

Summary
Hier stellen wir einen kombinatorischen Ansatz vor, bei dem hochauflösende Mikroskopie, Rechenwerkzeuge und Einzellen-Beschriftungen in lebenden C. eleganischen Embryonen verwendet werden, um die Einzelzellendynamik während der Neuroentwicklung zu verstehen.
Abstract
Caenorhabditis elegans (C. elegans) sticht als einziger Organismus hervor, in dem die Herausforderung, die zellulären Ursprünge eines gesamten Nervensystems zu verstehen, mit einer einzigen Zellauflösung in vivo beobachtet werden kann. Hier stellen wir ein integriertes Protokoll zur Untersuchung der Neuroentwicklung in C. eleganischen Embryonen vor. Unser Protokoll kombiniert die Bildgebung, die Linienführung und die neuroanatomische Verfolgung einzelner Zellen bei der Entwicklung von Embryonen. Durch den Einsatz der Dual-View-Invertitionslinge-Illuminationsmikroskopie (DiSPIM) erreichen wir eine langfristige, vierdimensionale (4D) Bildgebung lebender C . eleganischer Embryonen mit nahezu isotropher räumlicher Auflösung. Nuklei und neuronale Strukturen in den Nematoden Embryonen werden abgebildet und isotropical verschmolzen, um Bilder mit einer Auflösung von ~ 330 nm in allen drei Dimensionen zu erzeugen. Diese minutenlangen hochauflösenden 4D-Datensätze werden dann analysiert, um definitive zelllineare Identitäten mit Genexpression und morphologischer Dynamik auf einzelligen und subzellulären Detailebenen zu korrelieren. Unser Protokoll ist so strukturiert, dass es eine modulare Umsetzung der einzelnen beschriebenen Schritte ermöglicht und Studien zu Embryogenese, Genexpression oder Neuroentwicklung verbessert werden kann.
Introduction
C. elegans sticht als einziger Organismus hervor, in dem jede Zelle im Embryo während der gesamten Neuroentwicklung beobachtet werden kann. Mit der gesamten Zelllinie bekannt und invariant1, und mit der Entwicklung neuer Werkzeuge, die die Kennzeichnung und kontinuierliche Abbildung von einzelnen Zellen in Embryonen ermöglichen, können Biologen nun beginnen, verschiedene Schritte in der Entwicklung der Nematoden nervös zu untersuchen System aus allen Blickwinkeln-Zellgeburt; Migration und Differenzierung; NeuritBildung, gezieltes Auswachsen und Faschisierung; Synapsbildung; Und Abstimmung von Funktionskreisen. Die Erfassung der neuronalen Auswuchsdynamik im C. eleganischen Embryo durch die Kombination von stabil ausgedrückten Reportern und Fluoreszenzmikroskopie ist für die wissenschaftliche Gemeinschaft wertvoll.
Die Entwicklungsstudien in C. elegans nutzen oft die invarianten Zellabstammung und die Karten des Zellschicks dieser Art, um das kontextbezogene Verständnis auf der Einzeller-Ebene imintakten Organismus zu erweitern. Die Auto-Line-Age-Analyse-mit der Software StarryNite2,3, 4 und AceTree5,6, 7, 8-profitiertvon hohem Kontrast,hoher Auflösung Bilder von fluoreszierenden Kernen. Um optimal zu arbeiten, hängt StarryNite/AceTree auch von einer vorhersehbaren eingeschränkten Orientierung der abgebildeten Embryonen während der Entwicklung ab. Die konfokale Mikroskopie, die in C. eleganischem Embryonen, die zwischen zwei Coverslips komprimiert werden, durchgeführt wird, ist seit mehr als einem Jahrzehnt die Standardmethode der Autolinesmikroskopie, weil sie sowohl eine hohe kontrast-/hohe Auflösung als auch eine vorhersehbare Beschränkung bietet. Ausrichtung des Embryos7,8. Wir haben zuvor den Bau und die Verwendung eines neuartigen Lichtbogen-Basis, das selektive Flächenbeleuchtungsmikroskop (diSPIM) für die Live-Probenbildgebungwie C. elegans embryogenesis 9, 10 verwendet . , 11 , 12 , 13. Die Lichtbogenmikroskopie bietet in der Regel eine geringe Phototoxizität, hohe Geschwindigkeit und eine langfristige Abbildung von lebenden 3D-Exemplaren14,15. Die DiSPIM-Methode erzeugt insbesondere vierdimensionale (4D) Bilder mit einer nahezu isotropen Raumauflösung von etwa 330 nm9.
Im Vergleich zur konfokalen Mikroskopie bietet diSPIM eine höhere Signal-zu-Rausch-und-geschwindigkeit, eine isotropere Raumauflösung und eignet sich besser für die langfristige Bildgebung16 . Deshalb haben wir daran gearbeitet, die diSPIM-Daten für die Eingabe in StarryNite/AceTree anzupassen und zu untersuchen, ob dies die Linienanalysen verbessern würde. Eine große Hürde ist, dass DiSPIM-Exemplare nicht einfach durch Auberginen-Kompression eingeschränkt werden können, um erwartete Orientierungen für StarryNiteTree zu übernehmen. Die zufällige Ausrichtung der Zellpositionen in dem analysierten Volumen mindert die Genauigkeit der Auto-Line-Analyse.
