

Summary
이 프로토콜 세부 단계, 비용, 그리고 대장균을 생성 하는 데 필요한 장비-셀 추출 물 및 단백질 합성 반응을 시험관에서 4 일 안에 구현. 광범위 한 응용 프로그램에 대 한이 플랫폼의 유연한 특성을 활용 하 여 우리는 적응 되 고 최적화 될 수 있는 반응 조건 논의.
Abstract
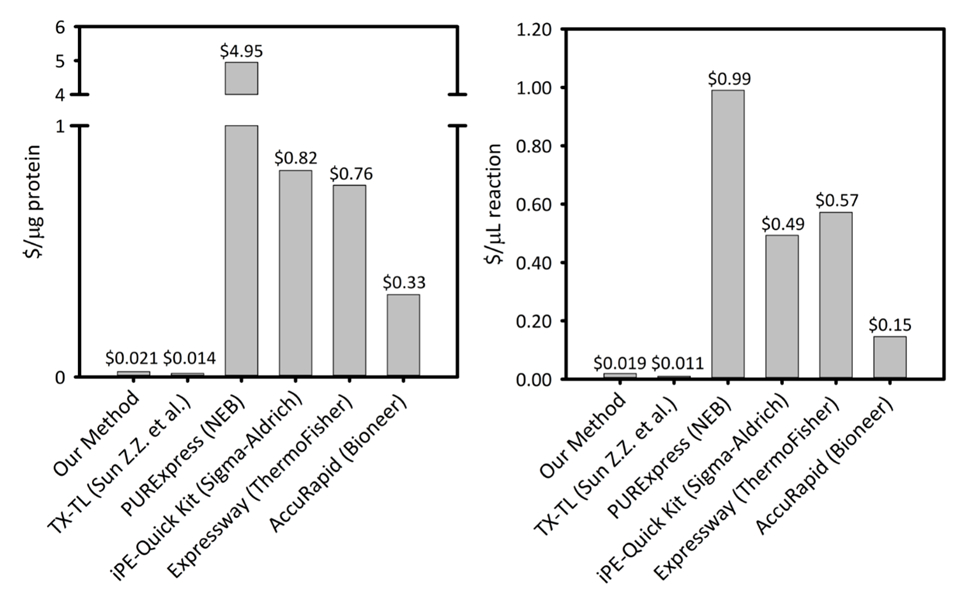
지난 50 년 동안 셀 무료 단백질 합성 (CFPS)는 시험관 내에서 셀의 transcriptional 변환 용량을 활용 하는 강력한 기술으로 떠오르고 있다. 고, 세포의 생존 능력을 유지 하는 필요를 만듦으로써 세포 장벽을 제거 함으로써, CFPS 되었습니다 전통적으로 도전 단백질의 biomanufacturing에 신흥 응용 프로그램 뿐만 아니라 응용 프로그램에 기초에 대 한 신속한 프로토 타입 대사 공학 및 기능 유전체학 대장균을 구현 하기 위한 우리의 방법-CFPS 플랫폼을 기반으로 새 사용자가 이러한 응용 프로그램의 대부분을 액세스할 수 허용. 여기, 우리는 풍부한 미디어, 당황 하 고 플라스 크와 가변 쥡니다 기반 세포 세포의 용 해의 재현 방법을 통해 추출 준비 하는 방법을 설명 합니다. 그 적절 한 시 약 주식 미리 준비 된이 추출 다음 단백질 발현 데이터 분석, 실험적인 체제에서 단지 5 h에서 슈퍼 폴더 녹색 형광 단백질 (sfGFP)의 900 µ g/mL 이상의 생산 능력에 대 한 사용할 수 있습니다. 시 약의 예상된 시작 비용 $4500 단백질 생산의 µ g 당 0.021 달러 또는 $0.019 반응의 µ L 당 예상 비용 반응의 수천을 유지할 것입니다. 또한, 단백질 식 방법 거울 최적화 시 사전 믹스, 비용의 일부분에서의 상용 시스템에서 본 반응 설치의 용이성. 광범위 한 응용 프로그램에 대 한 CFPS 플랫폼의 유연한 특성을 활용 하 여 사용자 수 있도록, 다양 한 조정 하 고 사용할 수 있는 리소스와 단백질 식 결과 따라 최적화 될 수 있는 플랫폼의 측면 확인 했습니다. 원하는.
Introduction
셀 무료 단백질 합성 (CFPS) 등에 대 한 단백질 생산, 기능 유전체학, 대사 공학, 지난 50 년1,2내에서 새로운 기회의 번호를 잠금 해제는 기술로 떠오르고 있다. 표준 vivo에서 단백질 식 플랫폼에 비해, CFPS 세 가지 주요 이점을 제공 합니다: 1) 플랫폼의 셀 무료 자연 수는 잠재적으로 독성 또는 셀3,4 외국 것 단백질의 생산 ,,56; 2) 게놈 DNA의 비활성화 및 서식 파일의 gene(s)를 인코딩 하는 DNA의 소개의;는 protein(s)의 생산에 대 한 반응 내에서 조직의 에너지의 모든 채널 그리고 3) 플랫폼의 오픈 자연 수정 하 고 반응 조건 및 실시간7,8구성 모니터링 사용자 수 있습니다. 반응에 대 한이 직접 액세스 지원 확장 된 화학 및 산화 환 원 조건 새로운 단백질의 생산 및 대사 과정2,9, 의 튜닝에 대 한 생물 학적 시스템의 확대 10. 또한 수 있습니다 더 빠른 디자인-빌드-테스트11주기 위한 단일 포트 시스템에 CFPS 반응 활동 분석 실험을 결합 하 여 사용자를 직접. 작은 볼륨 방울 CFPS 반응을 수행 또는 종이 기반 장치에 더 높은 처리량 검색 노력을 지원 및 신속한 프로토 타입12,,1314,15 용량 ,16. 이러한 장점와 시스템의 플러그 앤 플레이 성격, CFPS 고유 어렵다 녹 표현 vivo에서단백질의 생산 등 생명 공학 응용 프로그램의 다양 한 활성화는17, 18,,1920, 질병21,,2223, 수요 biomanufacturing18,24 의 검출 25,,2627및 교육28,29, 유연성과 셀 무료 플랫폼의 유틸리티 모두 보기.
CFPS 시스템 모두 간결한과 진 핵 세포 라인에서 원유 lysates의 다양 한에서 생성할 수 있습니다. 다양 한 옵션 선택, 시스템에서 각각의 장점과 단점의 응용 프로그램에 따라 수 있습니다. CFPS 시스템 또한 준비 시간, 비용 및 생산성에 크게 다릅니다. 가장은 일반적으로 활용 하는 세포 추출 물 밀 세균, 토끼 reticulocyte, 곤충 세포, 및 후자 되 단백질30의 가장 높은 체적 수확량을 생산 하는 동안 날짜를 가장 비용 효율적인 대장균 세포에서 생산 됩니다. . 다른 CFPS 시스템 될 수 있지만 그들의 타고 난 포스트 번역 상 수정 기계에 대 한 유리한, 신흥 대장균을 사용 하 여 응용 프로그램-기반된 기계 site-specifically phosphorylated 생성 하 여 격차를 해소 수 있다 고 수요31,32,33,,3435당화 단백질.
CFPS 반응 중 일괄 처리, 연속 교환 셀 프리 (CECF) 또는 연속 흐름 셀 무료 (CFCF) 형식에서에서 실행할 수 있습니다. 일괄 처리 형식은 닫힌된 시스템 반응 수명이 감소 양의 반응 및 반응의 억제 부산물의 축적으로 인해 제한 됩니다. CECF 및 CFCF 메서드는 반응의 수명을 늘릴를 그로 인하여 증가 체적 단백질 수율 일괄 처리 반응에 비해. 이것은 새로운 반응 반응2과정을 통해 제공 하는 동안 반응 용기에서 제거할 단백질 합성의 부산물을 함으로써 수행 됩니다. CFCF, 경우 관심의 단백질은 또한 CECF, 반 투과성 막36,37의 구성 하는 반응 챔버에 관심이 남아의 단백질에에서 반응 약 실에서 제거할 수 있습니다. 이러한 메서드는 관심38,39,40,,4142, 표현 하기 어려운 단백질의 가난한 체적 수확량을 극복에서 특히 귀중 한 43. CECF 및 CFCF 접근을 구현 과제는 1) 동안 그들은 결과 바이오 기계 전사 및 번역에 대 한 책임을 더 효율적으로 사용, 그들은 특히 2 및 전반적인 비용을 증가 시 약의 대량 필요) 그들은 더 복잡 한 반응 설정 및 배치 형식44에 비해 특수 장비 필요 합니다. 새 사용자에 대 한 접근성을 보장 하기 위해 프로토콜 설명 여기 반응이 볼륨 증가 밀리 리터 규모 반응 볼륨에 대 한 특정 권장 사항을 15 µ L의 배치 형식에 초점.
