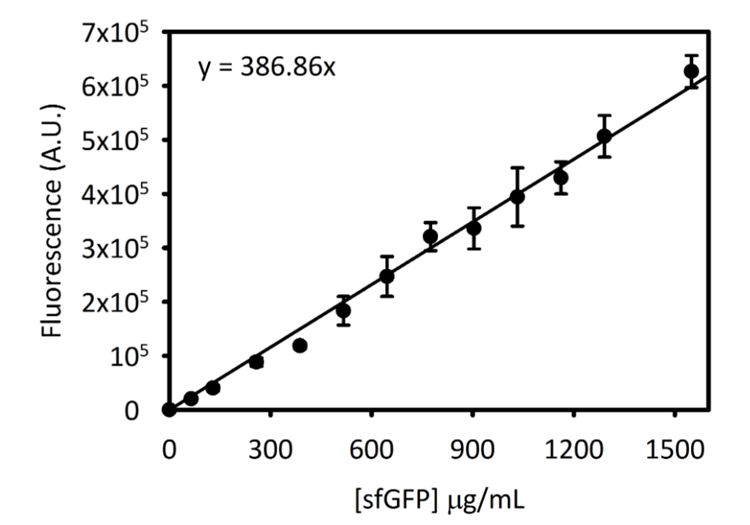
Supplémentaire Figure 3: courbe d’étalonnage pour sfGFP sur 5 Cytation. Cette courbe a été déterminée en utilisant les méthodes décrites ci-dessus. Toutes les données recueillies pour ce manuscrit a été converti à partir des lectures de fluorescence de point de terminaison à [sfGFP] en µg/mL à l’aide de cette courbe d’étalonnage. S’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
La synthèse protéique acellulaire est devenue une technologie puissante et propice pour une variété d’applications allant de la biofabrication de prototypage rapide de systèmes biochimiques. L’ampleur des demandes est pris en charge par la capacité à contrôler, manipuler et d’augmenter la machinerie cellulaire en temps réel. En dépit de l’impact grandissant de cette technologie de plate-forme, adaptation large est restée lente en raison des nuances techniques dans la mise en œuvre des méthodes. Grâce à cet effort, nous visons à fournir des simplicité et clarté pour l’établissement de cette technologie dans les nouveaux laboratoires. À cette fin, notre protocole pour un e. coli-plate-forme de synthèse basé protéine acellulaire peut être réalisé dans un délai de démarrage de quatre jours en laboratoire formé profanes, tels que les étudiants de premier cycle (Figure 1). En outre, une fois produits, un stock de réactifs et extrait des réactions subséquentes de lot des produits forestiers certifiés peuvent être mises en place, incubées et quantifiées en seulement 5 h. Un seul, la croissance des cellules 1 L peut entraîner assez extrait pour les réactions de quatre cents 15 µL de PFC, tandis que les préparations de lot unique des autres réactifs acellulaire peuvent prendre en charge des milliers de réactions. Préparations de réactif peuvent également évoluer vers le haut si un stock encore plus important est nécessaire. Les réactions de PFC peuvent être configuré de manière à haut débit, en utilisant une plaque de 96 puits ou tubes PCR pour l’essai des diverses conditions en parallèle. Les rendements volumétriques vont diminuer lors de l’utilisation de petits bateaux comme le montre la Figure 2. Réactions de PFC peuvent également être transposées de microlitres à quelques dizaines de millilitres du volume total de réaction afin d’augmenter le rendement de protéine totale pour une seule condition. Lors du mesurage de volume, la considération primordiale est que les rendements volumétrique réactions diminuent à mesure que le ratio surface de zone-volume de la réaction diminue37,52. Pour mesurent-vers le haut tout en maintenant des rendements volumétriques similaires d’expression de la protéine, utilisateurs devraient diviser le volume réactionnel en nombreuses cuves de réaction et/ou augmenter la taille des navires. Balances de réaction allant de 15 µL - 100 µL en volume, de nombreuses réactions de 15 µL en parallèle sont recommandées. Pour les réactions dépassant 100 µL en volume, plaques 24 puits à fond plat sont recommandées, et 12 plats sont recommandés pour les volumes de réaction dépassant 600 µL. Ces appariements des volumes de réaction et des bateaux assurent la cohérence des rendements volumétriques réaction à intensification17,31,37,52. Mise à l’échelle vers le haut, au-delà de ces volumes est possible en utilisant plusieurs puits de la plaque en parallèle. Ce format, la réaction peut être dimensionnée au volume total de plus de 10 mL. Optimiser la combinaison de navire de volume-réaction de réaction peut prendre en charge les applications de la biofabrication sans sacrifier la productivité de la réaction.