Wir haben daher CytoSHOW, eine von den Seiten geführte Benutzeroberfläche, eingesetzt, die es Nutzern ermöglicht, bei der Vorverarbeitung von DiSPIM-Bildern eine präzise 3D-Ausrichtung von Embryonen auszuwählen, die sowohl qualitätsoptimierte als auch kontextbewusste Bilddaten für den Einstieg in StarryNite liefert. /AceTree. Bei der Nutzerauswahl von abgebildeten Embryonen inszeniert CytoSHOW eine automatisierte Datenverarbeitungs-Pipeline. Eingeschreifte und rückensubtrahierte Embryo-Bilder werden in TIFF-Stapeldateien für jede Position, jeden Zeitpunkt und jede Ansicht gespeichert. CytoSHOW ruft dann iterativ das Programm SpimFusion auf, die beiden vorverarbeiteten Ansichten gemeinsam zu registrieren und gemeinsam zu dekonvolvieren, indem er den Richardson-Lucy 17,18Algorithmus verwendet, um isotrope hochauflösende volumetrische Bilder zu liefern. Für StarryNite wurde ein diSPIM-spezifischer Parametersatz optimiert, um sein Verhalten während der Bildsegmentierung und der Nukleus-Tracking in isotropical verschmolzenen Bildern zu steuern. Geschmolzene Bilder und Liniengebnisse werden dann mit AceTree bearbeitet, was es dem Benutzer ermöglicht, Fehler in der von StarryNite generierten Auto-Linie zu erkennen und zu beheben. AceTree kann auch Line-Baum-und 3D-modellierte Renderings von geprägten Kernen im Embryo präsentieren. Wir stellen fest, dass die Geschwindigkeit und Genauigkeit der automatischen Linie durch isotropical verschmolzene Bilder deutlich verbessert wird, im Vergleich zu Rohbildern der beiden SPIM-Kameras. Unser Protokoll, das für die hier beschriebene C . elegans-Anwendung optimiert wurde, könnte in der Regel für die automatische Abfertigung von DiSPIM-Daten, die für andere Arten oder Proben produziert werden, angepasst werden. Wenn es sich um die beabsichtigte Verwendung des Protokolls handelt, beachten Sie bitte, dass für neue Exemplare eine zusätzliche Abstimmung der StarryNite-Parameter erforderlich ist, wie beschrieben3,4.
Die erfolgreiche Umsetzung dieses Protokolls führt zu Bildern mit 4D-isotropen Auflösung und ermöglicht Biologen, Zelllinien zu verfolgen und gleichzeitig Neuronen im sich entwickelnden C. eleganischen Embryo zu identifizieren und zu analysieren. Darüber hinaus können wir durch die Verschmelzung mehrerer nachverarbeiteter Algorithmen-wobei die Hardwarebeschleunigung die zeitaufwändigste dieser Algorithmen ist-nun sowohl feine subzelluläre Details als auch die Zelllinien und Zellverschlüsse lebender Embryonen in einer im Wesentlichen realen Zeit analysieren. Dieses neue Protokoll ermöglicht eine präzise, informierte Manipulation und Beobachtung des Zellverhaltens während der probativen Studien der Differenzierung und Morphogenese in vivo. In diesem Manuskript stellen wir eine detaillierte Erklärung der verbesserten Protokolle vor, die wir für die Linienführung und die Zellverfolgung in der Entwicklung von C. eleganischem Embryonen entwickelt haben, um Studien zur Embryogenese, Genexpression oder Neuroentwicklung zu verbessern.
Protocol
1. Montage der DiSPIM-Stahlbildkammer mit Poly-Lysin-beschichtetem Coverslip
Hinweis: Die folgenden Schritte sind alle erforderlich, um die Line-Age-Analyse von Nematoden Embryonen durch StarryNite/AceTree zu optimieren und zu automatisieren. Für Experimente, die eine Verfolgung von C. elegans Zelllinien erfordern, können mehrere Optionen (die durch solche angezeigt werden) weggelassen werden.
- Zeichnen Sie ein kleines Rechteck (2mm x 5mm) in der Mitte eines sauberen rechteckigen Kenterlips (Nr. 1,5, 24 mm x 50 mm) mit einem Schärding (oder ähnlichem Stift).
- Drehen Sie den Kerkel auf die unmarkierte Seite um und legen Sie 10 μL Poly-L-Lysin (Kat. Nr. P1524) über das markierte Rechteck.
Hinweis: Machen Sie eine Arbeitskonzentration von 1 mg/mL Poly-L-Lysin, die sich in gefiltertem Wasser (oder gleichwertig) aufgelöst. Für die langfristige Lagerung, bereiten Sie 5 – 10 mL Aliquots vor und lagern Sie bei-20 ° C. Nach dem Auftauen können die Aliquots 3 – 4 Wochen bei Raumtemperatur (23 ° C) gelagert werden. - Den Poly-L-Lysin 5 Minuten lang den Kitzelzug anziehen lassen (Abbildung 1A). Poly-L-Lysin wird für die Beschichtung von Glasabdeckungsschlitzen verwendet, bei denen die Embryonen montiert werden, so dass die Embryoneneschale fest an der Koppel haften kann, auch wenn die Ziele in das Bildmedium eingetaucht sind.
- Legen Sie den Poly-Lysin-beschichteten Kopfsteinpflap in die untere Hälfte der Stahlbildkammer.
- Die obere Hälfte der Stahlbildkammer mit dem Kitzel auf die untere Hälfte legen und mit den vier Schrauben, die mit der Kammer verbunden sind, festziehen. Überprüfen Sie visuell von der Seite, um sicherzustellen, dass die obere Hälfte gleichmäßig in der unteren Hälfte sitzt (Abbildung 1B).
- Füllen Sie die Kammer mit 7 – 8 mL M9 Puffer 19, einem isotonischen Puffer, der verhindert, dass Embryonen im frühen Stadium dem abnormalen osmotischen Druck nachgeben. 1-Zell-und sogar 2-Zellen-Embryonen sind osmotisch empfindlich und können sich in diesem isotonischen Puffer ungewöhnlich entwickeln. Embryonen neigen auch dazu, sich in der dreifachen Phase zu verhaften, wenn M9 durch Wasser ersetzt wird.
2. Zubereitung von C. elegans Embryoproben für die Montage
Hinweis: Etwa 18 Stunden vor der Bildgebung werden fünf junge (1 Tage seit dem letzten Molt) Erwachsenen und zehn Larvenstadien 4 (L4) C. elegans auf eine mit E . coli-Stamm gesäten Agar-Platte (NGM) Agar Platte gepflückt. Platin Drahtpicks werden verwendet, um Larven und junge erwachsene C. elegans zu bewegen, ohne das Tierzuschädigen 19.
- Bereiten Sie eine 1% Methylzellulose (Kat. Nr. H7509-25G) Lösung im M9-Puffer.