여기에 소개 하는 방법을 구현 세포 성장, 준비, 추출 및 형식 반응 설치는 대장균에 대 한 일괄 (학부 학생) 등 기본적인 실험실 기술 전문가 들은 활성화-CFPS 시스템을 기반으로. 이 방법은 반응 키트 기반 설치의 용이성을 희생 하지 않고 상업적으로 사용 가능한 키트에 비해 비용 효과적입니다. 또한,이 방법을 통해 응용 프로그램에서에서 실험실 그리고 분야에서. CFPS를 구현 하도록 결정, CFPS 모든 경우에 우수한 되지 않을 수도 있습니다 새로운 사용자 시작 투자에 대 한 기존의 단백질 표정 시스템의 효능을 평가 철저 하 게 해야 합니다. 여기에 설명 된 CFPS 메서드는 사용자가 직접 구현 하는 다양 한 응용 프로그램, 기능 유전체학, 높은 처리량 테스트, vivo에서 식 필드에 대 세공 되지 않은 단백질의 생산을 포함 하 여를 사용합니다 바이오 센서 및 합성 생물학에 대 한 교육 키트를 포함 하 여 응용 프로그램. 대사 공학 같은 추가 응용 프로그램 단백질 식 조건, 질병 탐지, 및 확장을 사용 하 여 CECF 또는 CFCF 메서드가 여전히 가능 하지만 반응의 추가 수정에 대 한 CFPS 플랫폼 경험을 해야 튜닝 조건입니다. 우리의 방법을 상대적으로 신속 하 고 재현성 쥡니다, 최적화 된 예비 배합 물45를 사용 하는 간소화 된 CFPS 반응 설치 다음을 통해 세포 세포의 용 해 방법 풍부한 미디어 및 당황 하 게 플라스 크, 성장에 결합 되어 있습니다. 세포 성장 방법이이 분야 내에서 다소 표준화 되, 하는 동안 세포 세포의 용 해 방법 넓게 변화 한다. 쥡니다, 뿐만 아니라 일반적인 세포의 용 해 방법 포함 프랑스 언론 활용, 한 균질 화기, 비드 고물 차, 또는 lysozyme 다른 생 화 학적 및 물리적 중단 방법46,47,48, 49. 조 세포 추출 물의 약 2 mL 셀의 1 L 당 가져온 우리의 방법을 사용 하 여. 이 양의 세포 추출 물 4 백을 지원할 수 있습니다 15 µ L CFPS 반응, 각 생산 ~ 900 µ g/mL 템플릿 플라스 미드 pJL1-sfGFP에서 기자 sfGFP 단백질의. 이 방법은 비용 $0.021 / µ g의 sfGFP 제작 ($.019 / µ L의 반응), 노동 및 장비 (보충 그림 1)의 비용을 제외 하 고. 처음부터 시작,이 방법은 한 사람에 의해 4 일에 구현할 수 있다 고 CFPS 반응 시간 (그림 1) 내에서 완료 될 수 있다 반복 합니다. 또한, 프로토콜 사용자의 요구에 맞게 시 약 준비의 큰 배치에 대 한 볼륨 확장할 수 있습니다. 중요 한 것은, 여기에 제시 된 프로토콜만 기본적인 실험실 기술을 필요로 실험실 훈련 비-학부 학생 등 전문가 의해 구현할 수 있습니다. 아래에서 설명 하는 절차와 함께 비디오 특별히 개발 되었습니다 광범위 한 사용에 대 한 대장균 CFPS 플랫폼의 접근성을 개선 하기 위해.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. 미디어 준비 및 세포 성장
-
주 1
- 글리세롤에서 행진 대장균 BL21*(DE3) 셀 파운드 한 천 격판덮개에 재고 및 적어도 18 h 37 ° c.에 품 어
- 121 ° c.에 30 분 동안 액체 주기 파운드 미디어와 고압 솔루션 50 mL를 준비 실내 온도에 상점.
-
주 2
- 추가 정보에 설명 된 대로 2 x YTP 미디어와 0.4 M D-포도 당 솔루션의 250 mL 750 mL를 준비 합니다.
- 2 x YTP 미디어를 압력가 2.5 L 당황 하 고 플라스 크와 D-포도 당 솔루션 압력가 500 mL 유리병에 붓으십시오. 고압 액체는 121 ° c.에 30 분에 두 솔루션
- 세포 성장 접종 시 성장 속도 최대화 하기 위해 다음 날에 수행 하는 경우 모두 살 균 솔루션 37 ° C에 저장 됩니다 확인 하십시오. 접종까지 솔루션을 결합 하지 마십시오.
참고: 솔루션 2 x YTP 미디어는 높게 오염 하는 경향이 있지만, 필요한 경우 1-2 d 4 ° C에 저장할 수 있습니다. - 오염을 피하기 위하여 소독된 루프 및 무 균 기법을 사용 하 여 BL21(DE3)의 단일 식민지와 파운드 미디어의 50 mL를 접종 하 여 BL21(DE3)의 숙박 문화를 시작 합니다.
- BL21*(DE3) LB 문화 50 mL 37 ° C에 250 rpm 인큐베이터를 흔들어 놓고 하룻밤 15-18 h에 대 한 성장.
- 준비 하 고 3, 4, 등 일에 필요한 모든 재료를 소독: 두 1 L 원심 분리기 병, 4 x 찬 50 mL 원뿔 튜브 (무게 및 3에 대 한 기록 중), 그리고 많은 1.5 mL microfuge 관.
-
3 일
- 떨고 인큐베이터에서 파운드에 BL21*(DE3)의 50 mL 숙박 문화를 제거 하 고 측정 하는 1시 10분을 사용 하 여 분 광 광도 계에 OD600 파운드 미디어와 함께 희석. 계산 시작 OD600 0.1에 대 한 미디어의 1 패를 추가 하는 데 필요한 하룻밤 문화의 볼륨 (예를 들어 1:10 희석은 0.4, 읽기의 세600 undiluted OD600 의 25 mL를 접종 하는 경우 = 4.0 하룻밤 문화 1 L 2로 x YTP G)입니다.
- LB 문화 50 mL와 함께 37 ° C 배양 기에서 따뜻하게 2 x YTP 미디어와 D-포도 당 솔루션을 제거 합니다. 무 균 기술을 사용 하 여 신중 하 게 붓고 D-포도 당 솔루션 (당황 하 고 플라스 크의 측면을 피하고) 2 x YTP 미디어.
참고: D-포도 당의 추가 완료 2의 1 L에 대 한 제조 법 x YTPG. - 유지 하 고 메 마른 기술, 접종 0.1 세600에서 1 L 문화를 시작 적절 한 양의 50 mL 문화 2 x YTPG 솔루션의 1 리터. 즉시 200 rpm에서 인큐베이터를 떨고 37 ° C에 접종된 1 L 문화 장소.
- 받아 성장의 첫 번째 시간 후 읽기 첫 번째 OD600 (지연 위상 전형적인 1 시간 소요). 문화를 희석 하지 마십시오. 측정 세600 매 20-30 분 약 세600 0.6에 도달할 때까지 계속 합니다.
- OD600 에 도달 되 면 = 0.6, 1 M IPTG의 1 mL을 추가 (최종 농도 1 L 문화에서 = 1 m m) 2 x YTPG 문화.
참고: 이상적인 유도 OD600 은 0.6; 그러나, 0.6-0.8의 범위는 허용. IPTG에 의해 유도 생 생산의 T7 RNA 중 합 효소 (T7RNAP)입니다. - 유도 후 측정 OD600 매 20-30 분 약 3.0에 도달할 때까지.
참고:이 시간 동안에 4 ° C 원심 분리기를 식혀. 추가 정보에 설명으로 차가운 S30 버퍼를 준비 합니다. S30 버퍼는 사전에 준비 하 고, DTT 사용의 날까지 추가 되지 않습니다 확인 합니다. - OD600 3.0 (그림 2A)에 도달 하면, 문화는 얼음 물을 욕조에 차가운 1 L 원심 분리기 병에 붓으십시오. 원심 분리기에 균형으로 사용 될 동등한 무게의 물 가득 1 L 원심 분리기 병을 준비 합니다.
참고: 흡 광도 값 악기 악기에서 다. 동안 BL21(DE3)의 수확의 OD600 민감한 변수, 사용자 평가 및 문제 해결 측정으로이 변수를 최적화 하는 것이 좋습니다. 큰 광 작은 cuvette 기반 광에 비해 상대적으로 낮은 OD600 읽기 발생할 수 있습니다. - 1 L 병 5000 x g와 작은 셀에 10 ° C에서 10 분 원심
- 천천히 상쾌한 및 기관의 생물 학적 폐기물 절차에 따라 처분 하 거 라. 얼음에 펠 릿을 놓습니다.
- 메 마른 주걱을 사용 하 여, 원심 분리기 병에서 셀 펠 릿을 긁 고 찬 50 mL 원뿔 튜브에 그것을 전송.
- 차가운 S30 버퍼의 30 mL 원뿔 튜브에 추가 하 고 완전히 없는 청크와 resuspended 때까지 얼음에 짧은 파열 (20-30 s)와 나머지 기간 (1 분) vortexing에 의해 셀 펠 릿 resuspend.
- 일단 펠 릿 resuspended 완전히 x g와 10 ° C (-4 ° C에 냉각) 5000에서 밸런스와 10 분 동안 원심 분리기로 다른 50 mL 원뿔 튜브 물으로 사용 합니다.
