

Summary
פרוטוקול זה מפרט את השלבים, ' עלויות ' הציוד הנדרש לייצר e. coli-המבוסס על תמציות תא וליישם במבחנה תגובות סינתזת חלבון תוך 4 ימים או פחות. למנף את אופי גמיש הפלטפורמה עבור יישומים רחב, נדון תנאי ריאקציה, כי ניתן להתאים אופטימיזציה.
Abstract
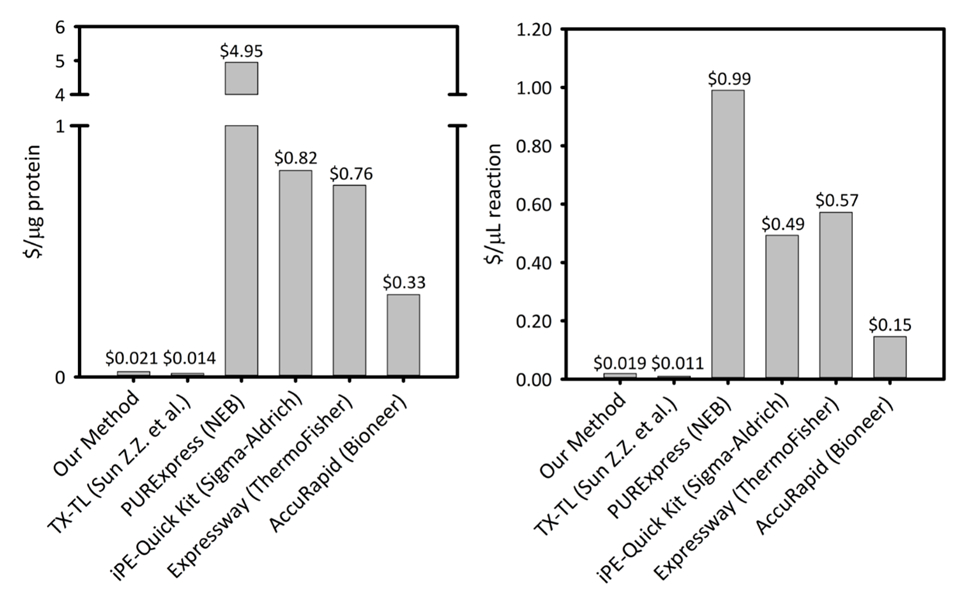
במהלך 50 השנים האחרונות, סינתזה של חלבון נטול תאים (CFPS) התפתחה טכנולוגיה רבת עוצמה כדי לרתום את קיבולת גנים ברמת השעתוק והתרגום של תאים בתוך מבחנה. על ידי obviating הצורך לשמר את הכדאיות של התא, על ידי ביטול המכשול הסלולר, CFPS כבר למעין המתעוררים יישומים ב- biomanufacturing של חלבונים מאתגרת באופן מסורתי, כמו גם יישומים שטנץ מהירה עבור הנדסה מטבולית, גנומיקה תפקודית. השיטות שלנו ליישום של e. coli-מבוסס פלטפורמת CFPS לאפשר למשתמשים חדשים רבים של יישומים אלה. כאן, אנו מתארים שיטות להכין תמצית באמצעות מדיה מועשר מבחנות במבוכה, שיטה ישימה של פירוק התא מבוסס sonication tunable. זו תמצית ואז ניתן ביטוי חלבון מסוגל לייצר µg 900/mL או יותר של חלבון פלואורסצנטי ירוק סופר תיקיה (sfGFP) ברק 5 שעות מהגדרת הניסוי כדי ניתוח נתונים, בהתחשב בכך ריאגנט המתאים מניות שהוכנו מראש. העלות המשוערת הפעלה של קבלת ריאגנטים הוא 4,500 דולר אשר יתמוך אלפי תגובות בעלות של $0.021 לכל µg של חלבון המיוצר או $0.019 לכל µL של תגובה. בנוסף, השיטות ביטוי חלבון מראה את הקלות של ההתקנה התגובה ראיתי במערכות זמינים מסחרית עקב אופטימיזציה של ריאגנט תערובות מראש, בכל חלק של העלות. כדי לאפשר למשתמש למנף את אופי גמיש פלטפורמת CFPS עבור יישומים רחב, זיהינו במגוון היבטים של פלטפורמת שניתן מכוון או מותאם בהתאם המשאבים הזמינים לבין התוצאות של ביטוי חלבון הרצוי.
Introduction
סינתזת חלבון נטול תאים (CFPS) התפתחה טכנולוגיה נעול מספר הזדמנויות חדשות עבור ייצור החלבון, גנומיקה תפקודית, הנדסה מטבולית ועוד בתוך1,50 שנה שעברה2. לעומת תקן ויוו פלטפורמות ביטוי חלבון, CFPS מספק שלושה יתרונות מרכזיים: 1) הטבע ללא תא של הפלטפורמה מאפשרת הייצור של חלבונים זה יהיה פוטנציאל רעיל או זרות תא3,4 5, ,6; 2) איון של הדנ א, ההקדמה של תבנית ה-DNA קידוד של gene (s) עניין לנתב את כל האנרגיה מערכתית בתוך התגובה לייצור של protein(s) עניין; ו 3) מהות הפלטפורמה הפתוחה מאפשר למשתמש לשנות ולנטר את תנאי ריאקציה ו קומפוזיציה בזמן אמת7,8. זו גישה ישירה אל התגובה תומך את הגדלת של מערכות ביולוגיות עם תנאים חמצון-חיזור לייצור חלבונים הרומן, כוונון של תהליכים מטבוליים2,9, וא מורחב 10. ישיר גישה גם מאפשר למשתמש לשלב את התגובה CFPS עם פעילות מבחני במערכת יחיד-סיר עבור מהירה יותר-מבחן עיצוב לבנות מחזורי11. היכולת לבצע את התגובה CFPS נפח קטן טיפות או במכשירים המבוססים על נייר נוספת תומך במאמצי הגילוי תפוקה גבוהה, שטנץ מהירה12,13,14,15 ,16. בשל יתרונות אלה ואת אופי הכנס-הפעל של המערכת, CFPS באופן ייחודי איפשר מגוון רחב של יישומי ביוטכנולוגיה למשל, ייצור חלבונים שקשה לבטא solubly ויוו17, 18,19,20, הגילוי של המחלה21,22,23, על פי דרישה biomanufacturing18,24 ,25,26,27, ו חינוך28,29, אשר כולם להראות את הגמישות ואת השירות של פלטפורמת נטול תאים.
CFPS מערכות יכול להיווצר מתוך מגוון של lysates גסה של שני הקווים תא prokaryotic, האיקריוטים. דבר זה מאפשר אפשרויות מגוונות במערכת של בחירה, שלכל אחד מהם יש יתרונות וחסרונות בהתאם ליישום של עניין. CFPS מערכות גם שוני רב בין זמן הכנה, עלות, יעילות. הכי נפוץ מנוצל תא תמציות מופקות מן נבט חיטה, ארנב רטיקולוציט, חרקים תאים ותאים Escherichia coli , הלה. החסכוני ביותר לתאריך תוך כדי לייצר את התשואות הנפחי הגבוה ביותר של חלבון30 . בעוד מערכות אחרות CFPS יכול להיות יתרון עבור מכונות מולדת השינוי post-translational שלהם, המתעוררים יישומים המשתמשים של e. coli-מכונות מבוסס מסוגלים לגשר על הפער על ידי יצירת site-specifically phosphorylated, חלבונים glycosylated על פי דרישה31,32,33,34,35.
תגובות CFPS ניתן להפעיל גם אצווה, רציף-exchange תא-חינם (CECF) או תבניות (CFCF) ללא תאים זרימה רציפה. הפורמט אצווה הוא מערכת סגורה החיים התגובה שלו הוא מוגבל בשל שמפחית כמויות המגיבים ואת ההצטברות של תוצרי לוואי המעכבת של התגובה. CECF ושיטות CFCF להגדיל את אורך החיים של התגובה, ובכך לגרום התשואות חלבון נפח מוגבר לעומת התגובה אצווה. זו מושגת על-ידי מתן לוואי של סינתזת חלבונים להסיר מיכל התגובה בזמן המגיבים חדשים מסופקים לאורך כל הקורס של התגובה2. במקרה של CFCF, החלבון של עניין ניתן גם להסיר מן החדר התגובה, תוך כדי CECF, החלבון שרידי עניין בבית הבליעה התגובה המורכב של36,קרום חדיר למחצה37. שיטות אלה יקרים במיוחד להתגבר על עניים התשואות הנפחי של חלבונים-קשה-אקספרס של עניין38,39,40,41,42, 43. האתגרים ביישום הגישות CECF ו- CFCF הם 1) בעוד הם תוצאה של שימוש יעיל יותר של מכונות ביו אחראי תמלול ותרגום, הם דורשים כמויות גדולות יותר ובמיוחד של ריאגנטים המגבירה העלות הכוללת ו- 2) הם דורשים setups התגובה מורכבים יותר, ציוד מיוחד לעומת אצווה תבנית44. על מנת להבטיח נגישות עבור משתמשים חדשים, הפרוטוקולים המפורטים להלן המיקוד על הפורמט אצווה בעוצמות התגובה של µL 15 עם המלצות ספציפיות להגברת שעוצמת התגובה הסולם מיליליטר.