Achtung: Methylzellulose muss in warmem M9 gerührt werden, bis sie aufgelöst ist. Einmal vorbereitet, kann diese Lösung bei Raumtemperatur gelagert werden. - 500 μL von 1% Methylzellulose-in-M9-Lösung in die Depression einer konkaven Mikroskop-Rutsche einfügen. Dieser viskose Puffer wird bei zwei Gelegenheiten verwendet: 1) bei der Ernte von frühen Embryonen durch den Schnitt erwachsener Würmer und 2) beim Waschen von Embryonen, die im Spätstadium von einer NGM-Platte abgeholt werden.
NOTE: Methylzellulose wird verwendet, um zu verhindern, dass Embryonen an der Mikroskope rutschen. - Für die Abbildung von Embryonen im Spätstadium werden C . eleganische Embryonen (vorzugsweise von einer NGM-Platte mit jungen Erwachsenen) mit einem Wimpernpick abgelegt und die Embryonen auf die 1% Methylzellulose auf dem konkaven Mikroskop-Dia gebracht. Der Wimpernpflücker hilft, die Kraft zu reduzieren und so Stress oder Beschädigungen der Embryonen während der Handhabung zu minimieren. Die Prozedur, um einen Wimpernpick zu machen, wird von Hart20abgedeckt.
- Mit einem zweiten Wimpernpflücker (in der gegenüberliegenden Hand) beide Wimpern vorsichtig zusammenzapfen, um die Embryonen in der Methylzellulose auszusetzen.
- Optional: Wenn man plant, embryonale Zellen mit StarryNite zu beinien, muss man 1-Zell-bis 4-Zell-Embryonen montieren. Dazu wählen Sie zunächst junge Erwachsene aus einer NGM-Platte aus und schieben sie mit einem Platin-Draht-Pick in die M9-Methylzellulose-Lösung auf der konkaven Mikroskop-Rutsche.
- Optional: Mit den geschärften Spitzen hypodermischer Nadeln (Nr. 18G x 1 1/2) schneiden Sie das Tier quer am Mittelkörper ab, um 1-Zell-bis 4-Zellen-Embryonen freizusetzen.
3. Die Mundpipette: Assembling-Aspirator-Röhre mit Mikrokapillarpipette
Hinweis: Wir verwenden ein Saugrohr mit einer handgezogenen Mikrokapillarpfeipette, die in die Gummidichtung des Rohres eingelegt ist. So können wir Embryonen vom Trennrutsch auf die Poly-L-Lysin-beschichtete Oberfläche in der Pufferkammer übertragen.
- Manuell ziehen Sie die Mikrokapillarpipette über eine offene Flamme, um zwei Hälften mit gestreckten Spitzen zu schaffen.
- Nehmen Sie die Hälfte der Mikrokapillarpipette und legen Sie das stumpfe Ende in die Gummidichtung des Saugerrohres (Abbildung 1C). Stellen Sie die andere Hälfte der Mikrokapillarpipette beiseite, um sie später (falls nötig) zu verwenden.
- Mit dem mit Mikrokapillarpipette ausgestatteten Saugrohr brechen Sie sanft die Spitze der mikrokapillaren Pipette und bilden eine Öffnung, die zu etwa 1 – 2 zwei Embryonen passt (von hier aus wird dieses Instrument als "Mundpipette" bezeichnet).
4. Montage C. elegans Embryonen auf Poly-Lysin beschichteten Coverslip
- Mit dem Saug-Mundstück sanft zwischen den Zähnen gehalten, die mikrokapillare Pipette mit 10 – 15 μL M9-Buffer vorfüllen und dann mehrere Embryonen aus dem Konkave sanft in die Kapillare saugen.
- Übertragen Sie die Embryonen auf die mit M9-Buffer gefüllte Stahlbildkammer und positionieren Sie die Kapillarspitze so, dass die Embryonen in das zentrale Rechteck des Kenterlips fallen.
- Vermeiden Sie Verletzungen der Embryonen, bewegen Sie sie sanft mit einem Wimpernpick oder verwenden Sie die Mundpipette, um die Embryonen vertikal zu positionieren, um die Embryonen so zu orientieren, dass die lange Achse des Embryos senkrecht zur langen Achse des Coverslips ist (Abbildung 1B inset , untere Tafel).
Hinweis: Die Positionierung des Embryos in dieser Ausrichtung minimiert die Anzahl der Bildscheiben, wodurch die Lichtdosierung und die Datenverarbeitungszeit verkürzt werden und gleichzeitig die Aufnahmegeschwindigkeit verbessert wird. - Legen Sie die Stahlbildkammer auf der Mikroskop-Stufe in den Probenhalter (Abbildung 1D).
5. Montage, Software-Einrichtung und Laseroptimierung für die embryonale Bildgebung mit dem diSPIM
- Siehe Schritt für Schritt Anweisungen, wie man die gesamte fasergekoppelte DiSPIM aus kommerziell erhältlichen Teilen in Kumar et al.10,11 und an http://www.dispim.org zusammenbaut. Ein Video-Protokoll zur Montage des DiSPIM ist auch auf der ASI-Website (http://www.asiimaging.com) verfügbar.
Hinweis: Das Instrumenten-Setup für dieses Protokoll ist identisch mit Kumar etal. 10,11 diSPIM,das 40x 0,8NA Wassereintauchlinsen für die Bildgebung verwendet. Der einzige Unterschied zwischen dem Setup in diesem Protokoll und Kumar et al.10,11 ist die Zugabe eines dichromen Spiegels (Spalten auf 560 nm) und roten und grünen Bandpass-Filtern in einem bildgebenden Spaltgerät (Modell A12801-01), das auf einem installierten Bildverteilgerät (Modell A12801-01) installiert ist. Beide diSPIM-Abbildungsarme. Die Addition der Bildspaltoptik ermöglicht die gleichzeitige Erfassung von Bildern aus zwei verschiedenen Fluorophoren – die durch 561 nm und 488 nm Laser – durch die Trennung der Emissionsbänder auf zwei Hälften desselben Kamerapiches angeregt werden. - Überprüfen Sie nach der Instrumentenmontage die optische Ausrichtung des DiSPIM vor der Bildgebung.