참고: 셀을 수확 때 필요한 3 세척의 1세인트 완료 했습니다. - 상쾌한 및 기관의 생물 학적 폐기물 절차에 따라 처분 하 거 라. S30 버퍼 춥고 5000 x g와 10 ° C (-4 ° C에 냉각) 10 분 동안 원심 분리기의 20-25 mL와 펠 릿 resuspend
참고: 3 세척의 2차 완료 했습니다. - 다시, 상쾌한 및 기관의 생물 학적 폐기물 절차에 따라 처분 밖으로 붓는 다. S30 버퍼 및 resuspend는 펠 릿을 다시 소용돌이의 정확 하 게 30 mL를 추가 합니다.
- 사용 3 미리 무게, 차가운 50 mL 원뿔 튜브와 혈 청 학적인 피 펫 필러 살 균 피 펫, aliquot와 resuspended 펠 릿/s 30 버퍼 혼합물의 10 mL 3 원뿔 관의 각각에.
참고: 3 관으로 셀 분할는 필요 하지 않습니다, 하지만이 단계 작은 세포 펠 릿에 결과 (~ 1 g) 나중 단계에서 증가 편의 위해. - X g와 10 ° C (-4 ° C에 냉각) 5000에서 10 분에 대 한 필요에 따라 적절 한 균형을 사용 하 여 모든 튜브 원심
참고:이 최종 세척 단계를 완료합니다. - 상쾌한 및 기관의 생물 학적 폐기물 절차에 따라 처분 하 거 라. 신중 하 게 원뿔 튜브 및 깨끗 한 조직; 캡의 내부를 닦아 하 여 초과 S30 버퍼를 제거 펠 릿을 만지지 마십시오.
- 분석 균형에 튜브를 reweigh 하 고 각 관에 마지막 펠 릿 체중을 기록 합니다.
참고: 프로토콜 수 수 일시 중지이 시점에서. 액체 질소에서 냉동 고 최대 1 년까지 추출 준비를 위한 필요-80 ° C에서 저장 된 펠 릿 플래시 수 있습니다.
2. 원유 셀 추출 준비-주 4
- 추출 물 준비를 위해 계속 셀 차가운 얼음에 각 단계. 펠 릿의 세포 질량의 1 g 당 찬 S30 버퍼의 1 mL를 추가 합니다. 그 dithiothreitol (DTT) 2 m m의 최종 농도에 s 30 버퍼에 보완 되어 있는지 확인 합니다.
참고:이 기간 동안 4 ° C에 microcentrifuge를 식혀. - 얼음까지 완벽 하 게 resuspended에 짧은 파열 (20-30 s)와 나머지 기간 (1 분) vortexing에 의해 셀 펠 릿을 resuspend. 물의 resuspension 어려운 경우 서 리 없애다에 30 분 동안 얼음에 펠 릿을 둡니다.
- Resuspended 셀의 1.4 mL 1.5 mL microfuge 관으로 전송 합니다.
- Resuspended 셀의 1.4 mL 비 커에 얼음 물 목욕으로 포함 한 1.5 mL 튜브를 배치 합니다. 45 sonicate 59 다음에 s 3 총 주기, 진폭에 대 한에서 s 50%로 설정. 닫고 오프 기간 동안 섞어 부드럽게 튜브를 반전 합니다. 총, 포함 하는 resuspended 세포 (그림 3A & 3B)의 1.4 mL 각 1.5 mL microfuge 관에 800-900 J의 에너지를 제공 합니다.
참고:이 단계는 sonicator 종류에 민감한 모델 사용 하 고 장비는이 절차에 나열 된 것 보다 다른 경우 최적화 되어야 합니다. 두 개의 상호 보완적인 접근을 추출이이 단계 동안 준비의 금액을 사용할 수 있습니다: 1) 여러 1.5 mL microfuge 관 병렬로 sonicated 수 / 2) 더 큰 볼륨 원뿔 튜브에서 sonicated 수 있습니다 (최대 셀 튜브 당 물의 resuspension의 15 mL) 전달 하는 에너지의 양을 확장으로 설명한 29,45. - 즉시 완료 된 후 쥡니다, lysate의 1.4 mL에 1 M (보충 추가 2 mM DTT) DTT의 4.5 µ L을 추가 하 고 섞어 여러 번 반전. 얼음에 튜브를 놓습니다. 원심 분리를 진행 하기 전에 resuspended 세포의 어떤 추가 튜브에 대 한 2.4 및 2.5 단계를 반복 합니다.
- 18000 x g와 10 분 (그림 3C) 4 ° C에서 Microcentrifuge 샘플.
- 새로운 1.5 mL microfuge 관으로는 상쾌한 플라스틱. 펠 릿;를 방해 하지 마십시오 수익률을 극대화 하는 노력에 펠 릿을 방해 보다 순도 유지 하기 위해 일부 상쾌한을 남겨두고 하는 것이 바람직합니다.
- (이것은 결선 반응) 인큐베이터의 떨고 플랫폼에 튜브를 녹화 하 여 상쾌한 250 rpm 및 60 분 동안 37 ° C에 이전 단계에서 품 어.
- 10000 x g와 10 분 동안 4 ° C에서 Microcentrifuge 샘플.
- 펠 릿을 방해 하지 않고는 상쾌한을 제거 하 고 새 튜브를 전송. 스토리지에 대 한 추출의 많은 100 µ L aliquots를 만듭니다.
참고: 프로토콜 일시 중지할 수 있습니다 여기에, 그리고 액체 질소에서 냉동 고 CFPS 반응에 필요한 때까지 1 년까지-80 ° C에 저장 된 추출 플래시 수 있습니다. 생산성 (그림 4)를 추출할 손해 없이 적어도 5 freeze-thaw 주기를 겪고 수 있습니다.
3. 셀 무료 단백질 합성 배치 형식 반응
- Thaw 솔루션 A와 B, DNA 템플릿, BL21(DE3) 추출 (냉동) 하는 경우, T7RNAP, 고 분자 학년 물 약 수.
참고: CFPS 반응 템플릿은 추가 정보에서 찾을 수 있습니다. 솔루션 A와 B 요리법 보충 정보 에 제공 되며 CFPS 위한 PANOx SP 기반으로 에너지 시스템을 지원 하기 위해 수많은 시 약에 대 한 특정 농도에 해당. 각 시 약 및 CFPS 지원할 수 있는 이러한 시 약 농도에서 허용 가능한 변이의 역할 결정된50되었습니다. T7RNAP 정화 프로토콜 추가 정보51에서 찾을 수 있습니다. 추가 T7RNAP 체적 수율을 높일 수 있습니다 하지만 T7RNAP 동안 세포 성장을 유도 하는 경우 필요 하지 않습니다. 플라스 미드 DNA 템플렛 (pJL1-sfGFP) 두 세척 키트, 후 처리 DNA 정리 PCR 정화 키트 (그림 2B)를 사용 하 여 뒤에 워시 버퍼를 사용 하 여 maxiprep 키트를 사용 하 여 준비 될 수 있다. 선형 DNA 템플릿 CFPS 반응에도 사용할 수 있습니다. - 필요한 양의 microfuge 관 CFPS 반응에 필요한 레이블을 지정 합니다.
참고: 반응 다양 한 선박 크기에서 수행할 수 있습니다 하지만 작은 용기 체적 단백질 수율 (그림 2C)을 줄일 수 있습니다. 같은 크기 용기에 반응 확장 기능 감소 하는 산소 교환 볼륨 비 표면적의 감소 때문으로 체적 수익률 또한 줄일 수 있습니다. 100 μ 이상 반응이 볼륨 증가, 플랫 바닥 잘 플레이트 31,,3752를 사용 하는 것이 좋습니다. - 추가 솔루션의 2.2 µ L, 2.1 µ L 솔루션 B, BL21*(DE3)의 5 µ L의 추출, T7RNAP의 0.24 μ g (16 μ g/mL 최종 농도) 0.24 DNA 템플렛 (16 ng/mL 최종 농도) 및 15 µ L를 최종 볼륨을가지고 물 ng.
참고: A와 B 자주 소용돌이 솔루션 반응 설치 부품의 침전을 방지 하 고 각 반응 하는 동안 받습니다 각 솔루션의 동종 약 수 있습니다. 혼합 튜브 대신 반전, vortexing 추출 하지 마십시오. - 모든 시 약 반응에 추가 되었습니다, 최종 반응 혼합물을 1.5 mL microfuge 관의 맨 아래에 단일 15 µ L 비드 결합 하면서 위아래로 pipetting 또는 부드럽게 vortexing 각 튜브를 혼합.
- 37 ° C 배양 기에 각 반응 없이 하룻밤 4 h 또는 30 ° C를 흔들어 놓습니다.
참고: 성공적인 반응은 수 수 질적 평가 시각 CFPS 반응 혼합물 (그림 3D) 이내 sfGFP 제품의 녹색 색상에 따라. 관심사의 단백질의 표정 또한 SDS 페이지 (보충 그림 2)에 의해 확인할 수 있습니다.
4. 기자 단백질, [sfGFP]의 정량화
- 잘 정량화 (일반적으로 반응 관 당 3 중에서 수행 됨)에 필요한 각 0.05 m HEPES, pH 8, 48 µ L를 로드 합니다.