Lors de l’exécution de ce protocole, il y a quelques facteurs clés qui influent sur les rendements volumétrique de réaction ainsi que des indicateurs associés à peu performants extrait. Afin d’assurer la lyse adéquate et pour éviter la dénaturation de la machinerie de transcription/traduction fonctionnelle, il est important d’atténuer la chaleur produite lors de la lyse. Immerger la remise en suspension des cellules dans un bain d’eau glacée pendant sonication à dissiper rapidement la chaleur au cours de la sonication (Figure 3 a). Un indicateur de la lyse cellulaire efficace est l’émergence d’un aspect plus sombre du lysat cellulaire par rapport aux échantillons pré-sonication (Figure 3 b). Pour la flexibilité de l’utilisateur, le sonicateur et sonde illustré à la Figure 3 a est adaptable à un éventail de volumes de 100 µL à 15 mL de cellules resuspendues. Pour ce faire, l’utilisateur peut ajuster le nombre de joules livré pour la lyse du volume désiré des cellules. En outre, des volumes plus importants de l’extrait peuvent être préparés au moyen de deux approches complémentaires. Les utilisateurs peuvent soniquer tubes multiples en parallèle, et/ou soniquer des volumes plus importants de remise en suspension de cellules, mise à l’échelle de la quantité d’énergie proportionnellement avec le volume comme décrit précédemment29,45. Une autre étape qui indique la qualité de l’extrait est l’étape de centrifugation après la lyse cellulaire. Après la lyse cellulaire, nous vous recommandons de centrifugation à 18 000 x g pour fournir une séparation claire entre le surnageant (machines de transcription/traduction, fragments d’ADN génomique qui ne fonctionnement plus modèle transcription/traduction) et la (pellet considérée comme indésirable des composants cellulaires tels que la membrane cellulaire et les protéines précipitées) (Figure 3). Nous avons trouvé cette centrifugation à 18 000 x g améliore la séparation, résultant en une meilleure reproductibilité par rapport aux tours à basse vitesse tels que 12 000 x g. Pour plus de commodité, nous vous recommandons d’utiliser une centrifugeuse réfrigérée table-top, capable d’atteindre un minimum de 12 000 x g. Cette étape est aussi couramment effectuée à 30 000 x g, qui devrait être envisagée si l’équipement approprié est disponible54,55,56,57,58, 59 , 60. extrait de performance n’est pas affectée par la vitesse de centrifugation à cette étape étant donné que la séparation est atteint. Lorsque vous retirez le surnageant désiré, il est préférable d’éviter tout matériel nuageux qui existent à la frontière entre le surnageant et le culot puisque cette contamination permettra de réduire la productivité de l’extrait. Pureté du surnageant se traduit par des extraits plus productifs et vaut la quantité réduite de l’extrait obtenu pour les nouveaux utilisateurs.