Hinweis: Um die DiSPIM richtig auszurichten, sehen Sie https://youtu.be/qnOrg30NNuE, und für Hardware-Informationen, http://dispim.org/hardware/objectives und http://www.asiimaging.com. - Nutzen Sie die Open-Source-Plattform Micro-Manager(https://micro-manager.org/) 21, die für den Betrieb von Lichtblech-Mikroskopen für die Hochdurchsatz-Zellgebung22optimiert wurde. Wir empfehlen die Verwendung des ASI diSPIM-Plugins für die Mehrpositionserfassung, das eine gleichzeitige Abbildung von bis zu 30 Embryonen wiebeschrieben23 ermöglicht.
- Mit dem Micro-Manager open, set laser intensities to ~ 179 μW (0,5) for 488 nm und ~ 79 μW (0,25) für 561 nm (Abbildung 2A, rotes Rechteck).
Hinweis: Dies sind empfohlene Einstellungen für die langfristige Abbildung von C. eleganischen Embryonen in 1-Minuten-Intervallen. Bei der Langzeit-Dual-Farben-Bildgebung wird der 561 nm-Laser verwendet, um Kerne (mCherry:: Histone) abzubilden, bis sich Embryonen im Bohnenstadium befinden, an dem der 488 nm-Laser dann eingeschaltet wird, um auch die GFP-beschrifteten Neuronen abzubilden. Diese bildgebenden Bedingungen sind optimiert, um die Phototoxizität zu minimieren und das Überleben und Schlüpfen der Embryonen zu sichern, während sie eine kontinuierliche (12 – 14 Stunden) kontinuierliche Erfassung von neuroentwicklungs-und linearnden Daten ermöglichen. - Wählen Sie im Micro-Manager das Menü Plugins > Device Control > ASI diSPIM , um das ASI-DiSPIM-Fenster zu öffnen (Abbildung 2B). Wählen Sie die Akquisition Tab. In der Sektion " Dateneinsparung" (grünes Rechteck), der Volumeneinstellungen (blaues Quadrat) und der Slice Settings (orangefarbenes Quadrat) stellen Sie sicher, dass jeder Parameter wie in Abbildung 2B dargestellt gesetzt wird.
NOTE: Unsere Bildanalyse-Software CytoSHOW ist an die Arbeit mit anderen optionalen Ausgabedatenformaten angepasst, wie zum Beispiel mit der in Micro-Manager eingebauten, paketverketteten OME-TIFF-Dateireihe und der TIFF-Stack-Dateireihe, die nach der Erfassung durch die Verwendung einer Exportfunktion in Micro-Manager erstellt wurden. Typischerweise wird das mit einem Zeitstapel zusammengefollte OME-TIFF-Dateidatenformat verwendet, weil es die Echtzeitbetrachtung und Verarbeitung des Bildvolumens ermöglicht, sobald die Rohdaten erfasst sind.
6. Optimierte Autofokus-Parameter für die langfristige Abbildung von C. eleganischen Embryonen
- Setzen Sie Micro-Manager-Autofokus-Parameter auf die Einstellungen, die für die langfristige Qualität von DiSPIM-Bildgebung von C. elegans Embryonen optimiert sind. Klicken Sie im ASI-DiSPIM-Fenster auf das Autofokus-Tab (Abbildung 2C). In der Sektion Allgemeine Autofokus-Optionen (schwarzes Quadrat) geben Sie die Parameter genau wie gezeigt an. Beachten Sie, dass der Autofokus-Kanal (rotes Quadrat) Ihren Nuklearkanal Fluoreszenzkanal in Linienexperimenten angeben sollte.
Hinweis: Wenn der maximale Offset größer als 5 μm ist, driften die Bilder meist aus dem Fokus. - Klicken Sie, Plugins > Acquisition Tools > Pattern Overlay.
- Klicken Sie im Pattern-Overlay-Fenster auf "Gitter anzeigen".
- Klicken Sie im ASI-DiSPIM-Fenster auf die Registerkarte Navigation .
- Klicken Sie auf Kästchen für Beam und Blatt von Path A oder B, dann klicken Sie auf Live. Die Bildaufnahme beginnt. Ein Live-View-Fenster öffnet sich. Wählen Sie die Autofokus-Analyseregion des Embryos aus, indem Sie eine Box um den Embryo auf dem ausgewählten Kanal von 6.1 zeichnen.
Hinweis: Wir erfassen in der Regel 420 Zeitpunkte für 10 Embryonen pro Bildgebungssitzung. Die Rohdaten pro Bildstunde sind in der Regel 1,7TB, während die Daten von Starrynite dekonditiert und die Starrynite-verarbeiteten Daten 1,4TB betragen (siehe Schritte 9 und 10). Wir empfehlen die Verwendung von GroßraumHDD (18TB auf unserem aktuellen System) für die Bildaufnahme und Cloud-Plattformen für die Bildspeicherung. - Klicken Sie auf die Akquisition im "Akquisitionstabelle ", um mit der langfristigen multidimensionalen Bildaufnahme zu beginnen (Abbildung 2B).
7. Öffnen roher Micro-Manager-Bilder in CytoSHOW
- Laden Sie das Software-Paket von http://dispimlineage.wormguides.org herunter.
Hinweis: Das Software-Paket wird als .zip-Datei heruntergeladen und muss auf die "C:\" extrahiert werden. Verzeichnis vor der Verwendung. Weitere Details zur Installation finden Sie auf http://dispimlineage.wormguides.org/diSPIMlineaging_InstallationInstructions.htm. - Doppelklicken Sie auf die Datei C:\CytoSHOWextrasForC\CytoSHOW_APP.jnlp, um mit CytoSHOW zu beginnen.
- Wählen Sie das Menü Datei > Neu > diSPIM-Monitor (Micro-Manager). Suchen Sie den Root-Datenstand-Ordner, in dem die Erfassungszeitpunkt-Ordner gespeichert wurden. Wählen Sie einen beliebigen Zeitpunkt aus und klicken Sie auf Öffnen. Mehrdimensionale Navigationsfenster ( DiSPIM-Monitorfenster genannt) werden sowohl für SPIMA als auch für SPIMB automatisch geöffnet (Abbildung 3A).