- 인큐베이터에서 반응을 제거 합니다. 위쪽 및 아래쪽 각 반응 혼합 플라스틱 다음 0.05 M HEPES, pH 8의 48 µ L에 반응의 2 µ L를 전송 합니다. 피펫으로 위쪽 및 아래쪽에 다시를 섞어 잘.
- 일단 모든 반응 로드 혼합, 96 잘 접시는 fluorometer로 놓고 sfGFP 끝점 형광을 측정 합니다.
참고: sfGFP 형광 정량화에 대 한 여기 및 방출 파장은 485 nm와 510 nm, 각각. - 이전에 생성 된 표준 곡선을 사용 하 여 얻은 형광 신호에서 [sfGFP]를 확인 합니다.
참고: 추가 정보 (보충 그림 3)에 형광 강렬 대 sfGFP 농도의 표준 곡선을 생성 하기 위한 지침 제공 됩니다. 사용자는 악기 감도 다를 수 있습니다 이후 그들의 악기에 대 한 표준 곡선을 설정 해야 합니다.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
우리 쥡니다 기반 대장균 추출 제시 준비 프로토콜을 매일 절차 분석을 보여주는 그림 1 4 일 기간 동안 완료 될 수 있다. 가 단성 다양 한 일시 중지 포인트, 매일에 완료할 수 있는 단계에 있다 하지만 우리이 워크플로 실행에 가장 효과적인 것으로 나타났습니다. 또한, 셀 펠 릿 (단계 1.3.18)와 완벽 하 게 준비 된 추출 (단계 2.10)는 안정-80 ° C에서 1 년 이상 사용자 나중 시간17에 사용 하기 위해 저장 하려면 각의 더 큰 주식을 만들 수 있도록. 뿐만 아니라 추출 안정 오래 기간 동안 이지만 추출 또한 생산성 (그림 4)의 뜻깊은 손실 없이 적어도 5 동결 해 동 주기를 받을 수 있습니다. 이 추출의 큰 aliquots 냉장고 저장 공간이 한정 된 경우 여러 용도로 저장 될 수 있습니다. 그러나, 가능 하면 추출의 여러 개의 작은 aliquots를 (~ 100 μ)이 좋습니다.
모든 새로운 추출 준비, 사용자 추출의 해당 일괄 처리에 대 한 마그네슘의 최적의 금액을 결정 하기 위하여 마그네슘 적정을 수행 하는 것이 좋습니다. 사용자가 Bradford 분석 실험에 의해 셀 추출의 총 단백질 농도에 일괄 배치 변화를 계량 수 있습니다. 우리가 일반적으로 30-50 mg/mL의 총 단백질 농도 참조 높은 수행 추출 물, 고이 범위 내에서 총 단백질 농도 세포 추출 성능 사이 분명 한 상관 관계가 있다. 따라서,는 사용자 조정 마그네슘 농도 따라 각 추출 일괄 처리에 대 한 단백질 및 핵 산 기능 극대화 되도록 하는 것이 좋습니다. 마그네슘 수준 적절 한 DNA 복제, 전사 및 번역에 대 한 중요 하지만 과도 한 수준은 이러한 프로세스53에 해로운 수 있습니다. 이 종속성을 설명 하기 위하여 우리는 마그네슘의 공동 적정 수행 하 고 볼륨 추출 생산 반응 (그림 5)을 유지 하면서 필요한 금액을 최소화 하는 최적의 조합 결정을 추출. 이 실험에서 sfGFP의 1000 μ g/mL를 받으려면 30 mg/mL의 총 단백질 함량과 추출의 추출 및 10 m m Mg2 + 5 µ L를 사용 하는 것이 좋습니다.
CFPS 우리의 경험 또한 높은 성능의 CFPS 시스템 통합, 시스템의 전반적인 생산성에 손해 없이 변화 될 수 있는 프로토콜 및 다른 단계를 결정 하 게 있었습니다. 특히, 셀 수확의 마지막 세600 크게 영향을 주지 않습니다 CFPS 반응의 최종 출력 그리고 셀 feasibly을 어디에서 든 지 2.7-4.0 세600으로 수확 하실 수 있습니다. 이 셀 당 리보솜 농도 최고 하 고 변환 기계 장치는 가장 급속 한 성장을 지원 하기 위해 적극적인 성장의 초기 지 수 단계를 나타냅니다. 이 관찰은 사용자가 그들의 자신의 절차를 최적화 하는 유연성을 수 있습니다. 완전 한 (그림 2A)은 수확 하는 시간으로는 세600 가까이 3.3 셀을 캡처하기 위해 약 3.0 OD600 에서 수확 하는 것이 좋습니다. CFPS 수율에 영향을 미칠 할 변수는 템플릿 DNA, 반응 선박 크기, 품질과 상대적 양의 셀 추출 물과 마그네슘 이온 반응에 포함 됩니다. 우리는 DNA 품질 주목할 만한 배치-배치 변화를 발견 했다. 이 문제를 해결 하려면 사용자가 midi 또는 맥시 준비 뒤 DNA 정화 열에는 maxiprep에 사용 되는 추가적인 DNA 정리 단계 또는 추가 DNA 정리 키트를 사용 하 여 후 정화를 통해 DNA를 정화 하는 것이 좋습니다. CFPS 반응에 대 한 DNA 품질 재현성 및 결과 (그림 2B) 더 강력한 단백질 생산에서 향상 됩니다. 또한 반응 배는 동일한 반응 설정 다양 한 선박 볼륨에서의 단백질 생산 최대 40%까지 다를 수 있습니다의 체적 수율에 미치는 영향. 큰 혈관에서 관찰 하는 체적 수율에서 향상은 반응 혼합물, 더 나은 산소 교환 (그림 2C), 허용의 증가 노출 영역 및 다른 사람 더 실행 하 여 체적 수확량 증가 이론 되었습니다. CFPS 반응은 큰 평면 바닥 접시는 100 µ L17,,3137,52이상 반응에 대 한 것이 좋습니다.

그림 1 : 문화 성장, 세포 추출, 설치 및 CFPS 반응의 정량화의 생산 일정. 사용자는이 4 일간 워크플로 통해 그들의 연구 응용 프로그램의 CFPS 플랫폼을 구현할 수 있습니다. 시 약 준비 기본 시간과이 실험의 첫 번째 라운드에 대 한 비용 투자를 대표 하 고 시 약 주식은 설립 후 실질적으로 감소. 또한, 셀 펠 릿 및 준비 세포 추출 물 저장할 수 있습니다에 대 한 빠른 결과 대 한 다양 한 단계에서 타임 라인을 시작 하는 사용자를 허용-80 ° C에서 1 년 이상. 사용자 수 있습니다 또한이 워크플로의 타임 라인을 수정 하기 위해 다양 한 단계에서 일시 중지 합니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭 하십시오.

그림 2 : CFPS 및 체적 반응 수율에 미치는 영향에 대 한 수정 조건. A. 수확 하는 다양 한 세600 수치에서 BL21(DE3) 셀 기반 생산성 비교를 추출 합니다. 3.3 대상 단백질의 적어도 1000 µ g/mL를 생산 하는 OD600 에서 수확이 플롯을 바탕으로, 좋습니다. 반응 15 μ 규모 1.5 mL microfuge 관에서 수행 했다. B. 두 개의 DNA maxiprep 세척 프로토콜 후 정화 DNA-정리 없이의 비교. pJL1-sfGFP 플라스 미드 PCR 정화 키트 후 정화 정리에 의해 선행 하는 하나 또는 두 개의 세척으로는 maxiprep를 받았다. 를 달성 하기 위해 ~ 900 µ g/mL 단백질 식의 maxiprep 세척 수와 관계 없이 사후 정화 DNA 정리를 수행 하는 것이 좋습니다. 반응 15 μ 규모 1.5 mL microfuge 관에서 수행 했다. C. 15 µ L CFPS 반응 2 mL에서 0.6 mL microfuge 관에 이르기까지 다양 한 선박에서 수행 합니다. "Neg" DNA 템플릿이 없는 반응에 추가 되었습니다 부정적인 컨트롤을 나타냅니다. 모든 오차 막대는 각각의 3 중에서 계량 된 각 조건에 대 한 3 개의 독립적인 반응의 1 표준 편차를 나타냅니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭 하십시오.

그림 3 : 키 절차 설정 및 생산성을 만들기 위한 결과 추출 합니다. A. 열 쥡니다 동안 생성 되는 동안 샘플의 냉각 되도록 쥡니다 얼음 물 목욕의 적절 한 설치. B. 1.5 mL microfuge 관 resuspended 셀 펠 릿 전 (왼쪽) 및 게시물 (오른쪽) 쥡니다. 그 결과 lysate resuspended 셀 펠 릿에 비해 어두운 색조를 표시 되어야 합니다. C. 상쾌한 세포 lysate 18000 x g 원심 분리 후의 펠 릿의 적절 한 분리. D. CFPS 반응 4 h 후 37 ° c.에 외피의 오른쪽 (성공적인 반응)에 1.5 mL microfuge 관 ~ 900 μ g/mL에서 sfGFP 기자 단백질의 보이는 형광을 보여준다. 왼쪽 서식 파일 DNA를 부족 하 고 실패 한 반응 시뮬레이션에 부정적인 제어 튜브 아무 형광으로 명확한 솔루션을 표시 합니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭 하십시오.