Il est important de noter que tandis que les méthodes que nous avons présentés sont reproductibles et peuvent être exécutées par des scientifiques connaissant un minimum, il peut y avoir de lot et de la réaction à réaction-variation. On peut attribuer à la variation dans la composition de protéomique de la sonication après lysat61. La variabilité de lot que nous avons observés est généralement diminuée après supplémentation avec T7RNAP et optimisation des concentrations en magnésium. Ajout exogène de T7RNAP est fréquente chez les réactions CFPS pour soutenir l’expression protéique optimal, et nous trouvons qu’avoir deux sources de T7RNAP - expression endogène dans les BL21*(DE3) et les T7RNAP supplémentaires à une concentration finale de 16 µg/mL - améliore reproductibilité de lot pour les nouveaux utilisateurs45,46. Avec l’expérience, les utilisateurs peuvent modifier leurs expériences afin d’utiliser uniquement une seule source de T7RNAP si vous le souhaitez. Quantification de la teneur en protéines totales d’un nouveau lot d’extrait et un ajustement approprié de Mg2 + concentration peut également contribuer à diminuer à lot variation volumétrique protein expression avec des rendements. Variations dans l’expression des protéines peuvent également être dues à des différences dans la taille et la structure de la protéine d’intérêt, l’utilisation de codon du gène et son site de liaison de ribosome correspondante du gène d’intérêt, ainsi que le type de vecteur d’expression utilisé62 ,,63. Pour ces raisons, certaines protéines expriment pas ainsi que le sfGFP de protéine modèle, résultant en une réduction du rendement volumétrique de réactions de la PFC.
Limites de la technique CFPS présentée comprennent qu’il ne peut pas être directement adapté à toutes les demandes d’acellulaire, tels que l’ingénierie métabolique et le réglage des conditions de l’expression, sans apporter de modifications supplémentaires aux protocoles. Cependant, nous croyons que ce protocole sera fournissent une base pour l’établissement d’une plateforme PFC dans les nouveaux laboratoires et donner la capacité à mettre en œuvre liminaires acellulaire réactions dans leurs laboratoires des non-spécialistes. Après la mise en oeuvre initiale, les chercheurs peuvent expérience avec la plateforme pour faire leurs propres modifications pour des applications plus spécifiques basées sur les autres documents publiés dans le domaine.
La protéine de 0,021 $/ µg de coûts plate-forme de PFC (excluant le coût de la main-d'œuvre et l’équipement), ce qui rend notre système compétitifs avec les kits commerciaux sans compromettre la facilité d’installation de la réaction. Évaluations des coûts comparatifs par µL de réaction montrent des tendances semblables (Supplemental Figure 1). Nous estimons les coûts de démarrage pour être ~ 4 500 $ pour tous les réactifs et un supplémentaire $ 3 200 pour des équipements spécialisés, tels qu’un sonicateur. Heures-personnes pour effectuer cette procédure sont estimés à ~ 26 h pour toutes les préparation de réactif par rapport au sol vers le haut. Cependant, une fois que les stocks importants de réactifs ont été préparés, exigences sur le travail diminuent sensiblement. En outre, comme l’expérience avec la plate-forme est acquise, nous recommande l’intensification de la taille de la croissance cellulaire, extraire la préparation et la préparation des réactifs pour maximiser l’efficacité de l’époque. Vu les coûts de démarrage, nous vous recommandons la plateforme PFC pour des applications en biologie synthétique, efforts de haut débit, et les conditions d’expression de protéine qui sont incompatibles avec les plates-formes d’expression traditionnelle de protéines due à l’encontre de la cellule contraintes de biochimie et de la viabilité. Dans ces cas spécialisés où la technique souhaitée est activée par la plateforme PFC, le coût plus élevé des PFC dans vivo surexpression est justifiée.
Poursuite du développement de la plateforme PFC est susceptible d’offrir un utilitaire plus large aux efforts de biotechnologie telles que l’ingénierie métabolique des voies enzymatiques, production et caractérisation des protéines insolubles traditionnellement, des acides aminés non standard incorporation et expression de la protéine anormale, fabrication de médecine stratifiée et s’étendant au-delà du laboratoire dans la salle de classe pour l’enseignement de souches64,65,66. Ces efforts seront appuiera davantage sur les efforts en cours pour une caractérisation détaillée de la plateforme PFC. Une meilleure compréhension de la composition de l’extrait cellulaire donnera lieu à un raffinement continu vers la réaction amélioré les rendements et la souplesse dans la réaction des conditions61,67,68.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Les auteurs déclarent qu’ils ont pas concurrentes d’intérêts financiers ou autres conflits d’intérêts.