השיטות שהוצגו במסמך זה לאפשר שאינם מומחים עם מיומנויות מעבדה בסיסיות (כגון סטודנטים לתואר ראשון) ליישם את צמיחת תאים, לחלץ הכנה של אצווה תבנית התגובה ההתקנה עבור e. coli-CFPS מערכת מבוססת. גישה זו היא חסכונית לעומת ערכות זמינים מסחרית מבלי להתפשר על הקלות של התגובה מבוסס ערכת ההתקנה. יתר על כן, גישה זו מאפשרת יישומים במעבדה ובשדה. כאשר אתה מחליט ליישם CFPS, משתמשים חדשים כדאי ביסודיות להעריך את היעילות של חלבון קונבנציונאלי מערכות ביטוי להשקעה הפעלה, כפי CFPS לא יכול להיות מעולה בכל מקרה. CFPS בשיטות המתוארות כאן לאפשר למשתמש ליישם באופן ישיר מגוון רחב של יישומים, כולל גנומיקה תפקודית, בדיקות, ייצור חלבונים שאינן פתירות ויוו הביטוי, כמו גם שדה תפוקה גבוהה יישומים כולל ביולוגיים וערכות חינוכיות עבור ביולוגיה סינתטית. יישומים נוספים כגון הנדסה מטבולית, כוונון של תנאי ביטוי חלבון, גילוי מחלות ושימוש הסולם בשיטות CECF או CFCF עדיין אפשריים אך עשויים לדרוש ניסיון עם פלטפורמת CFPS עבור שינוי נוסף של התגובה תנאים. השיטות שלנו לשלב הגידול, צלוחיות, בושם במבוכה ומדיה מועשר עם יחסית מהירה ושיטות לשחזור של פירוק התא דרך sonication, ואחריו מלכודת התגובה CFPS מפושטת אשר מנצל premixes ממוטבת45. בעוד השיטות צמיחה סלולרי צריך להיות סטנדרטית במידה מסוימת בתוך שדה זה, שיטות פירוק תאים משתנים במידה רבה. בנוסף sonication, שיטות פירוק הנפוצות כוללות הניצול של מכבש צרפתי, עם מהמגן, חלמוני חרוז, או ליזוזים אחרים שיבוש ביוכימי ופיזית שיטות46,47,48, 49. באמצעות השיטות שלנו, כ 2 מ ל תמצית תא גולמי מתקבלים לכל 1 ליטר של תאים. כמות זו של תמצית התא יכול לתמוך ארבע מאות תגובות CFPS µL 15, כל לעשיית ~ 900 µg/mL של הכתב sfGFP חלבון מן התבנית פלסמיד pJL1-sfGFP. בשיטה זו עלויות $0.021/µg של sfGFP מיוצר (.019$ / µL של תגובה), ללא העלות של עבודה וציוד (משלימה איור 1). מתחיל מאפס, בשיטה זו ניתן ליישם כבר 4 ימים על ידי אדם אחד וחזור CFPS תגובות יכולה להסתיים תוך שעות (איור 1). בנוסף, ניתן לשנות את הפרוטוקול באמצעי אחסון עבור אצוות גדול יותר של ריאגנט הכנה לצרכים של המשתמש. חשוב, ניתן ליישם את הפרוטוקול המובאת כאן על ידי מעבדה שהוכשרו הלא-מומחים כגון סטודנטים לתואר ראשון, כפי שהוא רק דורש מיומנויות מעבדה בסיסית. ההליכים המתוארים להלן ווידאו המלווה את פותחו במיוחד כדי לשפר את הנגישות של פלטפורמת CFPS e. coli לשימוש רחב.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. התקשורת והכנה צמיחת תאים
-
יום 1
- רצף של e. coli BL21*(DE3) תאים גליצרול במניה על גבי צלחת אגר LB ואת תקופת דגירה של פחות 18 h ב- 37 מעלות צלזיוס.
- הכנת 50 מ ל LB ומדיה אוטוקלב הפתרון על מחזור הנוזל למשך 30 דקות-121 מעלות צלזיוס. לאחסן בטמפרטורת החדר.
-
יום 2
- להכין 750 מ של מדיה x YTP 2 ו- 250 מ של תמיסת M D-גלוקוז 0.4 כפי שמתואר המידע משלימה.
- שופכים את המדיה x YTP 2 לתוך הבקבוק במבוכה בלוק של 2.5 L והפתרון D-גלוקוז לתוך בקבוק זכוכית בלוק 500 מ"ל. אוטוקלב שני פתרונות על מחזור הנוזל למשך 30 דקות-121 מעלות צלזיוס.
- ודא כי שני פתרונות סטרילי מאוחסנים 37 ° C אם צמיחת תאים המבוצעת ביום הבא, כדי להגדיל את שיעורי צמיחה על חיסון. אל תשלבו פתרונות עד חיסון.
הערה: פתרונות שניתן לאחסן ב 4 מעלות צלזיוס במשך 1-2 d במקרה הצורך, למרות התקשורת x YTP 2 הוא מאד מועדים לזיהום. - התחל תרבות לילה של BL21(DE3) מזריקים 50 מ של מדיה ליברות עם מושבה בודדת של BL21(DE3) באמצעות לולאה סטיריליים סטרילי טכניקה כדי למנוע זיהום.
- למקם את 50 מ של תרבות BL21*(DE3) ליברות 37 ° C 250 סל"ד רועד חממה ולגדול ללילה עבור h 15-18.
- להכין ולחטא את כל החומרים הנדרשים ימים 3 ו- 4, כולל: שני בקבוקים צנטריפוגה 1 ליטר, 4 x 50 מל קר חרוט צינורות (שוקלים וגושים שיא של שלוש), צינורות microfuge 1.5 mL רבים.
-
יום 3
- הסר התרבות לילה 50 מ של BL21*(DE3) ב LB החממה חזק ולמדוד את ה OD600 על ספקטרופוטומטרים באמצעות 1:10 דילול עם מדיה ליברות. חישוב הנפח של צורך להוסיף 1 ליטר של מדיה עבור ההתחלה OD600 של 0.1 לילה תרבות (לדוגמה, אם יתר600 של 1:10 דילול נקרא כמו 0.4, לחסן 25 מ של ה OD מדולל600 = 4.0 תרבות לילה לתוך 1 ליטר 2 x YTP ז).
- הסר את המדיה x YTP 2 ומחוממת והפתרונות D-גלוקוז החממה 37 ° C יחד עם 50 מ של תרבות ליברות. בזהירות בטכניקה סטרילי, שופכים את הפתרון D-גלוקוז לתוך התקשורת x YTP 2 (הימנעות הצדדים + בקבוקי שתייה צידניות במבוכה).
הערה: תוספת של D-גלוקוז משלים את המתכון 1 ליטר של 2 x YTPG. - שמירה על טכניקה סטרילי, לחסן את 1 ליטר של פתרון 2 x YTPG עם הכמות המתאימה של התרבות 50 מ ל כדי להתחיל את תרבות 1 ליטר-OD 0.1600. מיד למקם את תרבות 1 ליטר חוסנו 37 ° C רועד חממה-200 סל ד.
- . קח את יתר הראשון600 קריאה לאחר השעה הראשונה של צמיחה (שלב ההשהיה טיפוסי לוקח 1 h). לא לדלל את התרבות. ממשיכים את המידות600 OD כ כל 20-30 דקות עד OD600 מגיע 0.6.
- כשמגיעים OD600 = 0.6, להוסיף 1 מ"ל של 1 מ' IPTG (הריכוז הסופי בתרבות 1 ליטר = 1 מ מ) לתרבות x YTPG 2.
הערה: אינדוקציה אידיאלי OD600 היא 0.6; עם זאת, מגוון של 0.6-0.8 מקובל. אינדוקציה על ידי IPTG הוא ייצור אנדוגני של T7 RNA פולימראז (T7RNAP). - לאחר אינדוקציה, למדוד יתר600 כ כל 20-30 דקות עד שהוא מגיע 3.0.
הערה: לקרר לצנטריפוגה עד 4 ° C בתקופה זו. להכין מאגר S30 קר כמפורט המידע הנוסף. אם המאגר S30 מוכן מראש, ודא DTT לא יתווסף עד היום של שימוש. - ברגע ה OD600 מגיעה 3.0 (איור 2 א), שופכים את התרבות לתוך בקבוק קר של צנטריפוגה 1 ליטר באמבט קרח-מים. להכין מלא מים 1 ליטר צנטריפוגה בקבוק של משקל שווה כדי לשמש איזון בצנטריפוגה.
הערה: ספיגת ערכים משתנים בין כלי נגינה כלי נגינה. בעוד ה OD600 הקציר של BL21(DE3) הוא לא רגיש משתנה, מומלץ כי המשתמש להעריך ולמטב את המשתנה הזה כאמצעי לפתרון בעיות. מכשירים גדולים עלולים לגרום נמוכות יחסית קריאות OD600 לעומת מכשירים מבוססי cuvette קטנים יותר. - Centrifuge את הבקבוקים 1 ליטר 10 דקות ב- 5,000 x g ו- 10 ° C עד הצניפה תאים.
- מוזגים תגובת שיקוע, השלך אותו על פי הנהלים פסולת ביולוגית של המוסד. במקום בגדר על קרח.
- בעזרת מרית סטרילי, לגרד בגדר תא מהבקבוק צנטריפוגה, להעביר אותו צינור חרוטי קר 50 מ.
- להוסיף 30 מ של מאגר S30 קר צינור חרוטי, resuspend בגדר תא על-ידי vortexing עם צרורות קצרים (20-30 s), תקופות מנוחה (1דקות.) על הקרח עד resuspended באופן מלא עם שום גושים.
- לאחר בגדר מלאה resuspended, להשתמש עוד צינור חרוטי 50 מ עם מים כמו איזון צנטריפוגה 10 דקות ב 5000-g ו- 10 ° C (טרום מקורר עד 4 ° C).