그림 4 : 단백질 식 이상 5 freeze-thaw 주기 CFPS 변경 추출. 같은 성장에서 준비 된 추출 수술 5 액체 질소를 통해 해 동 주기를 얼음에 녹고에 의해 뒤 냉동 플래시 동결. SfGFP를 표현 하기 위한 추출 생산성에 중요 한 변경 5 freeze-thaw 주기 동안 보였다. 반응 15 μ 규모 1.5 mL microfuge 관에서 수행 했다. "Neg" DNA 템플릿이 없는 반응에 추가 되었습니다 부정적인 컨트롤을 나타냅니다. 모든 오차 막대는 각각의 3 중에서 계량 된 각 조건에 대 한 3 개의 독립적인 반응의 1 표준 편차를 나타냅니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭 하십시오.

그림 5 : CFPS 변화 반응에 대 한 [Mg2+] [sfGFP] 대 볼륨을 추출 하 고. [Mg2 +] 8 m m에서 2mm 단위로 14 mM에 배열 했다 고 압축 볼륨 1 µ L 단위로 7 µ L 3 µ L에서 배열 했다. 색상 코드 나타냅니다 [sfGFP] 낮은 (보라색) (레드)에서 생산. 높은 단백질 생산을 유지 하면서 시 효율을 극대화 하기 위해 추출 및 10 밀리미터 밀리 그램 5 µ L를 사용 하 여 것이 좋습니다2+ ~ 30 mg/mL의 총 단백질 함량을 Bradford 분석 실험에 의해 결정 되는 추출 물에 대 한. 원래 포인트 컨투어 플롯을 생성 하는 각각의 3 중에서 측정 했다 각 조건에 대 한 3 개의 독립적인 반응의 끝점 형광을 기반으로 했다. 반응 15 μ 규모 1.5 mL microfuge 관에서 수행 했다. 이 그림의 더 큰 버전을 보려면 여기를 클릭 하십시오.
보충 그림 1: 6 셀 무료 단백질 합성 플랫폼에서 생산 하는 단백질의 그램 당 및 반응의 microliter 당 비용. 우리의 플랫폼 생산성 변화 및 가격 5 다른 셀 무료 단백질 합성 키트/플랫폼을 비교 합니다. 우리의 기반 쥡니다 CFPS 플랫폼 보다 대부분 상업 키트 키트의 용이성을 제공 하는 나머지 비용 다른 학술 CFPS 플랫폼을 비교 하면서 반응 설정에 대 한 단백질의 두 $/ µ g에 $/ µ L 반응의 더 비용 효율적입니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭 하십시오.
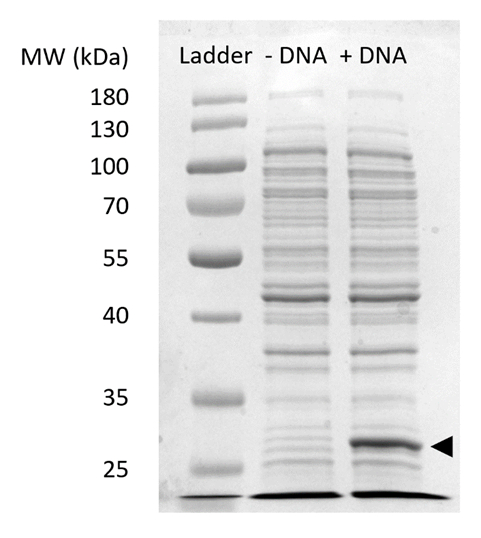
보충 그림 2: CFPS에 sfGFP 식의 SDS 페이지. 셀-무료와 단백질 합성 반응 (+ DNA) 없이 (-DNA) sfGFP에 대 한 DNA 템플렛 27 kDa (검은색 화살표)에서 관찰 하는 sfGFP의 식을 보여 주기 위해 SDSPAGE 젤 12%에서 실행 했다. 전통적인 SDS 페이지 기술은 사용 했다. 각 샘플 셀에 총 단백질의 Bradford 분석 결과 부 량에 따라 총 단백질의 포함된 18 µ g 추출 하는 젤에 로드. 우리는 추정 형광 강도 측정 및 우리의 표준 곡선을 바탕으로는 "+ DNA" sfGFP의 0.42 µ g를 포함 하는 차선. 이러한 샘플을 받으려면, CFPS 반응 15 µ L 규모 1.5 mL microfuge 관 체적 수익률 그림 3C와 생산에서 실행 되었습니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭 하십시오.
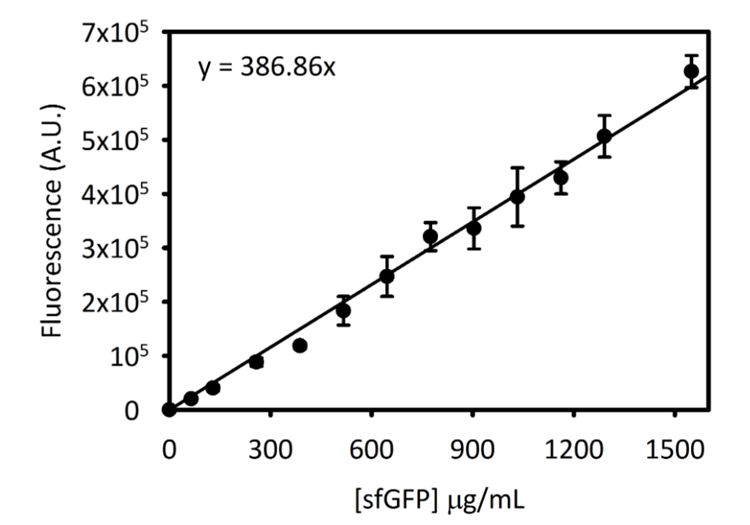
보충 그림 3: Cytation 5 sfGFP에 대 한 표준 곡선. 이 곡선 위에서 설명한 방법을 사용 하 여 결정 했다. 이 원고는 변환 된 끝점 형광 신호에서 [sfGFP] µ g/ml이 표준 곡선을 사용 하 여 수집 된 모든 데이터. 이 그림의 더 큰 버전을 보려면 여기를 클릭 하십시오.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
셀 무료 단백질 합성 응용 프로그램 biomanufacturing에서 생화학 시스템의 신속한 프로토 타입에 이르기까지 다양 한 강력 하 고 사용 기술로 떠오르고 있다. 애플 리 케이 션의 광범위 한 모니터링 하 고 조작 하 고, 실시간으로 세포질 기계 장치를 보강 하는 능력에 의해 지원 됩니다. 이 플랫폼 기술의 확대 영향에도 불구 하 고 광범위 한 적응 남아 있다 메서드의 구현에서 기술적인 뉘앙스로 인해 느린. 이 노력을 통해 우리는 새로운 실험실에이 기술을 설정에 대 한 단순 하 고 명확을 제공을 목표로. 이 끝으로, 대장균에 대 한 우리의 프로토콜-기반된 셀 무료 단백질 합성 플랫폼 실험실 훈련 비-전문가, 학부생 (그림 1) 등으로 4 일의 시작 시간에서 달성 될 수 있다. 또한, 시 약 및 추출의 재고 생산, 일단 후속 CFPS 배치 반응 설정, 인 큐베이 팅, 고 그냥 5 h에 계량. 단일, 1 L 세포 성장 될 수 있습니다 4 백 15 µ L CFPS 반응에 대 한 충분 한 추출 동안 단일 일괄 처리 준비는 다른 셀 무료 시 약의 반응의 수천을 지원할 수 있습니다. 더 큰 주식은 필요 시 약 준비를도 확장할 수 있습니다. CFPS 반응의 병렬에서 조건의 다양 한 테스트를 위해 96 잘 접시 또는 PCR 튜브를 사용 하 여 높은 처리 방식으로 설치 될 수 있습니다. 체적 수익률 그림 2C에서 보듯이 작은 혈관을 사용 하는 경우를 줄일 것 이다. CFPS 반응 또한 확장할 수 있습니다 microliters에서 총 반응 볼륨의 밀리 리터에 단일 조건에 대해 총 단백질 수확량을 증가 하기 위하여. 볼륨을 확장 하는 때 주요 고려 사항은 체적 반응 수율 감소 반응의 표면 영역 볼륨 비율 감소로37,52이다. 스케일 업 단백질 식의 비슷한 체적 수익률을 유지 하면서, 위해 사용자가 해야 수많은 반응 혈관으로 반응 볼륨을 분할 및/또는 선박 크기를 증가. 반응 비늘 15 µ L-볼륨, 100 µ L에서에서 배열 동시에 수많은 15 µ L 반응을 사용 하는 것이 좋습니다. 볼륨에서 100 µ L를 초과 하는 반응에 대 한 평면 바닥 24-잘 접시를 권장 하 고 12-잘 접시 600 µ L를 초과 하는 반응 볼륨에 대 한 추천 된다. 반응 볼륨 및 혈관의 같은 쌍 스케일 업17,31,,3752시 체적 반응 수율에 일관성을 제공합니다. 이러한 볼륨을 넘어 확장 하는 것은 동시에 접시의 여러 우물을 이용 하 여 수행할 수 있습니다. 이 형식을 사용 하 여, 반응 10 mL 전체 볼륨을 확장할 수 있습니다. 반응 볼륨 반응 선박 조합 최적화 반응의 생산성을 희생 하지 않고 biomanufacturing의 응용 프로그램을 지원할 수 있습니다.