Acknowledgments
Auteurs tient à remercier Dr. Jennifer VanderKelen, Andrea Laubscher et Tony Turretto pour le support technique, Wesley Kao, Layne Williams et Christopher Hight pour discussions utiles. Auteurs reconnaissent également de financement du projet de loi et Linda Frost fonds, centre pour les Applications de la biotechnologie Chevron biotechnologie appliquée recherche subvention de dotation, Cal Poly recherche, savantes, et programme de subvention des activités créatives (RSCA 2017), et la National Science Foundation (NSF-1708919). MZL reconnaît les subventions supérieures de l’Université California état. MCJ reconnaît l’Army Research Office W911NF-16-1-0372, National Science Foundation accorde MCB-1413563 et MCB-1716766, l’armée de l’Air recherche laboratoire centre d’Excellence Grant FA8650-15-2-5518, la subvention de l’agence défense menace la réduction HDTRA1-15-10052/P00001, la David and Lucile Packard Foundation, le programme d’enseignant-chercheur de Camille Dreyfus, le ministère de l’énergie BER Grant DE-SC0018249, le programme de sciences humaines de frontières (RGP0015/2017), la subvention ETOP DOE Joint Genome Institute, et le Consortium biomédicale de Chicago avec prise en charge par les fonds de Searle à la Chicago Community Trust pour la prise en charge.
Materials
| Name | Company | Catalog Number | Comments |
| Luria Broth | ThermoFisher | 12795027 | |
| Tryptone | Fisher Bioreagents | 73049-73-7 | |
| Yeast Extract | Fisher Bioreagents | 1/2/8013 | |
| NaCl | Sigma-Aldrich | S3014-1KG | |
| Potassium Phosphate Dibasic | Sigma-Aldrich | 60353-250G | |
| Potassium Phosphate Monobasic | Sigma-Aldrich | P9791-500G | |
| D-Glucose | Sigma-Aldrich | G8270-1KG | |
| KOH | Sigma-Aldrich | P5958-500G | |
| IPTG | Sigma-Aldrich | I6758-1G | |
| Mg(OAc)2 | Sigma-Aldrich | M5661-250G | |
| K(OAc) | Sigma-Aldrich | P1190-1KG | |
| Tris(OAc) | Sigma-Aldrich | T6066-500G | |
| DTT | ThermoFisher | 15508013 | |
| tRNA | Sigma-Aldrich | 10109541001 | |
| Folinic Acid | Sigma-Aldrich | F7878-100MG | |
| NTPs | ThermoFisher | R0481 | |
| Oxalic Acid | Sigma-Aldrich | P0963-100G | |
| NAD | Sigma-Aldrich | N8535-15VL | |
| CoA | Sigma-Aldrich | C3144-25MG | |
| PEP | Sigma-Aldrich | 860077-250MG | |
| K(Glu) | Sigma-Aldrich | G1501-500G | |
| NH4(Glu) | MP Biomedicals | 02180595.1 | |
| Mg(Glu)2 | Sigma-Aldrich | 49605-250G | |
| Spermidine | Sigma-Aldrich | S0266-5G | |
| Putrescine | Sigma-Aldrich | D13208-25G | |
| HEPES | ThermoFisher | 11344041 | |
| Molecular Grade Water | Sigma-Aldrich | 7732-18-5 | |
| L-Aspartic Acid | Sigma-Aldrich | A7219-100G | |
| L-Valine | Sigma-Aldrich | V0500-25G | |
| L-Tryptophan | Sigma-Aldrich | T0254-25G | |
| L-Phenylalanine | Sigma-Aldrich | P2126-100G | |
| L-Isoleucine | Sigma-Aldrich | I2752-25G | |
| L-Leucine | Sigma-Aldrich | L8000-25G | |
| L-Cysteine | Sigma-Aldrich | C7352-25G | |
| L-Methionine | Sigma-Aldrich | M9625-25G | |