הערה: בזאת 1סנט ממכוני 3 הנדרש כאשר קצירת התאים. - שופכים את תגובת שיקוע והשלך אותו על פי הנהלים פסולת ביולוגית של המוסד. Resuspend בגדר 20-25 מ של מאגר S30 וקר צנטריפוגה 10 דקות ב 5000 x g ו- 10 ° C (טרום מקורר עד 4 ° C).
הערה: בזאת את 2nd ממכוני 3. - שוב, שופכים את תגובת שיקוע והשלך אותו על פי הנהלים פסולת ביולוגית של המוסד. להוסיף בדיוק 30 מ של מאגר S30, מערבולת שוב כדי resuspend בגדר.
- שימוש השפופרות חרוט 3 שנשקל מראש, קר 50 מ ל ו המילוי פיפטה סרולוגית עם פיפטה סטרילי, aliquot 10 מ"ל של גלולה resuspended/S30 מאגר תערובת לתוך כל הצינורות חרוט 3.
הערה: פיצול התאים לתוך צינורות 3 אינו נדרש, אך שלב זה מתבטא כדורי תא קטן יותר (~ 1 g) לנוחות מוגברת על צעדים מאוחר יותר. - Centrifuge לכל הצינורות, באמצעות איזונים המתאימים לפי הצורך, עבור 10 דקות ב 5000-g ו- 10 ° C (טרום מקורר עד 4 ° C).
הערה: בזאת את הצעד הסופי לשטוף. - שופכים את תגובת שיקוע והשלך אותו על פי הנהלים פסולת ביולוגית של המוסד. להסיר את המאגר S30 עודף על ידי בקפידה מנגב הפנימי של צינור חרוטי, כיפה עם טישו נקי; להימנע מלגעת בגדר.
- Reweigh הצינורות על איזון האנליטי ולהקליט את המשקל צניפה הסופי על כל שפופרת.
הערה: הפרוטוקול ניתן להשהות בשלב זה. החבילות ניתן פלאש קפוא חנקן נוזלי ומאוחסנים ב-80 מעלות צלזיוס במשך שנה עד לצורך הכנת תמצית.
2. גולמי תא לחלץ הכנה - יום ד'
- להכנת תמצית, לשמור על תאים קר על קרח במהלך כל שלב. להוסיף 1 מ"ל קר S30 מאגר לכל 1g של התא המוני בגדר. ודא שכי dithiothreitol (DTT) יש כבר בתוספת למאגר S30 כדי ריכוז סופי של 2 מ מ.
הערה: לקרר את microcentrifuge עד 4 ° C בתקופה זו. - Resuspend בגדר תא על-ידי vortexing עם צרורות קצרים (20-30 s), תקופות מנוחה (1דקות.) על הקרח עד resuspended לגמרי. אם resuspension קשה, להשאיר את כדורי קרח למשך 30 דקות. להפשיר.
- העברה מ ל 1.4 של תאים resuspended לתוך צינור microfuge 1.5 mL.
- המקום המכיל 1.4 מ של תאים resuspended לתוך אמבט מים קרים בתוך שפופרת אחת של 1.5 mL. Sonicate עבור 45 s על ואחריו 59 s את מחזורי הכולל 3, עם משרעת להגדיר ב-50%. סגור, היפוך הצינורות לערבב בעדינות במהלך התקופות כבוי. בסך הכל, לספק 800-900 J של אנרגיה כל שפופרת microfuge 1.5 mL המכיל 1.4 מ של תאים resuspended (איור 3 א & 3B).
הערה: השלב זה רגיש לסוג sonicator, מודל המשמש צריך להיות מותאם אם ציוד שונה מאשר המפורטים עבור הליך זה. שתי גישות משלימות יכול לשמש כדי הסולם הסכום של תמצית שהוכנו במהלך שלב זה: צינורות microfuge 1.5 מ"ל במספר 1) יכול להיות sonicated במקביל, ו/או אחסון 2) גדולים יותר יכולים להיות sonicated חרוט צינורות (עד 15 מ"ל של תא resuspension למחזור) , קנה מידה של כמות האנרגיה מועברת כאמור תיאר 29,45. - מיד לאחר השלמת sonication, להוסיף µL 4.5 של 1 מ' DTT (שכשהם נוספים 2 מ"מ DTT) לתוך mL 1.4 של lysate, היפוך מספר פעמים כדי לערבב. מניחים את הצינורית על קרח. חזור על שלבים 2.4 ו- 2.5 עבור כל צינורות נוספים של תאים resuspended לפני שימשיך צנטריפוגה.
- דוגמאות Microcentrifuge-18,000-g ו- 4 מעלות צלזיוס למשך 10 דקות (איור 3C).
- Pipette את תגובת שיקוע לתוך צינור 1.5 מ ל microfuge. נא לא להפריע בגדר; עדיף להשאיר כמה תגובת שיקוע מאחור כדי לשמור על טוהר יותר כדי לשבש את צניפה במאמצים כדי למקסם את התשואה.
- דגירה של תגובת שיקוע מהשלב הקודם ב 250 סל ד ו- 37 מעלות צלזיוס במשך 60 דקות על ידי מקליטה הצינורות פלטפורמת חזק של החממה (זוהי התגובה נגר).
- דוגמאות Microcentrifuge ב 10,000 x g ו- 4 מעלות צלזיוס למשך 10 דקות.
- הסר את תגובת שיקוע מבלי להפריע בגדר, להעביר אותו צינור חדש. יוצרים רבים-100 µL aliquots של תמצית לאחסון.
הערה: הפרוטוקול אפשר לעצור כאן, התמצית וניתן פלאש קפוא חנקן נוזלי ומאוחסנים ב-80 מעלות צלזיוס במשך שנה עד זקוק לתגובות CFPS. לפחות 5 מחזורים ההקפאה-הפשרה ניתן עברו בלי לרעת כדי לחלץ את הפרודוקטיביות (איור 4).
3. תא ללא חלבון סינתזה אצווה תבנית תגובות
- הפשרת פתרונות A ו- B, DNA תבנית, BL21(DE3) תמצית (אם קפוא), T7RNAP, aliquot של מים ברמה המולקולרית.
הערה: ניתן למצוא CFPS התגובה תבנית מידע משלים. מתכונים פתרונות A ו- B הינם מסופקים מידע משלים , שיתאימו ריכוזים מסוים עבור ריאגנטים רבים לתמוך מערכת אנרגיה המבוסס על PANOx-SP עבור CFPS. התפקיד של כל ריאגנט מקובל וריאציה בריכוזים ריאגנט אלה שיכולים לתמוך CFPS היה נחוש50. פרוטוקול טיהור T7RNAP ניתן למצוא את המידע המשלים51. T7RNAP משלימה ניתן להגדיל את התשואות הנפחי אך אינה הכרחית אם T7RNAP הנגרמת במהלך גדילת התאים. תבנית ה-DNA פלסמיד (pJL1-sfGFP) ניתן להכין בעזרת ערכת maxiprep עם שני שוטף באמצעות המאגר שטיפת בערכה, ואחריו עיבוד דפוס DNA-ניקיון באמצעות ערכת טיהור PCR (איור 2B). תבניות DNA ליניארי יכול לשמש גם בתגובות CFPS. - תווית הסכום הדרוש של צינורות microfuge זקוק לתגובות CFPS.
הערה: ניתן לבצע תגובות בגדלים שונים של כלי, אך כלי קטן יכול להקטין נפחי חלבון התשואות (איור 2C). אולם תגובה באותו גודל הכלי שינוי קנה המידה עשויה להפחית גם את התשואות נפחי, כפונקציה של הפחתת ההחלפה חמצן, עקב ירידה ברמת פני השטח יחס נפח. בעת הגדלת נפח התגובה מעל 100 μL, מומלץ להשתמש בתחתית שטוח צלחות טוב 31,37,52. - להוסיף µL 2.2 של פתרון A, µL 2.1 של פתרון ב', 5 µL של BL21*(DE3) לחלץ, 0.24 μg של T7RNAP (16 μg/mL הסופי ריכוז), 0.24 ng של תבנית ה-DNA (16 ריכוז סופי ng/mL) ומים כדי להביא את עוצמת הקול הסופי 15 µL.
הערה: מערבולת פתרונות A ו- B לעתים קרובות במהלך התגובה ההתקנה כדי למנוע שיקוע של רכיבים ולהבטיח כי כל תגובה מקבל aliquot הומוגנית של כל פתרון. להימנע vortexing התמצית, במקום להפוך את הצינור כדי לערבב. - לאחר כל ריאגנטים נוספו התגובה, מערבבים כל שפופרת על-ידי pipetting למעלה ולמטה או בעדינות vortexing תוך הקפדה כי התערובת הסופית התגובה משולבים חרוז 15 µL בודדת בתחתית השפופרת microfuge 1.5 mL.
- מקם כל התגובה לתוך החממה 37 ° C מבלי לרעוד במשך 4 שעות, או 30 מעלות צלזיוס למשך הלילה.