이 프로토콜을 수행할 때 추출 저조한 연관 지표로 서 체적 반응 수율에 영향을 주는 몇 가지 주요 고려 사항 있다. 적절 한 세포를 확인 하 고 기능 녹음/번역 기계의 변성을 방지, 세포 동안 생산 하는 열을 완화 하기 위해 중요 하다. 빠르게 쥡니다 (그림 3A) 동안 열을 방산 하 쥡니다 동안 셀 물의 resuspension 얼음 물 욕조에 담가. 효과적인 세포 세포의 용 해의 지표로 사전 sonicated 샘플 (그림 3B)에 비해 세포 lysate의 어두운 외관의 출현 이다. 사용자 유연성을 sonicator 및 그림 3A 에 표시 된 프로브입니다 적응할 수 있는 볼륨 범위에 resuspended 세포의 15 mL를 100 µ L에서. 이 위해 사용자는 셀의 원하는 볼륨의 세포에 전달 하는 줄의 수를 조정할 수 있습니다. 또한, 추출의 더 큰 볼륨 두 가지 보완적인 방법을 통해 준비 될 수 있다. 사용자 수, 동시에 여러 개의 튜브를 sonicate 및/또는 앞에서 설명한29,45볼륨 비례와 에너지의 양을 확장 셀 물의 resuspension의 더 큰 볼륨을 sonicate. 추출 품질을 나타내는 또 다른 단계는 원심 분리 단계 다음 세포 세포의 용 해. 게시물 세포 세포의 용 해, 것이 좋습니다 제공 하는 상쾌한 (전사/번역 기계, 조각난된 genomic DNA 이상 서식 파일 전송/번역 기능) 사이 명확한 부분을 18000 x g에서 원심 분리와 펠 릿 ( 침전 된 단백질 및 세포 막과 같은 세포 구성 요소를 바라지 않는) (그림 3C). 우리는 원심 분리를 발견 했다 18000 x g 분리, 12000 x g 등 낮은 속도에서 회전에 비해 향상 된 재현성 결과 향상 시킵니다. 편의 위해, 테이블 상단 냉장된 원심 분리기, 12000 x g의 최소 달성 가능한를 사용 하는 것이 좋습니다. 이 단계는 또한 일반적으로 적절 한 장비는 사용할 수54,55,56,,5758, 하는 경우 30000 x g에서 수행 59 , 60. 추출 성능 영향을 받지 않습니다이 단계에서 원심 분리 속도 적절 한 분리를 달성 이다. 원하는 상쾌한을 제거할 때이 오염 추출의 생산성을 감소 시킬 것 이다 이후 상쾌한 펠 릿 사이 경계에 존재 하는 어떤 흐린 물질 최상 이다. 순도 상쾌한의 목표로 생산성 추출 결과 하 고 새로운 사용자에 대 한 얻은 추출의 감소 수량 가치가 있다.
그것은 우리가 제시 하는 방법을 재현할 수는 최소한의 전문성을 갖춘 과학자 들에 의해 실행 될 수 있습니다 하는 동안 일괄 처리에 배치 될 수 있는 메모와 반응에 반응 해야 합니다. 이것은 lysate 후 쥡니다61의 proteomic 구성 편차를 표시 될 수 있습니다. 우리가 관찰 일괄 배치 변화 일반적으로 T7RNAP와 보완 및 마그네슘 농도의 최적화에 점감 된다. T7RNAP의 추가 지 원하는 최적의 단백질 표정, CFPS 반응 중 일반적 이며 우리 T7RNAP의 두 소스를 데 그-생 식 BL21*(DE3) 및 16 µ g/mL의 최종 농도에 추가 T7RNAP-향상 찾을 외 인 새로운 사용자45,46에 대 한 일괄 배치 재현성. 경험으로, 사용자가 원하는 경우 T7RNAP의 단일 소스만을 활용 하는 그들의 실험을 수정할 수 있습니다. 추출의 새로운 배치와 Mg2 + 농도의 적절 한 조정의 총 단백질 함량의 부 량 또한 체적 단백질 식 수익률에 일괄 배치 변화를 줄이기 위해 도움이 됩니다. 단백질 표정에 변화 관심사의 단백질, 유전자의 코 돈 사용 및 해당 리보솜 바인딩 사이트 관심, 표정 벡터 사용62의 종류의 유전자의 구조와 크기에 차이 수도 있습니다. ,63. 이러한 이유로 일부 단백질은 모델 단백질 sfGFP, CFPS 반응에서 감소 체적 수율 결과 뿐만 아니라 표현 하지 않을 수 있습니다.
제시 CFPS 기술의 한계는 직접 셀-무료, 대사 공학 및 프로토콜에 대 한 추가 수정 없이 식 조건의 튜닝 등의 모든 응용 프로그램에 적합 하지 않을 수 있습니다를 포함 합니다. 그러나, 우리는이 프로토콜 새로운 실험실에 CFPS 플랫폼 구축을 위한 기초를 제공 하 고 비 전문가 소개 셀 무료 반응을 그들의 실험실에서 구현 하는 기능 제공 믿습니다. 초기 구현 후 연구 분야에서 다른 문학에 따라 특정 응용 프로그램에 대 한 그들의 자신의 수정 플랫폼 실험 수 있습니다.
CFPS 플랫폼 비용 $0.021 / µ g 단백질 (노동 및 장비 비용 제외), 경쟁력 있는 가격 상업 키트와 함께 반응 설치의 용이성을 타협 하지 않고 우리의 시스템 만들기. 반응의 µ L 당 비용 비교 평가 표시 비슷한 동향 (Supplemental 그림 1). 우리 수 시작 비용을 견적 ~ 모든 시 약에 대 한 4, 500 달러 그리고 추가적인 $3200는 sonicator 등 특수 장비에 대 한. 이 절차를 완료 하려면 시간 최대 지상에서 모든 시 약 준비 ~ 26 h을 것으로 추정 됩니다. 그러나, 일단 시 약의 큰 주식 준비 되어, 노동에 대 한 요구를 실질적으로 감소. 또한, 플랫폼 경험을 얻은 우리 세포 성장의 크기를 확장 하는 것이 좋습니다, 그리고 준비, 및 시간의 효율성을 극대화 하기 위해 시 약 준비. CFPS 플랫폼 합성 생물학, 높은 처리량 노력, 응용 프로그램에 대 한 권장 시작 비용을 감안할 때, 고 단백질 식 조건에 호환 되지 않습니다 인해 전통적인 단백질 식 플랫폼 셀의 충돌 생화학과 생존 능력 제약입니다. 이러한 특수 경우 원하는 기술을 CFPS 플랫폼을 설정한, vivo에서 식을 통해 CFPS의 큰 비용 정당 이다.
CFPS 플랫폼의 지속적인된 개발은 효소 경로, 생산 및 전통적으로 다루기 힘든 단백질, 비표준 아미노산의 대사 공학 등 생명 공학 노력에 광범위 한 유틸리티 제공 법인 및 부자연 스러운 단백질 표정, 층 화 약 제조, 그리고 줄기 교육64,,6566교실에 실험실을 넘어 확대. 이러한 노력 CFPS 플랫폼의 상세한 특성에 대 한 지속적인 노력에 의해 더 지원 될 것입니다. 세포 추출의 구성의 더 나은 이해는 지속적인된 세련미 향상 된 반응 수율 및 반응 조건61,,6768에 유연성으로 이어질 것입니다.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
저자 들은 아무 경쟁 금융 관심사 또는 다른 충돌을 선언 합니다.
Acknowledgments
저자 박사 제니퍼 VanderKelen, 안드레아 Laubscher, 및 기술 지원에 대 한 토니 Turretto, 웨슬리 Kao, Layne 윌리엄스, 그리고 유용한 토론에 대 한 크리스토퍼 Hight를 인정 하 고 싶습니다. 저자는 또한 자금 빌과 린다 리 펀드, 생명 공학의 쉐 브 론 생명 공학 적용 연구 기금 그랜트, 칼 폴 리 연구, Scholarly, 및 창조적인 활동 그랜트 프로그램 (RSCA 2017), 응용 프로그램에 대 한 센터에서 지원을 인정합니다 그리고 국립 과학 재단 (NSF-1708919). MZL은 캘리포니아 주립 대학 대학원 부여 인정합니다. MCJ는 육군 연구 사무실 W911NF-16-1-0372 인정, 국립 과학 재단 부여 MCB 1413563 및 MCB 1716766는 공군 연구 실험실 센터 우수 그랜트 FA8650-15-2-5518, 방위 위협 감소 기관 보조금 HDTRA1-15-10052/P00001, 데이빗과 루 패 커드 재단, 카밀 드레퓌스 교사 학자 프로그램의 에너지 BER 그랜트 드-SC0018249, 인간 프론티어 과학 프로그램 (RGP0015/2017), 미상 합동 게놈 연구소 ETOP 그랜트, 부서 및 Searle 자금 시카고 커뮤니티 트러스트 지원에 대 한 지원으로 시카고 생명 컨소시엄.