Hinweis: Diese Fenster überwachen den Root-Datenordner für neu gespeicherte Rohzeit-Stacks (falls ein Exemplar noch aufgenommen wird). Nachdem jeder neue Timepoint erworben ist, wird jedes der Fenster, die die einzelnen SPIM-Arme und Probenpositionen überwachen, aktualisiert, um den gesamten Multichannel-4D-Datensatz für jeden Embryo anzuzeigen.
8. Erzeugung von Max-Projektionsbildern mit CytoSHOW
Schon vor der Dekonvolution können die Rohdaten schnell verarbeitet werden, um die globalen Eigenschaften des Probens zu bewerten.
- Klicken Sie auf die Z-MIP-Taste auf der linken Seite des Bildfensters (Abbildung 3A, rotes Rechteck), um maximale Intensitätsprojektionen durch die volle Tiefe und den Vollzeitverlauf einer bestimmten Position oder eines SPIM-Arms zu erstellen. Ein Z-Projektion-Hyperstack-Fenster wird angezeigt.
- Im Z-Projektione-Hyperstack-Fenster wird der Projektionstyp auf die Max-Intensitäteingestellt. Geben Sie an, welche Kanäle, Scheiben und Zeit-Point-Frames auf der Grundlage der Benutzerpräferenz verarbeitet werden sollen.
- Klicken Sie auf OK, wenn Sie fertig sind.
- Wählen Sie den Ordner-Standort aus , um die Max-Intensitätsausgänge aus dem Dateidialogfenster zu speichern, und klicken Sie dann auf OK. Lassen Sie etwas Zeit (15 – 20 Minuten, je nach Größe des Datensatzes und der Rechenleistung des Computers) für CytoSHOW, um Projektionsbilder zu erzeugen.
9. Analyse von Zelllinien in isotropen hochauflösenden Volumendaten
- Optional: Wählen Sie mit den Rohdaten, die über den DiSPIM-Monitor in CytoSHOW geöffnet sind, das Polygon-Auswahlwerkzeug (Abbildung 3A, schwarze Pfeile) und klicken Sie direkt außerhalb der Vorder-, Hinter-, Rücken-und Ventralkanten des Embryos (in dieser exakten Reihenfolge) auf Erzeugen Sie ein "Bowtie"-Muster über den Embryo. Tun Sie für beide Ansichten (SPIM-A und SPIM-B, Abbildung 3A).
NOTE: Diese Auswahl gibt die elliptische Region von Interesse (ROI) an, in der der Embryo zentriert ist und zeichnet die anteriorder-nachträgliche Achse des Embryos auf. Das Bowtie-Muster würgt CytoSHOW, dass der Anwender plant, eine präzise Rotation der endgültigen isotropically geschmolzenen Volumina weiter zu spezifizieren, und zwar in eine Orientierung, die für Linienanalysen von StarryNite/AceTree optimal ist. In Fällen, in denen StarryNite Linie nicht Teil des Experimentierplans ist, können andere Auswahlwerkzeuge und-formen ausgewählt werden, um den ROI für die Bildverarbeitung einzustellen. - Wurden mehrere Embryonen gleichzeitig mit Hilfe der Multipositionserfassungsoption abgebildet, öffnen und führen Sie Schritt 9.1 für alle Embryonen aus. Dies wird die parallele Durchführung künftiger Schritte für alle Embryonen in einer Sitzung ermöglichen. Schließen Sie die Fenster SIMA und SPIMB für alle Embryonen, die Sie nicht verarbeiten möchten.
- Klicken Sie auf die DiSPIM-Taste auf der linken Seite des DiSPIM-Monitorfensters (Abbildung 3A, gelb markiert). Dies zeigt eine Untergruppe von Steuerungen, die speziell für die DiSPIM-Verarbeitung bestimmt sind.
- Richten Sie grüne und rote Kanäle für jeden SPIM-Arm aus. Da Emissionskanalsplitter verwendet werden, um bestimmte rote und grüne Bilder gleichzeitig auf der gleichen Kamera zu erfassen, ist es wichtig, die exakte Pixelregistrierung dieser beiden physisch angrenzenden Bildfelder visuell auszurichten, wenn sie überlagert werden. Die Wiederverwendung der gleichen Ausrichtungsanpassungen ist in der Regel über viele aufeinanderfolgende Bildverarbeitungssitzungen möglich, sollte aber überprüft werden (wie in Schritten 9.4.1 – 9.4.5).
- Beginnend mit dem SPIMA-Panel, wählen Sie den roten Kanal aus, indem Sie den CM-Scrollbalken nach links verschieben (Abbildung 3A, orangefarbener Pfeil, linkes Panel).
- Mit Hilfe der X-, Y-und z-Einstellgeräte (Abbildung 3A, orangefarbenes Quadrat) verschieben Sie den roten Kanal auf das Grün.
- Klicken Sie auf DiSPIM-Taste (Abbildung 3A, in gelb hervorgehoben), um das Unterfeld zu schließen und die Ausbreitung der gleichen Verschiebungen auf alle anderen Positionsfenster auszulösen.
- Bestätigen Sie, dass die richtige Ausrichtung, die auf andere Rahmen und Zeitpunkte propagiert wird, durch das Verschieben des "z"-Scrollbalkens (Abbildung 3A, blauer Pfeil, linkes Panel) und/oder "t"-Scrollleiste (Abbildung 3A, grüner Pfeil, linkes Panel). Wenn eine mehrstufige Akquisition durchgeführt wurde und mehrere Embryonen abgebildet wurden (Schritt 5,3), hätte sich die Ausrichtung auch auf diese Embryonen ausbreiten müssen. Bestätigen Sie auch die Zahlen für X-, Y-und z-Einstellgeräte (Abbildung 3A, orangefarbenes Quadrat, das für das SPIMA-Panel aller Embryonen gleich sein sollte).
- Wiederholen Sie die Schritte 9.4.1 – 9.4.4 für das SPIMB diSPIM-Monitorfenster (Abbildung 3A, rechte Tafel).