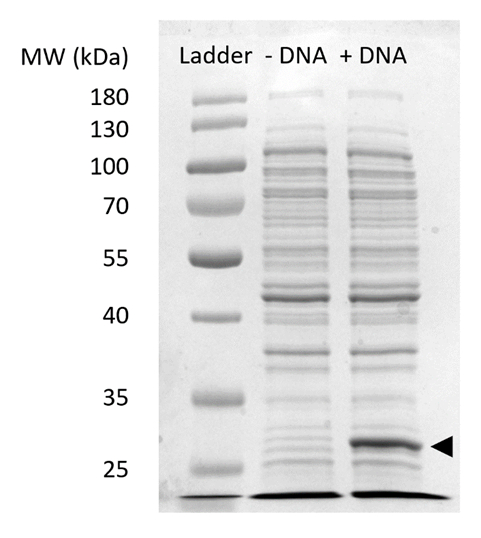
הערה: תגובות מוצלח יכול להיות איכותית העריכו באופן חזותי המבוסס על הצבע הירוק של המוצר sfGFP בתוך התערובת תגובה CFPS (דמות תלת-ממד). באפשרותך גם לאשר ביטוי של החלבון עניין מרחביות-דף (משלימה איור 2).
4. כימות של החלבון כתב, [sfGFP]
- לטעון µL 48 של 0.05 M HEPES, pH 8, לתוך כל טוב לצורך כימות (בדרך כלל מבוצע שהפקידים למחזור התגובה).
- הסר את התגובות של חממה. פיפטה למעלה ולמטה כדי לערבב כל תגובה ולאחר מכן להעביר µL 48 של 0.05 M HEPES, pH 8 2 µL של תגובה. Pipet למעלה ולמטה שוב בתוך הבאר לערבב.
- לאחר כל התגובות הם נטען, מעורב, למקם את הצלחת היטב 96 fluorometer, למדוד קרינה פלואורסצנטית את הקצה של sfGFP.
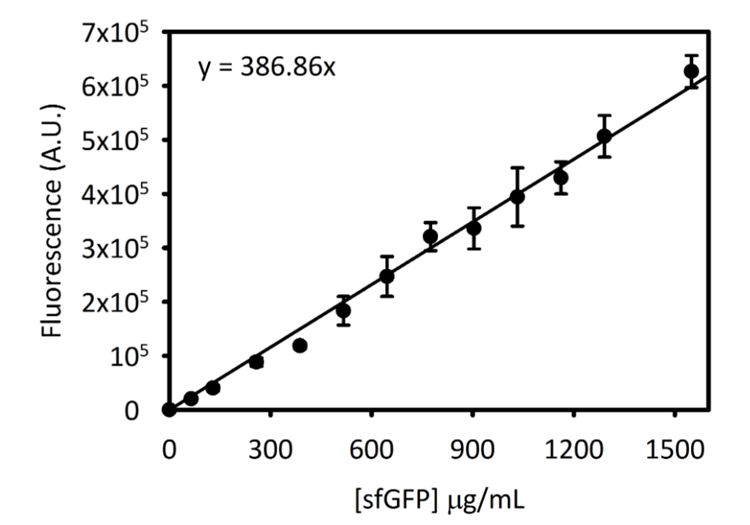
הערה: אורכי גל עירור, פליטה על כימות זריחה sfGFP הם 485 nm ו- 510 ננומטר, בהתאמה. - באמצעות עיקול רגיל שנוצרה קודם לכן, קבע את [sfGFP] מן המקראות פלורסצנטיות שהושג.
הערה: הוראות ליצירת עיקול רגיל של ריכוז sfGFP מול עוצמת קרינה פלואורסצנטית ניתנים מידע משלים (משלים איור 3). משתמשים יצטרכו להקים עיקול רגיל לכלי שלהם מאז רגישות המכשיר עשוי להשתנות.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
הוצגו מבוסס sonication e. coli תמצית פרוטוקול הכנה ניתן להשלים במשך תקופה ארבעה ימים, עם איור 1 הממחיש את התפלגות פרוצדורלי מעל כל יום. אין גמישות כדי השלבים ניתן להשלים כל יום עם נקודות משהה שונות, אבל מצאנו את זרימת עבודה זו כדי להיות היעיל ביותר לביצוע. בנוסף, לתא כדורי (שלב 1.3.18) והן בהכנה מלאה. תמצית (שלב 2.10) יציבים ב-80 מעלות צלזיוס לפחות שנה, המאפשר למשתמש ליצור מניות גדולים יותר מכל אחת כדי לשמור לשימוש בכל זמן מאוחר יותר17. לא רק היא התמצית יציבה על פני תקופות זמן ארוך, אך התמצית יכולות לעבור גם להקפיא לפחות חמישה מחזורים להפשיר ללא לאובדן משמעותי של פרודוקטיביות (איור 4). דבר זה מאפשר aliquots גדול יותר של תמצית יאוחסנו לשימושים רבים אם המקפיא שטח אחסון מוגבל. עם זאת, אנו ממליצים aliquots קטנים מרובים (~ 100 μL) של תמצית במידת האפשר.
עם כל הכנה תמצית חדשים, אנו ממליצים כי המשתמש יבצע טיטור של מגנזיום על מנת לקבוע את כמות אופטימלית מגנזיום עבור אצווה הזה של תמצית. משתמשים יכולים לכמת אצווה-כדי-אצווה ההשתנות בריכוז החלבון הכולל של התמצית תא על-ידי ברדפורד וזמינותו. עבור ביצוע גבוה יותר תמציות, שאנחנו בדרך כלל רואים ריכוז חלבון סך של 30-50 מ"ג/מ"ל, בטווח הזה יש מתאם ברור בין חלבון הכולל ריכוז וביצועים תמצית התא. לכן, אנו ממליצים למשתמשים לכוון ריכוזי מגנזיום בהתאם על מנת להבטיח כי חלבון ופונקציונליות חומצת גרעין מוגדלים עבור כל אצווה תמצית. רמות מגנזיום חשובים עבור שכפול ה-DNA תקין, תמלול ותרגום, אך רמות גבוהות מדי עלולה לפגוע בבריאותך אלה תהליכים53. כדי להדגים את התלות הזאת, אנחנו אמורה לבצע טיטור שיתוף של מגנזיום ולחלץ אמצעי האחסון כדי לקבוע את השילוב האופטימלי זה ממזער את כמות תמצית הצורך, תוך שמירה על תגובת פרודוקטיבי (איור 5). ומהניסוי הזה, אנו ממליצים להשתמש µL 5 של תמצית ו 10 מ מ ג2 + עבור תמצית עם תוכן חלבון סך של 30 מ"ג/מ"ל, על מנת לקבל מעל 1,000 μg/mL של sfGFP.
הניסיון שלנו עם CFPS גם אפשרה לנו לקבוע שלבים בתוך פרוטוקול יכולים להיות מגוונים ללא נזק פריון העבודה של המערכת, ואחרים הם חלק בלתי נפרד עבור מערכת CFPS בעלי ביצועים גבוהים. בעיקר, יתר סופית600 תא הקציר אינה משפיעה באופן משמעותי את הפלט הסופי של התגובה CFPS, תאים יתכן כבניין ובביצים לשום 2.7-4.0 OD600. זה מייצג את מעריכי בשלב מוקדם של צמיחה ריבוזום ריכוז עבור כל תא הוא הגבוה ביותר בהן המיכון translational הוא הפעיל כדי לתמוך בצמיחה מהירה. התבוננות זו מאפשרת למשתמשים גמישות לייעל את ההליכים משלהם. אנו ממליצים קציר-כ 3.0 OD600 כדי ללכוד את התאים על יתר600 קרוב עד 3.3 בבציר הזמן היא מלאה (איור 2 א). המשתנים המשפיעים על התשואות CFPS כוללות תבנית ה-DNA איכות, תגובת כלי בגודל כמויות יחסיות של תא תמצית ומגנזיום יון נוכח התגובה. . מצאנו את הדנ א איכות יש וריאציה אצווה-כדי-אצווה הבולטים. כדי לפתור בעיה זו, אנו ממליצים כי המשתמשים לטהר הדנ א באמצעות midi או מקסי הכנה, ואחריו צעד נוסף של ניקוי הדנ א גם על העמודה טיהור DNA המשמשים maxiprep, או שלאחר טיהור באמצעות ערכת ניקוי נוספים של ה-DNA. זה משפר את הפארמצבטית באיכות DNA לתגובות CFPS ותוצאות עמידים יותר ייצור החלבון (איור 2B). מיכל התגובה משפיע גם בתשואות נפח, כך ייצור חלבון של תגובה זהה setups בכמויות משתנות של כלי יכולים להיות שונים עד 40%. זה כבר תיאוריה כי בוסט כממוצע הנפחי שנצפתה כלי גדול יותר עקב שטח משטח מוגברת של תערובת התגובה, ומאפשר לחילופי חמצן טובה יותר (איור 2C), וכי אחרים יש מועלה התשואות נפח על-ידי הפעלת CFPS בתגובות צלחות שטוח התחתונה גדולים, אשר מומלץ עבור תגובות מעל 100 µL17,31,37,52.

איור 1 : ציר הזמן עבור תרבות גידול, ייצור תא תמצית, ההתקנה כימות של תגובות CFPS. המשתמש יכול ליישם פלטפורמת CFPS עבור יישומים המחקר שלהם באמצעות זרימת עבודה זו ארבעה ימים. הכנה ריאגנט מייצג את הזמן הראשי ואת עלות ההשקעה בסיבוב הראשון של הניסוי הזה, פוחתת באופן משמעותי אחרי מניות ריאגנטים נקבעו. בנוסף, תא כדורי ולחלץ תא מוכן ניתן לאחסן במשך מעל לשנה ב-80 מעלות צלזיוס, המאפשר למשתמש להתחיל על ציר הזמן-שלבים שונים עבור תוצאות מהירות יותר. המשתמש יכול גם להשהות על השלבים השונים כדי לשנות את ציר הזמן של זרימת עבודה זו. אנא לחץ כאן כדי להציג גירסה גדולה יותר של הדמות הזאת.