Materials
| Name | Company | Catalog Number | Comments |
| Luria Broth | ThermoFisher | 12795027 | |
| Tryptone | Fisher Bioreagents | 73049-73-7 | |
| Yeast Extract | Fisher Bioreagents | 1/2/8013 | |
| NaCl | Sigma-Aldrich | S3014-1KG | |
| Potassium Phosphate Dibasic | Sigma-Aldrich | 60353-250G | |
| Potassium Phosphate Monobasic | Sigma-Aldrich | P9791-500G | |
| D-Glucose | Sigma-Aldrich | G8270-1KG | |
| KOH | Sigma-Aldrich | P5958-500G | |
| IPTG | Sigma-Aldrich | I6758-1G | |
| Mg(OAc)2 | Sigma-Aldrich | M5661-250G | |
| K(OAc) | Sigma-Aldrich | P1190-1KG | |
| Tris(OAc) | Sigma-Aldrich | T6066-500G | |
| DTT | ThermoFisher | 15508013 | |
| tRNA | Sigma-Aldrich | 10109541001 | |
| Folinic Acid | Sigma-Aldrich | F7878-100MG | |
| NTPs | ThermoFisher | R0481 | |
| Oxalic Acid | Sigma-Aldrich | P0963-100G | |
| NAD | Sigma-Aldrich | N8535-15VL | |
| CoA | Sigma-Aldrich | C3144-25MG | |
| PEP | Sigma-Aldrich | 860077-250MG | |
| K(Glu) | Sigma-Aldrich | G1501-500G | |
| NH4(Glu) | MP Biomedicals | 02180595.1 | |
| Mg(Glu)2 | Sigma-Aldrich | 49605-250G | |
| Spermidine | Sigma-Aldrich | S0266-5G | |
| Putrescine | Sigma-Aldrich | D13208-25G | |
| HEPES | ThermoFisher | 11344041 | |
| Molecular Grade Water | Sigma-Aldrich | 7732-18-5 | |
| L-Aspartic Acid | Sigma-Aldrich | A7219-100G | |
| L-Valine | Sigma-Aldrich | V0500-25G | |
| L-Tryptophan | Sigma-Aldrich | T0254-25G | |
| L-Phenylalanine | Sigma-Aldrich | P2126-100G | |
| L-Isoleucine | Sigma-Aldrich | I2752-25G | |
| L-Leucine | Sigma-Aldrich | L8000-25G | |
| L-Cysteine | Sigma-Aldrich | C7352-25G | |
| L-Methionine | Sigma-Aldrich | M9625-25G | |
| L-Alanine | Sigma-Aldrich | A7627-100G | |
| L-Arginine | Sigma-Aldrich | A8094-25G | |
| L-Asparagine | Sigma-Aldrich | A0884-25G | |
| Glycine | Sigma-Aldrich | G7126-100G | |
| L-Glutamine | Sigma-Aldrich | G3126-250G | |
| L-Histadine | Sigma-Aldrich | H8000-25G | |
| L-Lysine | Sigma-Aldrich | L5501-25G | |
| L-Proline | Sigma-Aldrich | P0380-100G | |
| L-Serine | Sigma-Aldrich | S4500-100G | |
| L-Threonine | Sigma-Aldrich | T8625-25G | |
| L-Tyrosine | Sigma-Aldrich | T3754-100G | |
| Fisherbrand Premium Microcentrifuge Tubes: 2.0 mL | Fisher Scientific | 05-408-138 | |
| Fisherbrand Premium Microcentrifuge Tubes: 1.5 mL | Fisher Scientific | 05-408-129 | |
| Fisherbrand Premium Microcentrifuge Tubes: 0.6 mL | Fisher Scientific | 05-408-120 | |
| PureLink HiPure Plasmid Prep Kit | ThermoFisher | K210007 | |
| Ultrasonic Processor | QSonica | Q125-230V/50Hz | 3.175 mm diameter probe |
| Avanti J-E Centrifuge | Beckman Coulter | 369001 | |
| JLA-8.1000 Rotor | Beckman Coulter | 366754 | |
| 1L Centrifuge Tube | Beckman Coulter | A99028 | |
| Tunair 2.5L Baffeled Shake Flask | Sigma-Aldrich | Z710822 | |
| Microfuge 20 | Beckman Coulter | B30134 | |
| New Brunswick Innova 42/42R Incubator | Eppendorf | M1335-0000 | |
| Cytation 5 | BioTek | ||
| Strep-Tactin XT Starter Kit | IBA | 2-4998-000 | |
| pJL1-sfGFP | Addgene | 69496 | |
| BL21(DE3) | New England BioLabs |
References
- Jiang, L., Zhao, J., Lian, J., Xu, Z. Cell-free protein synthesis enabled rapid prototyping for metabolic engineering and synthetic biology. Synthetic and Systems Biotechnology. 3 (2), 90-96 (2018).
- Carlson, E. D., Gan, R., Hodgman, C. E., Jewett, M. C. Cell-free protein synthesis: Applications come of age. Biotechnology Advances. 30 (5), 1185-1194 (2012).
- Watanabe, M. Cell-Free Protein Synthesis for Structure Determination by X-ray Crystallography. Methods in molecular biology (Clifton, N.J). 607, 149-160 (2010).
- Martemyanov, K. A., Shirokov, V. A., Kurnasov, O. V., Gudkov, A. T., Spirin, A. S. Cell-Free Production of Biologically Active Polypeptides: Application to the Synthesis of Antibacterial Peptide Cecropin. Protein Expression and Purification. 21 (3), 456-461 (2001).
- Renesto, P., Raoult, D. From genes to proteins: in vitro expression of rickettsial proteins. Annals of the New York Academy of Sciences. 990, 642-652 (2003).
- Xu, Z., Chen, H., Yin, X., Xu, N., Cen, P. High-Level Expression of Soluble Human b-Defensin-2 Fused With Green Fluorescent Protein in Escherichia coli Cell-Free System. Applied Biochemistry and Biotechnology. 127 (1), 053-062 (2005).
- Baumann, A. In-situ observation of membrane protein folding during cell-free expression. PLoS ONE. 11 (3), 1-15 (2016).
- Wang, Y., Percival, Y. H. P. Cell-free protein synthesis energized by slowly-metabolized maltodextrin. BMC Biotechnology. 9, 1-8 (2009).
- Whittaker, J. W. Cell-free protein synthesis: the state of the art. Biotechnology Letters. 35 (2), 143-152 (2013).
- Martin, R. W. Cell-free protein synthesis from genomically recoded bacteria enables multisite incorporation of noncanonical amino acids. Nature Communications. 9 (1), 1203 (2018).
- Kwon, Y. C., Song, J. K., Kim, D. M. Cloning-Independent Expression and Screening of Enzymes Using Cell-Free Protein Synthesis Systems. Methods in Molecular Biology. 1118, 97-108 (2014).
- Chappell, J., Jensen, K., Freemont, P. S. Validation of an entirely in vitro approach for rapid prototyping of DNA regulatory elements for synthetic biology. Nucleic Acids Research. 41 (5), 3471-3481 (2013).
- Takahashi, M. K. Characterizing and prototyping genetic networks with cell-free transcription-translation reactions. Methods. 86, 60-72 (2015).
- Karim, A. S., Jewett, M. C. A cell-free framework for rapid biosynthetic pathway prototyping and enzyme discovery. Metabolic Engineering. 36, 116-126 (2016).
- Dudley, Q. M., Anderson, K. C., Jewett, M. C. Cell-Free Mixing of Escherichia coli Crude Extracts to Prototype and Rationally Engineer High-Titer Mevalonate Synthesis. ACS Synthetic Biology. 5 (12), 1578-1588 (2016).
- Pardee, K. Paper-based synthetic gene networks. Cell. 159 (4), 940-954 (2014).
- Zawada, J. F. Microscale to manufacturing scale-up of cell-free cytokine production-a new approach for shortening protein production development timelines. Biotechnology and Bioengineering. 108 (7), 1570-1578 (2011).
- Sullivan, C. J. A cell-free expression and purification process for rapid production of protein biologics. Biotechnology Journal. 11 (2), 238-248 (2016).
- Li, J. Cell-free protein synthesis enables high yielding synthesis of an active multicopper oxidase. Biotechnology Journal. 11 (2), 212-218 (2016).
- Heinzelman, P., Schoborg, J. A., Jewett, M. C. pH responsive granulocyte colony-stimulating factor variants with implications for treating Alzheimer's disease and other central nervous system disorders. Protein Engineering Design and Selection. 28 (10), 481-489 (2015).
- Pardee, K. Low-Cost Detection of Zika Virus Using Programmable Biomolecular Components. Cell. 165 (5), 1255-1266 (2016).
- Slomovic, S., Pardee, K., Collins, J. J. Synthetic biology devices for in vitro and in vivo diagnostics. Proceedings of the National Academy of Sciences. 112 (47), 14429-14435 (2015).