- Klicken Sie auf den "diSPIM"-Button und dann auf den "Fuse"-Button (Abbildung 3A, blauesRechteck), um ein Dialogfeld mit dem Namen "Deconvolve/FusediSPIM RAw Data Volumes" (Abbildung 3B) zu öffnen. Setzen Sie die Parameter, wie in Abbildung 3Bgezeigt. Diese Parameter werden in den folgenden Unterschritten kurz behandelt:
- Set Key-Registrierung auf Kanal 1 (488 nm Laser) oder 2 (561 nm Laser). Wählen Sie den Kanal mit dichterem oder allgegenwärtigerem Signal. Für Linienversuche wählen Sie immer den Kanal aus, der verwendet wird, um die allgegenwärtige nukleare Histonfluoreszenz abzubilden.
- Setzen Sie die Eingangsvolumina relative Orientierung auf + 1 oder-1. Der richtige Orientierungsindex hängt von den Kamerapazements eines bestimmten DiSPIM ab (Abbildung 4).
Hinweis: Wenn unsicher ist, testen Sie jede Option, indem Sie einen einzelnen Zeitpunkt aus dem Monitorfenster von SPIM A und B diSPIM duplizieren, indem Sie die Schritte 9.1 – 9.12 folgen und willkürlich eine relative Ausrichtung des Eingangsvolumens auswählen. Falsche Orientierungen ergeben unscharfe Bilder mit Artefakten, während korrekte Orientierungen klare Bilder ergeben. Der relative Orientierungswert der Eingangsvolumina , der das klare Bild ergibt, kann dann für alle zukünftigen Daten des gegebenen DiSPIM-Instruments wiederverwendet werden. - Wählen Sie, ob das verschmolzene Volumen genauso ausgerichtet sein soll wie das Eingangsvolumen A oder B (basierend auf der Nutzpräferenz).
- Wählen Sie "Neue Registrierung für jedes Band". Diese Option regelt, wie SpimFusion die Registrierungsmatrizen für jedes Volumenpaar zu jedem Zeitpunkt berechnet. Mit der Option "Fresh" kann der Algorithmus die Registrierung an jedem Zeitpunkt adaptiv optimieren.
- Setzen Sie die Anzahl der Dekonvolution-Iterationen auf 10. Diese Zahl führt tendenziell zuverlässig die gewünschte hohe Auflösung zeiteffizient aus.
- Optional: Wenn die automatische Linienführung gewünscht ist (sehr empfehlenswert), überprüfen Sie die Auto-Lande-Start StarryNite Linie von Fused Volumes. Mit dieser Option wird StarryNite automatisch auf den Markt gebracht, um Zellen in den von SpimFusion produzierten Bildvolumen zu segmentieren und zu verfolgen.
- Optional: Für höchste Genauigkeit in der automatisierten Linienführung ist es am besten, die isotropical verschmolzenen Embryonenbände in die kanonische "ADL" (Anterior [x-west], D orsal [y-north], Left[z-near]) zu positionieren. Wählen Sie die Ausgabeorientierung in der Vorschau-Option, um diese Wahl anzuzeigen. CytoSHOW reagiert mit einer ersten Bearbeitung eines ersten Paares von isotropical verschmolzenen Volumina, so dass der Benutzer die Rotationen, die für die ADL-Registrierung notwendig sind, genau beobachten und angeben kann.
- Klicken Sie auf Ja, sobald alle Parameter ausgewählt sind.
- Geben Sie das Ausgabeverzeichnis an, in dem die bearbeiteten Dateien gespeichert werden können. Klicken Sie auf OK.
- Optional: Wenn die Lautstärke-Ausgabeorientierung in der Vorschau-Option ausgewählt wurde , stellen Sie den t Scroll-Leisten (Abbildung3A, grüner Pfeil, linkes Panel) im SPIM-A-Fenster auf den frühen Timepoint, an dem ABa und ABp-Zellen die Metaphase erreicht haben. Stellen Sie die t Scroll-Leiste im SPIM-B-Fenster auf die spätere Komma-Phase der Entwicklung. Dies wird bei der Spezifikation der ADL-Orientierung helfen.
- Optional: Klicken Sie auf OK, wenn Sie bereit sind. Wenn die Vorschau-Option in 9.5.7 oben ausgewählt wurde, werden nur zwei Vorschauvolumina für die Zeitspunkte, die durch die t-Schieber der Bildfenster SPIM-A und SPIM-B angezeigt werden, isotropical verschmolzen. Diese beiden Vorschauzeigenzeigenpunkte können verwendet werden, um eine präzise Neuausrichtung der Output-Embryonenvolumen auf die ADL-Ausrichtung festzulegen, wie unten erläutert.
- Suchen Sie die neu angezeigte 3DProjY_Decon-Fuse _ ... fenster. Verschieben Sie den t Scroll-Balken auf den Punkt 2 dieses Vorschaufensters. Bewegen Sie den Z-Schieberegler, bis der Blick direkt die lange Achse des Embryos zeigt.
- Verschieben Sie den t Scroll-Balken zurück auf den Zeitpunkt 1 des 3DProjY_Decon-Fuse _ ... fenster. Wählen Sie das Linienauswahlwerkzeug aus und zeichnen Sie eine Linienauswahl aus der EMS-Zelle (der rundenste Kern) durch die Ebene der B-Zell-Metapphaben-Platten.
- Klicken Sie auf die orangefarbene DiSPIM-Vorschau-Taste auf der 3DProjY_Decon-Fuse _ ... fenster. Die Feineinstellungen an die Ausrichtung des vorhergesehenen Bildvolumens werden für die Bearbeitung des vollständigen Datensatzes gespeichert.
- Optional: Der Dialog Deconvolve/Fuse diSPIM Rohdatenvolumen wird wieder angezeigt, genau wie in Schritt 9.5 oben. Klicken Sie auf Ja, ohne in der Vorschau-Option die Ausgabeorientierung definieren . Geben Sie den Ausgabedorder für den kompletten Datenverarbeitungsablauf an.
- Stellen Sie die t Scroll-Balken (Abbildung 3A, grüner Pfeil, linkes Panel) des DiSPIM-Monitorfensters auf den Startpunkt (SPIMA) und den Endpunkt (SPIMB) der gesamten Bildspannweite, die zu verarbeiten ist. Dann klicken Sie auf Ok.