איור 2 : תנאים לשינוי CFPS, ההשפעות על התגובה הנפחי התשואות. א תמצית פרודוקטיביות השוואה בהתבסס על קצירת BL21(DE3) תאים על קריאות600 OD שונים. בהתבסס על מגרש זה, אנו ממליצים קציר-OD600 של 3.3 לייצר לפחות 1000 µg/mL של חלבון המטרה. תגובות בוצעו בקנה מידה μL 15 1.5 mL microfuge צינורות. B. השוואה של שני ה-DNA maxiprep שטיפת פרוטוקולים עם ובלי פוסט-טיהור דנ-אל פח האשפה. pJL1-sfGFP פלסמידים עבר של maxiprep עם אחד או שניים שוטף ואחריו ' ניקוי ' טיהור פוסט על-ידי ערכת טיהור PCR. כדי להשיג ~ 900 µg/mL ביטוי חלבון, אנו מציעים מבצע ניקיון DNA פוסט-טיהור ללא קשר למספר maxiprep שוטף. תגובות בוצעו בקנה מידה μL 15 1.5 mL microfuge צינורות. ג 15 תגובות CFPS µL לבצע בכלי שונים החל מ ל 2 צינורות microfuge 0.6 מ"ל. "מינוס" מייצג פקד שלילי שבו אין תבנית ה-DNA נוספה התגובה. כל קווי השגיאה לייצג סטיית תקן 1 של שלוש תגובות עצמאי עבור כל תנאי, שכל אחד מהם היה לכמת דולר. אנא לחץ כאן כדי להציג גירסה גדולה יותר של הדמות הזאת.

איור 3 : מפתח setups פרוצדורלי ותוצאות ליצירת פרודוקטיבי לחלץ. א ההתקנה הנכונה sonication אמבט מים קרים כדי להבטיח קירור של הדגימה בזמן חום נוצר במהלך sonication. נולד ב 1.5 mL microfuge שפופרת המכילה תאים resuspended גלולה לפני (משמאל) ו sonication פוסט (מימין). וכתוצאה מכך lysate צריך להציג גוון כהה יותר בהשוואה תא resuspended גלולה. ג. הפרדה נאותה של תגובת שיקוע, גלולה של תא lysate לאחר 18,000 x g צנטריפוגה. תגובות CFPS ד אחרי 4 שעות של דגירה ב 37 º C. 1.5 mL צינור microfuge מצד הימין (תגובה מוצלחת) מראה פלורסצנטיות גלוי של החלבון כתב sfGFP-~ 900 μg/mL. הצינור בקרה שלילית מצד השמאל, חסר תבנית ה-DNA, הדמיית לתגובה לא מוצלח, הצגת פתרון ברור עם אין קרינה פלואורסצנטית. אנא לחץ כאן כדי להציג גירסה גדולה יותר של הדמות הזאת.

איור 4 : שינוי בביטוי חלבונים יותר מ-5 מחזורים ההקפאה-הפשרה עבור CFPS לחלץ. תמצית מקריסטלים של הגידול אותו עברה חמישה להקפיא הפשרת מחזורים באמצעות חנקן נוזלי פלאש קפוא שהופעלו על ידי מפשיר על קרח. ללא שינויים משמעותיים בכל הנוגע לפרודוקטיביות תמצית לבטא sfGFP נראו מעל את 5 מחזורים ההקפאה-הפשרה. תגובות בוצעו בקנה מידה μL 15 1.5 mL microfuge צינורות. "מינוס" מייצג פקד שלילי שבו אין תבנית ה-DNA נוספה התגובה. כל קווי השגיאה לייצג סטיית תקן 1 של שלוש תגובות עצמאי עבור כל תנאי, שכל אחד מהם היה לכמת דולר. אנא לחץ כאן כדי להציג גירסה גדולה יותר של הדמות הזאת.

איור 5 : CFPS לתגובות והונגריות [מ"ג2+] חלץ כרכים לעומת [sfGFP]. [מ ג2 +] נע בין 8 מ"מ עד 14 מ מ עם גידול של 2 מ מ ולחלץ כרכים נע בין 3 µL אל µL 7 עם גידול של µL 1. מייצגת קוד צבע [sfGFP] המופק גבוהה (אדום) נמוך (סגול). כדי למקסם את היעילות ריאגנט תוך שמירה על ייצור חלבון גבוהה, אנו ממליצים על שימוש 5 µL של תמצית ו- 10 מ מ ג2+ על תמציות שיש את התוכן חלבון סה כ ~ 30 מ"ג/מ"ל, כפי שנקבע על ידי שיטת ברדפורד. נקודות המקורית כדי ליצור את העלילה מתאר היו מבוסס על הקצה של זריחה של שלוש תגובות עצמאי עבור כל תנאי, שכל אחד מהם נמדדה שהפקידים. תגובות בוצעו בקנה מידה μL 15 1.5 mL microfuge צינורות. אנא לחץ כאן כדי להציג גירסה גדולה יותר של הדמות הזאת.
משלים איור 1: עלות לכל מיקרוגרם של חלבון המיוצר ו לכל microliter של תגובה הפלטפורמות סינתזת חלבון נטול תאים שש. הפלטפורמה שלנו מושווה לענייני חוץ חלבון חינם תא שונים חמש, סינתזה ערכות/פלטפורמות עם פרודוקטיביות משתנה ותמחור. פלטפורמת CFPS שלנו sonication מבוסס היא חסכונית יותר בשני $/ µg של חלבון ו $/ µL של תגובה מאשר רוב ערכות ומסחר מספקת הקלות של ערכת ההתקנה התגובה, תוך עלות נותרת להשוות פלטפורמות אחרות-CFPS אקדמי. אנא לחץ כאן כדי להציג גירסה גדולה יותר של הדמות הזאת.
משלים איור 2: מרחביות-דף של ביטוי sfGFP CFPS. ללא תא סינתזת חלבון המגיבה (+ DNA) וללא (-DNA) תבנית ה-DNA sfGFP היו מריצים של 12% SDSPAGE ג'ל כדי להדגים את הביטוי של sfGFP נצפו על 27 kDa (חץ שחור). נעשה שימוש בטכניקות מסורתיות מרחביות-דף. כל מדגם מועמסים על הג'ל כלולים 18 µg של חלבון הכולל בהתבסס על ברדפורד כימות וזמינותו של סך החלבון בתא לחלץ. על סמך מדידות עוצמת קרינה פלואורסצנטית, עיקול רגיל שלנו, אנחנו מעריכים "+ DNA" ליין מכיל µg 0.42 של sfGFP. כדי להשיג דגימות אלה, CFPS תגובות היו לפעול בקנה מידה 15 µL 1.5 mL microfuge צינורות בהפקת בקנה אחד עם איור 3Cהנפחי התשואות. אנא לחץ כאן כדי להציג גירסה גדולה יותר של הדמות הזאת.
משלים איור 3: עקומת סטנדרטי לכל sfGFP 5 Cytation. עקום זה נקבע באמצעות השיטות שפורטו לעיל. כל הנתונים שנאספו עבור כתב יד זה הוסב הקצה של קרינה פלואורסצנטית בהנחה [sfGFP] µg/mL באמצעות עיקול רגיל זה. אנא לחץ כאן כדי להציג גירסה גדולה יותר של הדמות הזאת.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
סינתזה של חלבון נטול תא התפתחה טכנולוגיה חזקה ומאפשרת למגוון רחב של יישומים החל biomanufacturing וכלה שטנץ מהירה של מערכות ביוכימיות. רוחב היריעה של יישומים נתמך על ידי היכולת לפקח, לתמרן, להגדיל מכונות הסלולר בזמן אמת. למרות ההשפעה המתרחב של טכנולוגיה זו פלטפורמה, נותרה רחבה הסתגלות איטית עקב הדקויות הטכניות ביישום של השיטות. דרך המאמץ הזה, אנו שואפים לספק פשטות ובהירות להקמת טכנולוגיה זו במעבדות חדש. לצורך כך, הפרוטוקול שלנו עבור e. coli-פלטפורמה סינתזת חלבון נטול תא מבוסס יכולה להיות מושגת תוך זמן האתחול של ארבעה ימים על ידי מעבדה שהוכשרו הלא-מומחים, כגון סטודנטים לתואר ראשון (איור 1). בנוסף, ברגע מלאי של ריאגנטים, תמצית מיוצרים, תגובות עוקבות של אצווה CFPS יכול להיות להגדיר, מתפשט, לכמת ברק 5 שעות. יחיד, צמיחת תאים 1 ליטר יכול לגרום מספיק תמצית לתגובות CFPS ארבע מאות 15 µL, בזמן ההכנות אצווה אחת של ריאגנטים אחרים ללא תא יכול לתמוך אלפי תגובות. גם וניתן לשנותם ריאגנט ההכנות אם מלאי גדול עוד יותר יש צורך. התגובות CFPS ניתן להגדיר באופן תפוקה גבוהה, באמצעות צלחת 96-ובכן או המבחנות לבדיקת מגוון תנאים במקביל. התשואות הנפחי יקטן בעת שימוש בכלי קטן יותר כפי שניתן לראות באיור 2C. תגובות CFPS גם ניתנים לשינוי מ microliters עשרות התגובה סה כ נפח מיליליטר על מנת להגדיל את התשואה הכוללת חלבון עבור תנאי יחיד. כאשר קנה המידה הווליום, השיקול המרכזי הוא כי התגובה הנפחי התשואות להקטין כמו יחס נפח באיזור פני השטח של התגובה מקטין37,52. על מנת הסולם למעלה תוך שמירה על התשואות נפח דומה לביטוי חלבון, משתמשים צריכים לפצל שעוצמת התגובה רבים כלי התגובה ו/או להגדיל את כלי הקיבול. עבור התגובה סולמות החל µL 15 - 100 µL בכרך, תגובות µL 15 רבות במקביל מומלץ. לתגובות העולה על 100 µL בכרך, שטוח התחתונה. ובכן 24 צלחות מומלצים, ובכן 12 צלחות מומלצים עבור אמצעי אחסון התגובה העולה על 600 µL. זיווגים כאלה של כלי ואמצעי תגובה לספק עקביות של התגובה הנפחי התשואות על הסולם17,31,37,52. אולם שינוי קנה המידה מעבר אמצעי אחסון אלה יכול להתבצע על ידי ניצול בארות מרובים של הצלחת במקביל. באמצעות תבנית זו, התגובה וניתן לשנותם הנפח הכולל של מעל 10 מ. אופטימיזציה של השילוב כלי אחסון-תגובה תגובה יכול לתמוך ביישומים של biomanufacturing מבלי להקריב את הפרודוקטיביות של התגובה.