| L-Alanine | Sigma-Aldrich | A7627-100G | |
| L-Arginine | Sigma-Aldrich | A8094-25G | |
| L-Asparagine | Sigma-Aldrich | A0884-25G | |
| Glycine | Sigma-Aldrich | G7126-100G | |
| L-Glutamine | Sigma-Aldrich | G3126-250G | |
| L-Histadine | Sigma-Aldrich | H8000-25G | |
| L-Lysine | Sigma-Aldrich | L5501-25G | |
| L-Proline | Sigma-Aldrich | P0380-100G | |
| L-Serine | Sigma-Aldrich | S4500-100G | |
| L-Threonine | Sigma-Aldrich | T8625-25G | |
| L-Tyrosine | Sigma-Aldrich | T3754-100G | |
| Fisherbrand Premium Microcentrifuge Tubes: 2.0 mL | Fisher Scientific | 05-408-138 | |
| Fisherbrand Premium Microcentrifuge Tubes: 1.5 mL | Fisher Scientific | 05-408-129 | |
| Fisherbrand Premium Microcentrifuge Tubes: 0.6 mL | Fisher Scientific | 05-408-120 | |
| PureLink HiPure Plasmid Prep Kit | ThermoFisher | K210007 | |
| Ultrasonic Processor | QSonica | Q125-230V/50Hz | 3.175 mm diameter probe |
| Avanti J-E Centrifuge | Beckman Coulter | 369001 | |
| JLA-8.1000 Rotor | Beckman Coulter | 366754 | |
| 1L Centrifuge Tube | Beckman Coulter | A99028 | |
| Tunair 2.5L Baffeled Shake Flask | Sigma-Aldrich | Z710822 | |
| Microfuge 20 | Beckman Coulter | B30134 | |
| New Brunswick Innova 42/42R Incubator | Eppendorf | M1335-0000 | |
| Cytation 5 | BioTek | ||
| Strep-Tactin XT Starter Kit | IBA | 2-4998-000 | |
| pJL1-sfGFP | Addgene | 69496 | |
| BL21(DE3) | New England BioLabs |
References
- Jiang, L., Zhao, J., Lian, J., Xu, Z. Cell-free protein synthesis enabled rapid prototyping for metabolic engineering and synthetic biology. Synthetic and Systems Biotechnology. 3 (2), 90-96 (2018).
- Carlson, E. D., Gan, R., Hodgman, C. E., Jewett, M. C. Cell-free protein synthesis: Applications come of age. Biotechnology Advances. 30 (5), 1185-1194 (2012).
- Watanabe, M. Cell-Free Protein Synthesis for Structure Determination by X-ray Crystallography. Methods in molecular biology (Clifton, N.J). 607, 149-160 (2010).
- Martemyanov, K. A., Shirokov, V. A., Kurnasov, O. V., Gudkov, A. T., Spirin, A. S. Cell-Free Production of Biologically Active Polypeptides: Application to the Synthesis of Antibacterial Peptide Cecropin. Protein Expression and Purification. 21 (3), 456-461 (2001).
- Renesto, P., Raoult, D. From genes to proteins: in vitro expression of rickettsial proteins. Annals of the New York Academy of Sciences. 990, 642-652 (2003).
- Xu, Z., Chen, H., Yin, X., Xu, N., Cen, P. High-Level Expression of Soluble Human b-Defensin-2 Fused With Green Fluorescent Protein in Escherichia coli Cell-Free System. Applied Biochemistry and Biotechnology. 127 (1), 053-062 (2005).
- Baumann, A. In-situ observation of membrane protein folding during cell-free expression. PLoS ONE. 11 (3), 1-15 (2016).
- Wang, Y., Percival, Y. H. P. Cell-free protein synthesis energized by slowly-metabolized maltodextrin. BMC Biotechnology. 9, 1-8 (2009).
- Whittaker, J. W. Cell-free protein synthesis: the state of the art. Biotechnology Letters. 35 (2), 143-152 (2013).