- Im Laufe der SpimFusion öffnet und erfrischt CytoSHOW ein mehrdimensionales Fenster, das das geschnittene 4D-isotropische, verschmolzene Volumen für jeden Embryo zeigt, sowie zwei Fenster mit drehenden 4D-Mathematikintensitätsprojektionen des isotropen Volumens. Während dieser Zeit sollten Sie kein CytoSHOW-Fenster stören oder schließen, bis die isotrope Fusion und die Linienverfolgung abgeschlossen sind.
- Optional: Beachten Sie, dass, sobald der StarryNite-Splash-Screen erschienen ist und später verschwunden ist, die vollständige Datenverarbeitung abgeschlossen ist. Dieses Fenster darf während der Bearbeitung nicht geschlossen werden oder StarryNite wird unterbrochen.
10. Eröffnung der StarryNite Linie Trace Serie in AceTree (optional)
- Öffnen Sie die angepasste Version von "AceTree_16BitCompat.jar" zurVerfügung gestellt.
- Wählen Sie das Menü " Datei > Öffnen der Konfigurationsdatei". Suchen Sie Ihr Ausgabeverzeichnis , das zuvor auf CytoSHOW angegeben wurde. Öffnen Sie den Decon _ Fuse _ ... _ Pos [n ] Unterordner für Embryo [n]. Wählen Sie aaa _ edited.xml und Open.
- Verwenden Sie das Menü " Bearbeiten > Bearbeiten Werkzeuge bearbeiten, um die Fenster " Track bearbeiten "zu öffnen und die Fenster zu löschen oder zu löschen.
- Klicken Sie auf den halb schattigen Kreis Abbildung 5B, rotes Quadrat, um die Rot-Grün Intensitäten anzupassen.
- Gehen Sie mit der Abstammungsvisualisierung und-bearbeitung voran, wie bereits beschrieben 5,6,8 (Manuskripte sind auch in unserem Download-Bundle enthalten).
Representative Results
Wir haben zunächst die Lebensfähigkeit von Embryonen bestätigt, die mit den Parametern des Protokolls für die DiSPIM-Übernahme (Abschnitte 1-6) abgebildet wurden. Zehn Embryonen wurden gleichzeitig bei 20 ° C, einer Volumen-/Embryo/Minute, vom 2-Zellen-Stadium bis zum 2-fachen Stadium (7,5 Stunden, 451 Volumina/Embryo) abgebildet. Um die Zellteilung während der Embryogenese zu überwachen, haben wir die Sorte BV514 verwendet, die allgegenwärtig die mCherry::Histon-Reporter aus dem integrierten Transgenarray-Array ujIs113 24 ausdrückt. Abbildung 6 zeigt eine Zeitachse dieser ersten Hälfte der Embryonalentwicklung für einen der abgebildet Embryonen. Jedes Bild stellt eine Projektion mit einer maximalen Intensität (erzeugt durch die Schritte 7-8) des abgebildeten Embryos dar. Wir fanden heraus, dass die optimierten Protokolle keine nachweisbare Phototoxizität zu den Embryonen veranlassten, wie der Zeitpunkt der Zellteilung (nicht abgebildet), die Zeit des Schlüpfens und das Timing, das mit den Meilensteinen der Entwicklung zusammenhängt, bewertet wurden (Abbildung 6 und Referenzen 1). , 25 , 26).
Wir haben dann das Protokoll angewendet, um die Auswuchsdynamik einzelner Neuronen bei der Entwicklung von Embryonen zu analysieren. Wir haben DCR7692 (olaex4655)abgebildet, Eine transgene Nematode-Strafung, die GFP aus dem Neuroptid-Flp-19-Promoter in einer Untergruppe von nicht identifizierten Zellen (DACR2819, Pflp-19 ( 3.6kb) ausdrückt :: Syn21:CAAX::p 10 3 ' UTR) . Nach den Schritten des hier skizzierten Protokolls stellten wir fest, dass die nicht identifizierten Zellen den motorischen Neuronen RMDDL und RMDDR, der Ausscheidungskanalzelle und zwei Muskelzellen entsprechen (Abbildung7). Wir haben dann die Auswuchsdynamik der RMDDL und RMDDR-Neuronen untersucht und quantifiziert. Wir beobachteten, dass die RMDDL und RMDDR-Neuronen bereits 360 Minuten nach der Befruchtung schräg geformt sind, wobei die längere zelluläre Achse die nachfolgende Achse für Neurit-Auswuchs darstellt (Abbildung 7 und Film S1). Mit dem "einfachen Neurit-Tracing"-Plugin in FIJI und der Anwendung auf 3D-Rekonstruktionen isotropical verschmolzenen Volumina quantifizierten wir dann das stereotype Auswachsen der RMDDL und RMDDR-Neuriten für sechs Embryonen. Wir stellten fest, dass die Outwusting-Dynamik für RMDDL und RMDDR über Embryonen hinweg (hier RMDDs genannt) stereotypisiert wurde. Ab 385-410 Minuten Nachdüngung verlängerten die RMDDs Neuriten 6,0 ± 0,5 μm (Mittelwert ± SEM; n = 12 Neuriten) vor den Zellkörpern (Abbildung7B, C, I). Von 415-445 Minuten Nachdüngung dehnen sich beide Neuriten dorsal in und um den voraussichtlichen Nervenring aus (Sternchen in Abbildung 7D). Im Durchschnitt hat sich jeder RMDD-Neurit um 11,0 ± 0,6 μm verlängert (Mittelwert ± SEM; n = 12 Neuriten) aus dem Zellkörper, bevor er synchron auf sein kontralaterales Pendant an der Spitze des Ringes trifft (Abbildung 7I). Wichtig ist, dass unsere repräsentativen Ergebnisse zeigen, dass wir in der Lage sind, neuronale Entwicklungsmerkmale für einzelne identifizierbare Zellen anhand unseres integrierten Protokolls zu untersuchen, zu vergleichen und zu quantifizieren (Abbildung 7 und Abbildung 8).