בעת ביצוע פרוטוקול זה, ישנם מספר שיקולים מרכזיים שמשפיעים על התגובה הנפחי התשואות, כמו גם אינדיקטורים המשויך שתפקודו תמצית. על מנת להבטיח פירוק נכונה וכדי למנוע דנטורציה של מכונות תעתיק/תרגום פונקציונלי, חשוב להפחתת החום המופק במהלך פירוק. לטבול את resuspension תא באמבט קרח מים במהלך sonication להתפוגג במהירות חום במהלך sonication (איור 3 א). מחוון של פירוק התא יעילה היא הופעתם של מראה כהה יותר של התא lysate לעומת דגימות pre-sonicated (איור 3B). להשגת גמישות למשתמש, sonicator ואת המכשיר שמוצג באיור 3A היא יכולת הסתגלות מגוון של אמצעי אחסון מ 100 µL 15 מ של תאים resuspended. כדי לעשות זאת, המשתמש יכול לשנות את המספר של גאול עבור פירוק של אמצעי האחסון הרצויה של תאים. בנוסף, ניתן להכין כמויות גדולות יותר של תמצית דרך שתי גישות משלימות. משתמשים יכולים sonicate צינורות מרובות במקביל, ו/או sonicate אחסון גדולים יותר של התא resuspension, קנה מידה של כמות האנרגיה באופן יחסי עם נפח כמו שתואר לעיל29,45. צעד נוסף המציין תמצית איכות היא הצעד צנטריפוגה בעקבות פירוק התא. פוסט פירוק התא, אנו ממליצים צנטריפוגה ב g x 18,000 לספק חלוקה ברורה בין תגובת שיקוע (מכונות תרגום/תמלול, מפוצלים דנ א גנומי אשר כבר לא מתפקד תרגום/תמלול תבנית), את (גלולה לא רצויות, מרכיבי התא כגון קרום התא, זירז חלבונים) (איור 3C). מצאנו את צנטריפוגה-18,000 x g משפר את ההפרדה, וכתוצאה מכך הפארמצבטית משופרת לעומת ספינים במהירויות נמוכות כגון g 12,000 x. לנוחיותכם, אנו ממליצים להשתמש שולחני בקירור צנטריפוגה, ולתסכולים מינימום של 12,000 x g. שלב זה מבוצע בדרך כלל גם ב- g x 30,000, אשר יש לקחת בחשבון אם הציוד המתאים הוא זמין54,55,56,57,58, 59 , 60. תמצית הביצועים אינה מושפעת צנטריפוגה מהירויות בשלב זה בהתחשב בכך מושגת הפרדה נאותה. בעת הסרת את תגובת שיקוע הרצויים, מומלץ להימנע כל חומר מעונן הקיימים הגבול בין תגובת שיקוע צניפה מאז זיהום זה יקטין את הפרודוקטיביות של התמצית. מכוון טוהר תגובת שיקוע תוצאות תמציות פרודוקטיבי יותר, שווה כמות מופחתת של החילוץ שהתקבל עבור משתמשים חדשים.
חשוב שימו לב: בעוד השיטות הוצגו לשחזור, יכולה להתבצע על ידי מדענים בעלי מומחיות מינימלי, יכול להיות אצווה-כדי-אצווה, וריאציה התגובה-כדי-תגובה. זה אולי ניתן לייחס וריאציה בהרכב פרוטיאומיה מבנית lysate פוסט-sonication61. ההשתנות אצווה-כדי-אצווה זה הבחנו נחלש באופן כללי על תוספי עם T7RNAP ואופטימיזציה של ריכוזי מגנזיום. תוספת אקסוגני של T7RNAP נפוצה בקרב CFPS תגובות כדי לתמוך ביטוי חלבון אופטימלי, ומוצאים לכך שיש שני מקורות T7RNAP - אנדוגני הביטוי BL21*(DE3), את T7RNAP משלימה את ריכוז הסופי של µg 16/mL - משפר אצווה-כדי-אצווה הפארמצבטית עבור משתמשים חדשים45,46. עם ניסיון, משתמשים באפשרותך לשנות את הניסויים שלהם כדי לנצל רק מקור אחד של T7RNAP, אם רצונך בכך. כימות של תכולת החלבון הכולל של אוסף חדש של תמצית וכוונון המתאים מ ג2 + ריכוז עשוי גם לסייע לצמצם אצווה-כדי-אצווה וריאציה של התשואות ביטוי חלבון הנפחי. וריאציות בביטוי חלבון יכול להיות גם עקב הבדלי גודל ומבנה של החלבון עניין, השימוש codon הגן אתר האיגוד המקביל שלו ריבוזום של הגן של עניין, כמו גם את הסוג בביטוי המשמש וקטור62 ,63. מסיבות אלו, חלבונים מסוימים עשוי לבטא לא כמו גם מודל חלבון sfGFP, וכתוצאה מכך צמצום נפחי היבול של תגובות CFPS.
מגבלות של הטכניקה CFPS הציג לכלול יתכן שלא מתאימים ישירות על כל היישומים של התא-חינם, כגון הנדסה מטבולית, כוונון של תנאי ביטוי, בלי שינויים נוספים לפרוטוקולים. עם זאת, אנו מאמינים כי פרוטוקול זה לספק בסיס להקמת פלטפורמת CFPS במעבדות חדשים, שאינם מומחים לספק את היכולת ליישם תגובות ללא תא מבוא במעבדות שלהם. לאחר היישום הראשוני, חוקרים יכולים להתנסות פלטפורמת שינויים משלהם עבור יישומים ספציפיים יותר בהתבסס על ספרות אחרים בתחום.
CFPS פלטפורמה עלויות $0.021/µg החלבון (למעט עלות עבודה וציוד), ביצוע המערכת שלנו תחרותיים עם ערכות מסחרי מבלי להתפשר על נוחות ההתקנה התגובה. הערכות של השוואתי עלויות µL של תגובה מציגים מגמות דומות (המשלימים איור 1). אנו מעריכים עלויות הפעלה להיות ~ 4,500 דולר עבור כל ריאגנטים, של נוספים $3,200 עבור ציוד מיוחד, כגון sonicator. שעות אדם כדי להשלים הליך זה מוערך לעלות ~ 26 h להכנה ריאגנט כל מן הקרקע. עם זאת, ברגע היה מוכן מלאי גדול של ריאגנטים, הדרישות לעבודה להפחית באופן משמעותי. בנוסף, כמו ניסיון עם פלטפורמת נרכשת, אנו ממליצים על שינוי קנה מידה את גודל הגידול תא, לחלץ הכנה, הכנה ריאגנט כדי למקסם את היעילות זמן. לאור עלויות הפעלה, אנו ממליצים על פלטפורמת CFPS עבור יישומים בביולוגיה סינתטית, המאמצים תפוקה גבוהה, והם תנאי ביטוי חלבון תואם פלטפורמות ביטוי חלבון המסורתי בשל להתנגש עם התא ביוכימיה, הכדאיות אילוצים. במקרים מיוחדים אלה שבו הטכניקה הרצויה מופעלת על-ידי הפלטפורמה CFPS, עלות גדולה יותר CFPS על ביטוי ויוו היא מוצדקת.
המשך הפיתוח של פלטפורמת CFPS סביר לספק השירות רחבה יותר למאמצים ביוטכנולוגיה כגון ההנדסה מטבולית של מסלולים אנזימטיות, ייצור ואפיון של חלבונים פתירות באופן מסורתי, חומצת אמינו במימדים לא התאגדות ביטוי חלבון לא טבעי, רפואה מרובדת ייצור ו להתרחב מעבר המעבדה לכיתה עבור גזע החינוך64,65,66. מאמצים אלה ייתמכו עוד יותר על ידי המאמצים המתמשכים של אפיון מפורט של פלטפורמת CFPS. הבנה טובה יותר של ההרכב של התמצית תא יוביל עידון המשך לכיוון התשואות תגובה משופרים וגמישות, התגובה תנאים61,67,68.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
המחברים מצהירים כי יש להם אין מתחרים אינטרסים כלכליים או אחרים ניגודי עניינים.