- Martin, R. W. Cell-free protein synthesis from genomically recoded bacteria enables multisite incorporation of noncanonical amino acids. Nature Communications. 9 (1), 1203 (2018).
- Kwon, Y. C., Song, J. K., Kim, D. M. Cloning-Independent Expression and Screening of Enzymes Using Cell-Free Protein Synthesis Systems. Methods in Molecular Biology. 1118, 97-108 (2014).
- Chappell, J., Jensen, K., Freemont, P. S. Validation of an entirely in vitro approach for rapid prototyping of DNA regulatory elements for synthetic biology. Nucleic Acids Research. 41 (5), 3471-3481 (2013).
- Takahashi, M. K. Characterizing and prototyping genetic networks with cell-free transcription-translation reactions. Methods. 86, 60-72 (2015).
- Karim, A. S., Jewett, M. C. A cell-free framework for rapid biosynthetic pathway prototyping and enzyme discovery. Metabolic Engineering. 36, 116-126 (2016).
- Dudley, Q. M., Anderson, K. C., Jewett, M. C. Cell-Free Mixing of Escherichia coli Crude Extracts to Prototype and Rationally Engineer High-Titer Mevalonate Synthesis. ACS Synthetic Biology. 5 (12), 1578-1588 (2016).
- Pardee, K. Paper-based synthetic gene networks. Cell. 159 (4), 940-954 (2014).
- Zawada, J. F. Microscale to manufacturing scale-up of cell-free cytokine production-a new approach for shortening protein production development timelines. Biotechnology and Bioengineering. 108 (7), 1570-1578 (2011).
- Sullivan, C. J. A cell-free expression and purification process for rapid production of protein biologics. Biotechnology Journal. 11 (2), 238-248 (2016).
- Li, J. Cell-free protein synthesis enables high yielding synthesis of an active multicopper oxidase. Biotechnology Journal. 11 (2), 212-218 (2016).
- Heinzelman, P., Schoborg, J. A., Jewett, M. C. pH responsive granulocyte colony-stimulating factor variants with implications for treating Alzheimer's disease and other central nervous system disorders. Protein Engineering Design and Selection. 28 (10), 481-489 (2015).
- Pardee, K. Low-Cost Detection of Zika Virus Using Programmable Biomolecular Components. Cell. 165 (5), 1255-1266 (2016).
- Slomovic, S., Pardee, K., Collins, J. J. Synthetic biology devices for in vitro and in vivo diagnostics. Proceedings of the National Academy of Sciences. 112 (47), 14429-14435 (2015).
- Gootenberg, J. S. Nucleic acid detection with CRISPR-Cas13a/C2c2. Science. 356 (6336), 438-442 (2017).
- Pardee, K. Portable, On-Demand Biomolecular Manufacturing. Cell. 167 (1), 248-259 (2016).
- Karig, D. K., Bessling, S., Thielen, P., Zhang, S., Wolfe, J. Preservation of protein expression systems at elevated temperatures for portable therapeutic production. Journal of the Royal Society, Interface. 14 (129), (2017).
- Smith, M. T., Berkheimer, S. D., Werner, C. J., Bundy, B. C. Lyophilized Escherichia coli-based cell-free systems for robust, high-density, long-term storage. BioTechniques. 56 (4), 186-193 (2014).
- Hunt, J. P., Yang, S. O., Wilding, K. M., Bundy, B. C. The growing impact of lyophilized cell-free protein expression systems. Bioengineered. 8 (4), 325-330 (2017).
- Stark, J. C. BioBitsTM Bright: A fluorescent synthetic biology education kit. Science Advances. 4 (8), (2018).
- Huang, A. BioBitsTM Explorer: A modular synthetic biology education kit. Science Advances. 4 (8), (2018).
- Zemella, A., Thoring, L., Hoffmeister, C., Kubick, S. Cell-Free Protein Synthesis: Pros and Cons of Prokaryotic and Eukaryotic Systems. ChemBioChem. 16 (17), 2420-2431 (2015).