- Gootenberg, J. S. Nucleic acid detection with CRISPR-Cas13a/C2c2. Science. 356 (6336), 438-442 (2017).
- Pardee, K. Portable, On-Demand Biomolecular Manufacturing. Cell. 167 (1), 248-259 (2016).
- Karig, D. K., Bessling, S., Thielen, P., Zhang, S., Wolfe, J. Preservation of protein expression systems at elevated temperatures for portable therapeutic production. Journal of the Royal Society, Interface. 14 (129), (2017).
- Smith, M. T., Berkheimer, S. D., Werner, C. J., Bundy, B. C. Lyophilized Escherichia coli-based cell-free systems for robust, high-density, long-term storage. BioTechniques. 56 (4), 186-193 (2014).
- Hunt, J. P., Yang, S. O., Wilding, K. M., Bundy, B. C. The growing impact of lyophilized cell-free protein expression systems. Bioengineered. 8 (4), 325-330 (2017).
- Stark, J. C. BioBitsTM Bright: A fluorescent synthetic biology education kit. Science Advances. 4 (8), (2018).
- Huang, A. BioBitsTM Explorer: A modular synthetic biology education kit. Science Advances. 4 (8), (2018).
- Zemella, A., Thoring, L., Hoffmeister, C., Kubick, S. Cell-Free Protein Synthesis: Pros and Cons of Prokaryotic and Eukaryotic Systems. ChemBioChem. 16 (17), 2420-2431 (2015).
- Oza, J. P. Robust production of recombinant phosphoproteins using cell-free protein synthesis. Nature Communications. 6 (1), 8168 (2015).
- Zemella, A. Cell-free protein synthesis as a novel tool for directed glycoengineering of active erythropoietin. Scientific Reports. 8 (1), 8514 (2018).
- Jaroentomeechai, T. Single-pot glycoprotein biosynthesis using a cell-free transcription-translation system enriched with glycosylation machinery. Nature Communications. 9 (1), 2686 (2018).
- Kightlinger, W. Design of glycosylation sites by rapid synthesis and analysis of glycosyltransferases. Nature Chemical Biology. 14 (6), 627-635 (2018).
- Schoborg, J. A. A cell-free platform for rapid synthesis and testing of active oligosaccharyltransferases. Biotechnology and Bioengineering. 115 (3), 739-750 (2018).
- Chekulayeva, M. N., Kurnasov, O. V., Shirokov, V. A., Spirin, A. S. Continuous-Exchange Cell-Free Protein-Synthesizing System: Synthesis of HIV-1 Antigen Nef. Biochemical and Biophysical Research Communications. 280 (3), 914-917 (2001).
- Hong, S. H. Improving Cell-Free Protein Synthesis through Genome Engineering of Escherichia coli Lacking Release Factor 1. ChemBioChem. 16 (5), 844-853 (2015).
- Endo, Y., Otsuzuki, S., Ito, K., Miura, K. Production of an enzymatic active protein using a continuous flow cell-free translation system. Journal of Biotechnology. 25 (3), 221-230 (1992).
- Volyanik, E. V., Dalley, A., Mckay, I. A., Leigh, I., Williams, N. S., Bustin, S. A. Synthesis of Preparative Amounts of Biologically Active Interleukin-6 Using a Continuous-Flow Cell-Free Translation System. Analytical Biochemistry. 214 (1), 289-294 (1993).
- Martin, G. A., Kawaguchi, R., Lam, Y., DeGiovanni, A., Fukushima, M., Mutter, W. High-yield, in vitro protein expression using a continuous-exchange, coupled transcription/ translation system. BioTechniques. 31 (4), 948-950 (2001).
- Stech, M., Quast, R. B., Sachse, R., Schulze, C., Wüstenhagen, D. A., Kubick, S. A Continuous-Exchange Cell-Free Protein Synthesis System Based on Extracts from Cultured Insect Cells. PLoS ONE. 9 (5), e96635 (2014).
- Quast, R. B., Sonnabend, A., Stech, M., Wüstenhagen, D. A., Kubick, S. High-yield cell-free synthesis of human EGFR by IRES-mediated protein translation in a continuous exchange cell-free reaction format. Scientific Reports. 6 (1), 30399 (2016).
- Thoring, L., Dondapati, S. K., Stech, M., Wüstenhagen, D. A., Kubick, S. High-yield production of "difficult-to-express" proteins in a continuous exchange cell-free system based on CHO cell lysates. Scientific Reports. 7 (1), 11710 (2017).
- Hoffmann, M., Nemetz, C., Madin, K., Buchberger, B. Rapid translation system: A novel cell-free way from gene to protein. Biotechnology annual review. 10, 1-30 (2004).
- Kwon, Y. C., Jewett, M. C. High-throughput preparation methods of crude extract for robust cell-free protein synthesis. Scientific Reports. 5 (1), 8663 (2015).
- Katsura, K. A reproducible and scalable procedure for preparing bacterial extracts for cell-free protein synthesis. Journal of Biochemistry. 162 (June), 357-369 (2017).
- Fujiwara, K., Doi, N. Biochemical preparation of cell extract for cell-free protein synthesis without physical disruption. PLoS ONE. 11 (4), 1-15 (2016).
- Shrestha, P., Holland, T. M., Bundy, B. C. Streamlined extract preparation for Escherichia coli-based cell-free protein synthesis by sonication or bead vortex mixing. BioTechniques. 53 (3), 163-174 (2012).
- Krinsky, N. A Simple and Rapid Method for Preparing a Cell-Free Bacterial Lysate for Protein Synthesis. PLoS ONE. 11 (10), (2016).
- Jewett, M. C., Swartz, J. R. Mimicking the Escherichia coli cytoplasmic environment activates long-lived and efficient cell-free protein synthesis. Biotechnology and Bioengineering. 86 (1), 19-26 (2004).
- Swartz, J. R., Jewett, M. C., Woodrow, K. A. Cell-Free Protein Synthesis With Prokaryotic Combined Transcription-Translation. Recombinant Gene Expression. (267), 169-182 (2004).
- Voloshin, A. M., Swartz, J. R. Efficient and scalable method for scaling up cell free protein synthesis in batch mode. Biotechnology and Bioengineering. 91 (4), 516-521 (2005).
- Vernon, W. B. The role of magnesium in nucleic-acid and protein metabolism. Magnesium. 7 (5-6), 234-248 (1988).
- Pratt, J. M. Transcription and Translation: A Practical Approach. , IRL Press. New York. (1984).
- Kim, D. M., Kigawa, T., Choi, C. Y., Yokoyama, S. A Highly Efficient Cell-Free Protein Synthesis System from Escherichia coli. European Journal of Biochemistry. 239 (3), 881-886 (1996).
- Shin, J., Noireaux, V. Efficient cell-free expression with the endogenous E. Coli RNA polymerase and sigma factor 70. Journal of Biological Engineering. 4 (1), 8 (2010).
- Zubay, G. In Vitro Synthesis of Protein in Microbial Systems. Annual Review of Genetics. 7 (1), 267-287 (1973).
- Kigawa, T. Preparation of Escherichia coli cell extract for highly productive cell-free protein expression. Journal of Structural and Functional Genomics. 5 (1/2), 63-68 (2004).
- Liu, D. V., Zawada, J. F., Swartz, J. R. Streamlining Escherichia Coli S30 Extract Preparation for Economical Cell-Free Protein Synthesis. Biotechnology Progress. 21 (2), 460-465 (2008).
- Yang, W. C., Patel, K. G., Wong, H. E., Swartz, J. R. Simplifying and streamlining Escherichia coli-based cell-free protein synthesis. Biotechnology Progress. 28 (2), 413-420 (2012).
- Foshag, D. The E. coli S30 lysate proteome: A prototype for cell-free protein production. New Biotechnology. 40 (Pt B), 245-260 (2018).
- Caschera, F. Bacterial cell-free expression technology to in vitro systems engineering and optimization. Synthetic and Systems Biotechnology. 2 (2), 97-104 (2017).
- Chizzolini, F., Forlin, M., Yeh Martín, N., Berloffa, G., Cecchi, D., Mansy, S. S. Cell-Free Translation Is More Variable than Transcription. ACS Synthetic Biology. 6 (4), 638-647 (2017).
- Hong, S. H., Kwon, Y. C., Jewett, M. C. Non-standard amino acid incorporation into proteins using Escherichia coli cell-free protein synthesis. Frontiers in Chemistry. 2, 34 (2014).
- Sawasaki, T., Ogasawara, T., Morishita, R., Endo, Y. A cell-free protein synthesis system for high-throughput proteomics. Proceedings of the National Academy of Sciences of the United States of America. 99 (23), 14652-14657 (2002).
- Dudley, Q. M., Karim, A. S., Jewett, M. C. Cell-free metabolic engineering: Biomanufacturing beyond the cell. Biotechnology Journal. 10 (1), 69-82 (2015).
- Garcia, D. C. Elucidating the potential of crude cell extracts for producing pyruvate from glucose. Synthetic Biology. 3 (1), 1-9 (2018).
- Hurst, G. B. Proteomics-Based Tools for Evaluation of Cell-Free Protein Synthesis. Analytical Chemistry. 89 (21), 11443-11451 (2017).

{kind=link}
{kind=link}
{kind=link}