Acknowledgments
מחברים רוצה להכיר ד ר ג'ניפר VanderKelen, אנדריאה Laubscher, ואת טוני Turretto לתמיכה טכנית, ווסלי קאו, ליין ויליאמס ו כריסטופר Hight לדיונים מועיל. המחברים גם לאשר מימון תמיכה מן ביל ו לינדה פרוסט קרן, מרכז עבור יישומים של ביוטכנולוגיה שברון ביוטכנולוגיה חלה הקרן למחקר גרנט, קאל פולי מחקר, Scholarly ו תוכנית גרנט פעילות יצירתית (RSCA 2017), הקרן הלאומית למדע (NSF-1708919). MZL מאשר מענק בוגר את המדינה אוניברסיטת קליפורניה. MCJ מכיר את הצבא מחקר במשרד W911NF-16-1-0372, הלאומית למדע מעניקה MCB-1413563 ו MCB-1716766, את חיל האוויר מחקר מעבדה מרכז מצוינות גרנט FA8650-15-2-5518, המענק סוכנות ההגנה צמצום האיום HDTRA1-15-10052/P00001, דוד ואת ולוסיל פקארד קרן, לתוכנית המורה-מלומד קמיל דרייפוס, המחלקה של אנרגיה בער גרנט דה-SC0018249, תוכנית המדע האנושי הגבולות (RGP0015/2017), המענק ETOP המכון הגנום המאוחד DOE, ו שיקגו ביו. האיחוד עם תמיכה של הכספים סרל ב שיקגו הקהילה לסמוך על התמיכה.
Materials
| Name | Company | Catalog Number | Comments |
| Luria Broth | ThermoFisher | 12795027 | |
| Tryptone | Fisher Bioreagents | 73049-73-7 | |
| Yeast Extract | Fisher Bioreagents | 1/2/8013 | |
| NaCl | Sigma-Aldrich | S3014-1KG | |
| Potassium Phosphate Dibasic | Sigma-Aldrich | 60353-250G | |
| Potassium Phosphate Monobasic | Sigma-Aldrich | P9791-500G | |
| D-Glucose | Sigma-Aldrich | G8270-1KG | |
| KOH | Sigma-Aldrich | P5958-500G | |
| IPTG | Sigma-Aldrich | I6758-1G | |
| Mg(OAc)2 | Sigma-Aldrich | M5661-250G | |
| K(OAc) | Sigma-Aldrich | P1190-1KG | |
| Tris(OAc) | Sigma-Aldrich | T6066-500G | |
| DTT | ThermoFisher | 15508013 | |
| tRNA | Sigma-Aldrich | 10109541001 | |
| Folinic Acid | Sigma-Aldrich | F7878-100MG | |
| NTPs | ThermoFisher | R0481 | |
| Oxalic Acid | Sigma-Aldrich | P0963-100G | |
| NAD | Sigma-Aldrich | N8535-15VL | |
| CoA | Sigma-Aldrich | C3144-25MG | |
| PEP | Sigma-Aldrich | 860077-250MG | |
| K(Glu) | Sigma-Aldrich | G1501-500G | |
| NH4(Glu) | MP Biomedicals | 02180595.1 | |
| Mg(Glu)2 | Sigma-Aldrich | 49605-250G | |
| Spermidine | Sigma-Aldrich | S0266-5G | |
| Putrescine | Sigma-Aldrich | D13208-25G | |
| HEPES | ThermoFisher | 11344041 | |
| Molecular Grade Water | Sigma-Aldrich | 7732-18-5 | |
| L-Aspartic Acid | Sigma-Aldrich | A7219-100G | |
| L-Valine | Sigma-Aldrich | V0500-25G | |
| L-Tryptophan | Sigma-Aldrich | T0254-25G | |
| L-Phenylalanine | Sigma-Aldrich | P2126-100G | |
| L-Isoleucine | Sigma-Aldrich | I2752-25G | |
| L-Leucine | Sigma-Aldrich | L8000-25G | |
| L-Cysteine | Sigma-Aldrich | C7352-25G | |
| L-Methionine | Sigma-Aldrich | M9625-25G | |
| L-Alanine | Sigma-Aldrich | A7627-100G | |
| L-Arginine | Sigma-Aldrich | A8094-25G | |
| L-Asparagine | Sigma-Aldrich | A0884-25G | |
| Glycine | Sigma-Aldrich | G7126-100G | |
| L-Glutamine | Sigma-Aldrich | G3126-250G | |
| L-Histadine | Sigma-Aldrich | H8000-25G | |
| L-Lysine | Sigma-Aldrich | L5501-25G | |
| L-Proline | Sigma-Aldrich | P0380-100G | |
| L-Serine | Sigma-Aldrich | S4500-100G | |
| L-Threonine | Sigma-Aldrich | T8625-25G | |
| L-Tyrosine | Sigma-Aldrich | T3754-100G | |
| Fisherbrand Premium Microcentrifuge Tubes: 2.0 mL | Fisher Scientific | 05-408-138 | |
| Fisherbrand Premium Microcentrifuge Tubes: 1.5 mL | Fisher Scientific | 05-408-129 | |
| Fisherbrand Premium Microcentrifuge Tubes: 0.6 mL | Fisher Scientific | 05-408-120 | |
| PureLink HiPure Plasmid Prep Kit | ThermoFisher | K210007 | |
| Ultrasonic Processor | QSonica | Q125-230V/50Hz | 3.175 mm diameter probe |
| Avanti J-E Centrifuge | Beckman Coulter | 369001 | |
| JLA-8.1000 Rotor | Beckman Coulter | 366754 | |
| 1L Centrifuge Tube | Beckman Coulter | A99028 | |
| Tunair 2.5L Baffeled Shake Flask | Sigma-Aldrich | Z710822 | |
| Microfuge 20 | Beckman Coulter | B30134 | |
| New Brunswick Innova 42/42R Incubator | Eppendorf | M1335-0000 | |
| Cytation 5 | BioTek | ||
| Strep-Tactin XT Starter Kit | IBA | 2-4998-000 | |
| pJL1-sfGFP | Addgene | 69496 | |
| BL21(DE3) | New England BioLabs |
References
- Jiang, L., Zhao, J., Lian, J., Xu, Z. Cell-free protein synthesis enabled rapid prototyping for metabolic engineering and synthetic biology. Synthetic and Systems Biotechnology. 3 (2), 90-96 (2018).
- Carlson, E. D., Gan, R., Hodgman, C. E., Jewett, M. C. Cell-free protein synthesis: Applications come of age. Biotechnology Advances. 30 (5), 1185-1194 (2012).
- Watanabe, M. Cell-Free Protein Synthesis for Structure Determination by X-ray Crystallography. Methods in molecular biology (Clifton, N.J). 607, 149-160 (2010).
- Martemyanov, K. A., Shirokov, V. A., Kurnasov, O. V., Gudkov, A. T., Spirin, A. S. Cell-Free Production of Biologically Active Polypeptides: Application to the Synthesis of Antibacterial Peptide Cecropin. Protein Expression and Purification. 21 (3), 456-461 (2001).
- Renesto, P., Raoult, D. From genes to proteins: in vitro expression of rickettsial proteins. Annals of the New York Academy of Sciences. 990, 642-652 (2003).
- Xu, Z., Chen, H., Yin, X., Xu, N., Cen, P. High-Level Expression of Soluble Human b-Defensin-2 Fused With Green Fluorescent Protein in Escherichia coli Cell-Free System. Applied Biochemistry and Biotechnology. 127 (1), 053-062 (2005).
- Baumann, A. In-situ observation of membrane protein folding during cell-free expression. PLoS ONE. 11 (3), 1-15 (2016).
- Wang, Y., Percival, Y. H. P. Cell-free protein synthesis energized by slowly-metabolized maltodextrin. BMC Biotechnology. 9, 1-8 (2009).
- Whittaker, J. W. Cell-free protein synthesis: the state of the art. Biotechnology Letters. 35 (2), 143-152 (2013).
- Martin, R. W. Cell-free protein synthesis from genomically recoded bacteria enables multisite incorporation of noncanonical amino acids. Nature Communications. 9 (1), 1203 (2018).
- Kwon, Y. C., Song, J. K., Kim, D. M. Cloning-Independent Expression and Screening of Enzymes Using Cell-Free Protein Synthesis Systems. Methods in Molecular Biology. 1118, 97-108 (2014).
- Chappell, J., Jensen, K., Freemont, P. S. Validation of an entirely in vitro approach for rapid prototyping of DNA regulatory elements for synthetic biology. Nucleic Acids Research. 41 (5), 3471-3481 (2013).
- Takahashi, M. K. Characterizing and prototyping genetic networks with cell-free transcription-translation reactions. Methods. 86, 60-72 (2015).
- Karim, A. S., Jewett, M. C. A cell-free framework for rapid biosynthetic pathway prototyping and enzyme discovery. Metabolic Engineering. 36, 116-126 (2016).
- Dudley, Q. M., Anderson, K. C., Jewett, M. C. Cell-Free Mixing of Escherichia coli Crude Extracts to Prototype and Rationally Engineer High-Titer Mevalonate Synthesis. ACS Synthetic Biology. 5 (12), 1578-1588 (2016).
- Pardee, K. Paper-based synthetic gene networks. Cell. 159 (4), 940-954 (2014).
- Zawada, J. F. Microscale to manufacturing scale-up of cell-free cytokine production-a new approach for shortening protein production development timelines. Biotechnology and Bioengineering. 108 (7), 1570-1578 (2011).
- Sullivan, C. J. A cell-free expression and purification process for rapid production of protein biologics. Biotechnology Journal. 11 (2), 238-248 (2016).
- Li, J. Cell-free protein synthesis enables high yielding synthesis of an active multicopper oxidase. Biotechnology Journal. 11 (2), 212-218 (2016).
- Heinzelman, P., Schoborg, J. A., Jewett, M. C. pH responsive granulocyte colony-stimulating factor variants with implications for treating Alzheimer's disease and other central nervous system disorders. Protein Engineering Design and Selection. 28 (10), 481-489 (2015).