- Oza, J. P. Robust production of recombinant phosphoproteins using cell-free protein synthesis. Nature Communications. 6 (1), 8168 (2015).
- Zemella, A. Cell-free protein synthesis as a novel tool for directed glycoengineering of active erythropoietin. Scientific Reports. 8 (1), 8514 (2018).
- Jaroentomeechai, T. Single-pot glycoprotein biosynthesis using a cell-free transcription-translation system enriched with glycosylation machinery. Nature Communications. 9 (1), 2686 (2018).
- Kightlinger, W. Design of glycosylation sites by rapid synthesis and analysis of glycosyltransferases. Nature Chemical Biology. 14 (6), 627-635 (2018).
- Schoborg, J. A. A cell-free platform for rapid synthesis and testing of active oligosaccharyltransferases. Biotechnology and Bioengineering. 115 (3), 739-750 (2018).
- Chekulayeva, M. N., Kurnasov, O. V., Shirokov, V. A., Spirin, A. S. Continuous-Exchange Cell-Free Protein-Synthesizing System: Synthesis of HIV-1 Antigen Nef. Biochemical and Biophysical Research Communications. 280 (3), 914-917 (2001).
- Hong, S. H. Improving Cell-Free Protein Synthesis through Genome Engineering of Escherichia coli Lacking Release Factor 1. ChemBioChem. 16 (5), 844-853 (2015).
- Endo, Y., Otsuzuki, S., Ito, K., Miura, K. Production of an enzymatic active protein using a continuous flow cell-free translation system. Journal of Biotechnology. 25 (3), 221-230 (1992).
- Volyanik, E. V., Dalley, A., Mckay, I. A., Leigh, I., Williams, N. S., Bustin, S. A. Synthesis of Preparative Amounts of Biologically Active Interleukin-6 Using a Continuous-Flow Cell-Free Translation System. Analytical Biochemistry. 214 (1), 289-294 (1993).
- Martin, G. A., Kawaguchi, R., Lam, Y., DeGiovanni, A., Fukushima, M., Mutter, W. High-yield, in vitro protein expression using a continuous-exchange, coupled transcription/ translation system. BioTechniques. 31 (4), 948-950 (2001).
- Stech, M., Quast, R. B., Sachse, R., Schulze, C., Wüstenhagen, D. A., Kubick, S. A Continuous-Exchange Cell-Free Protein Synthesis System Based on Extracts from Cultured Insect Cells. PLoS ONE. 9 (5), e96635 (2014).
- Quast, R. B., Sonnabend, A., Stech, M., Wüstenhagen, D. A., Kubick, S. High-yield cell-free synthesis of human EGFR by IRES-mediated protein translation in a continuous exchange cell-free reaction format. Scientific Reports. 6 (1), 30399 (2016).
- Thoring, L., Dondapati, S. K., Stech, M., Wüstenhagen, D. A., Kubick, S. High-yield production of "difficult-to-express" proteins in a continuous exchange cell-free system based on CHO cell lysates. Scientific Reports. 7 (1), 11710 (2017).
- Hoffmann, M., Nemetz, C., Madin, K., Buchberger, B. Rapid translation system: A novel cell-free way from gene to protein. Biotechnology annual review. 10, 1-30 (2004).
- Kwon, Y. C., Jewett, M. C. High-throughput preparation methods of crude extract for robust cell-free protein synthesis. Scientific Reports. 5 (1), 8663 (2015).
- Katsura, K. A reproducible and scalable procedure for preparing bacterial extracts for cell-free protein synthesis. Journal of Biochemistry. 162 (June), 357-369 (2017).
- Fujiwara, K., Doi, N. Biochemical preparation of cell extract for cell-free protein synthesis without physical disruption. PLoS ONE. 11 (4), 1-15 (2016).
- Shrestha, P., Holland, T. M., Bundy, B. C. Streamlined extract preparation for Escherichia coli-based cell-free protein synthesis by sonication or bead vortex mixing. BioTechniques. 53 (3), 163-174 (2012).
- Krinsky, N. A Simple and Rapid Method for Preparing a Cell-Free Bacterial Lysate for Protein Synthesis. PLoS ONE. 11 (10), (2016).
- Jewett, M. C., Swartz, J. R. Mimicking the Escherichia coli cytoplasmic environment activates long-lived and efficient cell-free protein synthesis. Biotechnology and Bioengineering. 86 (1), 19-26 (2004).
- Swartz, J. R., Jewett, M. C., Woodrow, K. A. Cell-Free Protein Synthesis With Prokaryotic Combined Transcription-Translation. Recombinant Gene Expression. (267), 169-182 (2004).
- Voloshin, A. M., Swartz, J. R. Efficient and scalable method for scaling up cell free protein synthesis in batch mode. Biotechnology and Bioengineering. 91 (4), 516-521 (2005).
- Vernon, W. B. The role of magnesium in nucleic-acid and protein metabolism. Magnesium. 7 (5-6), 234-248 (1988).
- Pratt, J. M. Transcription and Translation: A Practical Approach. , IRL Press. New York. (1984).
- Kim, D. M., Kigawa, T., Choi, C. Y., Yokoyama, S. A Highly Efficient Cell-Free Protein Synthesis System from Escherichia coli. European Journal of Biochemistry. 239 (3), 881-886 (1996).
- Shin, J., Noireaux, V. Efficient cell-free expression with the endogenous E. Coli RNA polymerase and sigma factor 70. Journal of Biological Engineering. 4 (1), 8 (2010).
- Zubay, G. In Vitro Synthesis of Protein in Microbial Systems. Annual Review of Genetics. 7 (1), 267-287 (1973).
- Kigawa, T. Preparation of Escherichia coli cell extract for highly productive cell-free protein expression. Journal of Structural and Functional Genomics. 5 (1/2), 63-68 (2004).
- Liu, D. V., Zawada, J. F., Swartz, J. R. Streamlining Escherichia Coli S30 Extract Preparation for Economical Cell-Free Protein Synthesis. Biotechnology Progress. 21 (2), 460-465 (2008).
- Yang, W. C., Patel, K. G., Wong, H. E., Swartz, J. R. Simplifying and streamlining Escherichia coli-based cell-free protein synthesis. Biotechnology Progress. 28 (2), 413-420 (2012).
- Foshag, D. The E. coli S30 lysate proteome: A prototype for cell-free protein production. New Biotechnology. 40 (Pt B), 245-260 (2018).
- Caschera, F. Bacterial cell-free expression technology to in vitro systems engineering and optimization. Synthetic and Systems Biotechnology. 2 (2), 97-104 (2017).
- Chizzolini, F., Forlin, M., Yeh Martín, N., Berloffa, G., Cecchi, D., Mansy, S. S. Cell-Free Translation Is More Variable than Transcription. ACS Synthetic Biology. 6 (4), 638-647 (2017).
- Hong, S. H., Kwon, Y. C., Jewett, M. C. Non-standard amino acid incorporation into proteins using Escherichia coli cell-free protein synthesis. Frontiers in Chemistry. 2, 34 (2014).
- Sawasaki, T., Ogasawara, T., Morishita, R., Endo, Y. A cell-free protein synthesis system for high-throughput proteomics. Proceedings of the National Academy of Sciences of the United States of America. 99 (23), 14652-14657 (2002).
- Dudley, Q. M., Karim, A. S., Jewett, M. C. Cell-free metabolic engineering: Biomanufacturing beyond the cell. Biotechnology Journal. 10 (1), 69-82 (2015).
- Garcia, D. C. Elucidating the potential of crude cell extracts for producing pyruvate from glucose. Synthetic Biology. 3 (1), 1-9 (2018).
- Hurst, G. B. Proteomics-Based Tools for Evaluation of Cell-Free Protein Synthesis. Analytical Chemistry. 89 (21), 11443-11451 (2017).

{kind=link}
{kind=link}
{kind=link}