- Pardee, K. Low-Cost Detection of Zika Virus Using Programmable Biomolecular Components. Cell. 165 (5), 1255-1266 (2016).
- Slomovic, S., Pardee, K., Collins, J. J. Synthetic biology devices for in vitro and in vivo diagnostics. Proceedings of the National Academy of Sciences. 112 (47), 14429-14435 (2015).
- Gootenberg, J. S. Nucleic acid detection with CRISPR-Cas13a/C2c2. Science. 356 (6336), 438-442 (2017).
- Pardee, K. Portable, On-Demand Biomolecular Manufacturing. Cell. 167 (1), 248-259 (2016).
- Karig, D. K., Bessling, S., Thielen, P., Zhang, S., Wolfe, J. Preservation of protein expression systems at elevated temperatures for portable therapeutic production. Journal of the Royal Society, Interface. 14 (129), (2017).
- Smith, M. T., Berkheimer, S. D., Werner, C. J., Bundy, B. C. Lyophilized Escherichia coli-based cell-free systems for robust, high-density, long-term storage. BioTechniques. 56 (4), 186-193 (2014).
- Hunt, J. P., Yang, S. O., Wilding, K. M., Bundy, B. C. The growing impact of lyophilized cell-free protein expression systems. Bioengineered. 8 (4), 325-330 (2017).
- Stark, J. C. BioBitsTM Bright: A fluorescent synthetic biology education kit. Science Advances. 4 (8), (2018).
- Huang, A. BioBitsTM Explorer: A modular synthetic biology education kit. Science Advances. 4 (8), (2018).
- Zemella, A., Thoring, L., Hoffmeister, C., Kubick, S. Cell-Free Protein Synthesis: Pros and Cons of Prokaryotic and Eukaryotic Systems. ChemBioChem. 16 (17), 2420-2431 (2015).
- Oza, J. P. Robust production of recombinant phosphoproteins using cell-free protein synthesis. Nature Communications. 6 (1), 8168 (2015).
- Zemella, A. Cell-free protein synthesis as a novel tool for directed glycoengineering of active erythropoietin. Scientific Reports. 8 (1), 8514 (2018).
- Jaroentomeechai, T. Single-pot glycoprotein biosynthesis using a cell-free transcription-translation system enriched with glycosylation machinery. Nature Communications. 9 (1), 2686 (2018).
- Kightlinger, W. Design of glycosylation sites by rapid synthesis and analysis of glycosyltransferases. Nature Chemical Biology. 14 (6), 627-635 (2018).
- Schoborg, J. A. A cell-free platform for rapid synthesis and testing of active oligosaccharyltransferases. Biotechnology and Bioengineering. 115 (3), 739-750 (2018).
- Chekulayeva, M. N., Kurnasov, O. V., Shirokov, V. A., Spirin, A. S. Continuous-Exchange Cell-Free Protein-Synthesizing System: Synthesis of HIV-1 Antigen Nef. Biochemical and Biophysical Research Communications. 280 (3), 914-917 (2001).
- Hong, S. H. Improving Cell-Free Protein Synthesis through Genome Engineering of Escherichia coli Lacking Release Factor 1. ChemBioChem. 16 (5), 844-853 (2015).
- Endo, Y., Otsuzuki, S., Ito, K., Miura, K. Production of an enzymatic active protein using a continuous flow cell-free translation system. Journal of Biotechnology. 25 (3), 221-230 (1992).
- Volyanik, E. V., Dalley, A., Mckay, I. A., Leigh, I., Williams, N. S., Bustin, S. A. Synthesis of Preparative Amounts of Biologically Active Interleukin-6 Using a Continuous-Flow Cell-Free Translation System. Analytical Biochemistry. 214 (1), 289-294 (1993).
- Martin, G. A., Kawaguchi, R., Lam, Y., DeGiovanni, A., Fukushima, M., Mutter, W. High-yield, in vitro protein expression using a continuous-exchange, coupled transcription/ translation system. BioTechniques. 31 (4), 948-950 (2001).
- Stech, M., Quast, R. B., Sachse, R., Schulze, C., Wüstenhagen, D. A., Kubick, S. A Continuous-Exchange Cell-Free Protein Synthesis System Based on Extracts from Cultured Insect Cells. PLoS ONE. 9 (5), e96635 (2014).
- Quast, R. B., Sonnabend, A., Stech, M., Wüstenhagen, D. A., Kubick, S. High-yield cell-free synthesis of human EGFR by IRES-mediated protein translation in a continuous exchange cell-free reaction format. Scientific Reports. 6 (1), 30399 (2016).
- Thoring, L., Dondapati, S. K., Stech, M., Wüstenhagen, D. A., Kubick, S. High-yield production of "difficult-to-express" proteins in a continuous exchange cell-free system based on CHO cell lysates. Scientific Reports. 7 (1), 11710 (2017).
- Hoffmann, M., Nemetz, C., Madin, K., Buchberger, B. Rapid translation system: A novel cell-free way from gene to protein. Biotechnology annual review. 10, 1-30 (2004).
- Kwon, Y. C., Jewett, M. C. High-throughput preparation methods of crude extract for robust cell-free protein synthesis. Scientific Reports. 5 (1), 8663 (2015).
- Katsura, K. A reproducible and scalable procedure for preparing bacterial extracts for cell-free protein synthesis. Journal of Biochemistry. 162 (June), 357-369 (2017).
- Fujiwara, K., Doi, N. Biochemical preparation of cell extract for cell-free protein synthesis without physical disruption. PLoS ONE. 11 (4), 1-15 (2016).
- Shrestha, P., Holland, T. M., Bundy, B. C. Streamlined extract preparation for Escherichia coli-based cell-free protein synthesis by sonication or bead vortex mixing. BioTechniques. 53 (3), 163-174 (2012).
- Krinsky, N. A Simple and Rapid Method for Preparing a Cell-Free Bacterial Lysate for Protein Synthesis. PLoS ONE. 11 (10), (2016).
- Jewett, M. C., Swartz, J. R. Mimicking the Escherichia coli cytoplasmic environment activates long-lived and efficient cell-free protein synthesis. Biotechnology and Bioengineering. 86 (1), 19-26 (2004).
- Swartz, J. R., Jewett, M. C., Woodrow, K. A. Cell-Free Protein Synthesis With Prokaryotic Combined Transcription-Translation. Recombinant Gene Expression. (267), 169-182 (2004).
- Voloshin, A. M., Swartz, J. R. Efficient and scalable method for scaling up cell free protein synthesis in batch mode. Biotechnology and Bioengineering. 91 (4), 516-521 (2005).
- Vernon, W. B. The role of magnesium in nucleic-acid and protein metabolism. Magnesium. 7 (5-6), 234-248 (1988).
- Pratt, J. M. Transcription and Translation: A Practical Approach. , IRL Press. New York. (1984).
- Kim, D. M., Kigawa, T., Choi, C. Y., Yokoyama, S. A Highly Efficient Cell-Free Protein Synthesis System from Escherichia coli. European Journal of Biochemistry. 239 (3), 881-886 (1996).
- Shin, J., Noireaux, V. Efficient cell-free expression with the endogenous E. Coli RNA polymerase and sigma factor 70. Journal of Biological Engineering. 4 (1), 8 (2010).
- Zubay, G. In Vitro Synthesis of Protein in Microbial Systems. Annual Review of Genetics. 7 (1), 267-287 (1973).
- Kigawa, T. Preparation of Escherichia coli cell extract for highly productive cell-free protein expression. Journal of Structural and Functional Genomics. 5 (1/2), 63-68 (2004).
- Liu, D. V., Zawada, J. F., Swartz, J. R. Streamlining Escherichia Coli S30 Extract Preparation for Economical Cell-Free Protein Synthesis. Biotechnology Progress. 21 (2), 460-465 (2008).
- Yang, W. C., Patel, K. G., Wong, H. E., Swartz, J. R. Simplifying and streamlining Escherichia coli-based cell-free protein synthesis. Biotechnology Progress. 28 (2), 413-420 (2012).
- Foshag, D. The E. coli S30 lysate proteome: A prototype for cell-free protein production. New Biotechnology. 40 (Pt B), 245-260 (2018).
- Caschera, F. Bacterial cell-free expression technology to in vitro systems engineering and optimization. Synthetic and Systems Biotechnology. 2 (2), 97-104 (2017).
- Chizzolini, F., Forlin, M., Yeh Martín, N., Berloffa, G., Cecchi, D., Mansy, S. S. Cell-Free Translation Is More Variable than Transcription. ACS Synthetic Biology. 6 (4), 638-647 (2017).
- Hong, S. H., Kwon, Y. C., Jewett, M. C. Non-standard amino acid incorporation into proteins using Escherichia coli cell-free protein synthesis. Frontiers in Chemistry. 2, 34 (2014).
- Sawasaki, T., Ogasawara, T., Morishita, R., Endo, Y. A cell-free protein synthesis system for high-throughput proteomics. Proceedings of the National Academy of Sciences of the United States of America. 99 (23), 14652-14657 (2002).
- Dudley, Q. M., Karim, A. S., Jewett, M. C. Cell-free metabolic engineering: Biomanufacturing beyond the cell. Biotechnology Journal. 10 (1), 69-82 (2015).
- Garcia, D. C. Elucidating the potential of crude cell extracts for producing pyruvate from glucose. Synthetic Biology. 3 (1), 1-9 (2018).
- Hurst, G. B. Proteomics-Based Tools for Evaluation of Cell-Free Protein Synthesis. Analytical Chemistry. 89 (21), 11443-11451 (2017).

{kind=link}
{kind=link}
{kind=link}