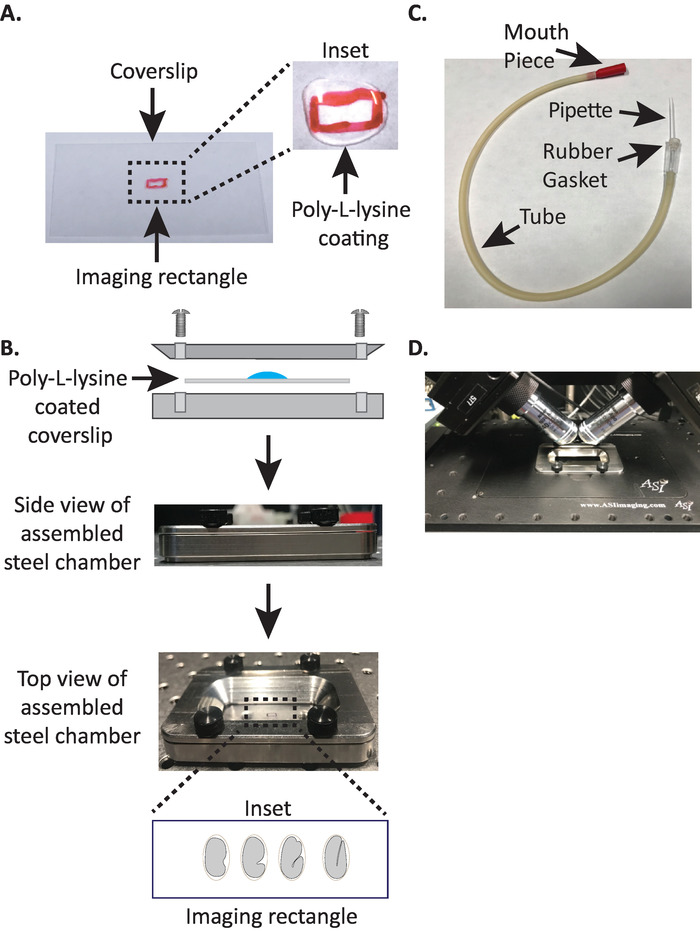
Abbildung 1: DiSPIM-Probenmontage. (A) Zubereitung der Kerkel mit Poly-L-Lysin. Im Einsetzen wurde 10 μL Poly-L Lysin verwendet, um den Keillip für 5 Minuten zu beschichten. Poly-L-Lysin ermöglicht es der Embryonalechale, fest an der Keillinie im Rechteck zu haften. B) Schematic der Stahlbildkammer und der zusammengesetzten Kammer. Im Einsetzen orientieren sich repräsentative Embryonen mit der anterioralen Nachachse senkrecht zur langen Achse am Keillip. (C) Assembled Aspirator-Röhre mit mikrocapillaren Pipette. D) Stahlbildkammer, die im Probenhalter nach den Zielen von diSPIM 40x montiert ist. Bitte klicken Sie hier, um eine größere Version dieser Figur zu sehen.

Abbildung 2: Langfristige DiSPIM-Bildgebungseinrichtung im Micro-Manager. A) empfohlene DiSPIM-Laserleistungsparameter (rotes Rechteck), die für eine längere Bildgebung bei gleichzeitiger Reduzierung der Phototoxizität optimiert sind (gemessen an einer höheren Schraffurate von C. eleganischen Embryonen). Set 561 nm Laser auf 79 μW (0,25) und 488 nm Laser auf 179 μW (0,5). Beachten Sie, dass die exakte Kalibrierung von Software-Einstellungen auf Laserleistung zwischen den DiSPIM-Installationen variiert. Es wird empfohlen, dass Anwender die Laserleistung messen und kalibrieren, um 79 μW (561 nm) und 179 μW (488 nm) Laserleistung zu erreichen. (B) DiSPIM-Parameter zur Datenspeicherung (grünes Rechteck), Lautstärke-Einstellungen (blaues Quadrat) und Slice-Einstellungen (orange Quadrat). (C) diSPIM-Autofokus-Parameter für die langfristige Abbildung von C. elegans Embryogenese (siehe Schritte 6.1-6.6). Bitte klicken Sie hier, um eine größere Version dieser Figur zu sehen.

Abbildung 3: Bildvisualisierung und Datenverarbeitung mit CytoSHOW. (A) Raw diSPIM Bilder, die von CytoSHOW geöffnet wurden. CytoSHOW ist in der Lage, Bilder zu öffnen, die von beiden Kamerawegen (SPIM A und B) aufgenommen wurden. Diese Rohbilder werden in mehrdimensionalen Fenstern geöffnet, die diSPIM-Monitorgenannt werden. Im DiSPIM-Monitorwird ein "Bowtie-Muster" erzeugt, um die vorderen, hinteren, dorsalen und ventralen Kanten des Embryos auszuwählen (siehe Schritt 9.1). Bow-tie Selektionen zeigen die Embryonenorientierung für die Dekonvolution und die StarryNite-assistierte Linienverfolgung an. (B) Optimierte Parameter zur Erzeugung von isotropen Bildern. Setzen Sie im Dekonvolved beim Erwerb des Fensters die in Schritten 9.5.1-9.5.8 angegebenen Parameter. Bitte klicken Sie hier, um eine größere Version dieser Figur zu sehen.

Bild 4: DiSPIM-Kamerakonfiguration. (A) Foto von DiSPIM-Kamera-Platzierungen und Orientierungen. B) Darstellung von + 90-Grad-Rotationen von SPIM A, um SPIM B-Bilder zu entsprechen. (C) Eingabebumina im Verhältnis zum Orientierungsindex + 1 auf Basis der Kamerakonfiguration unseres DiSPIM (siehe Schritt 9.5.2). Wir drehen SPIM A Bild (s) + 90 Grad um Y-Achse vor der Registrierung, um SPIM B Bilder (s) zu entsprechen. Skalierbalken = 10 μm. Bilder sind repräsentative Einzelansicht, Maximalintensitätsprojektionen und Dekonvolution-Bilder des 1,5-fachen Embryos mit beschrifteten Kernen (561-nm, rot) und Neuronen (488-nm, grün). Bitte klicken Sie hier, um eine größere Version dieser Figur zu sehen.

Abbildung 5: Kuration und Bearbeitung C . Eleganten embryonale Linie in AceTree. (A) Wir verwenden AceTree, um die Abstammungsspuren von StarryNite zu bearbeiten (siehe Referenzen 5,6,8; Manuskripte sind auch in unserem Download-Bundle enthalten). AceTree zeigt C . elegans systematische Namen für jeden Kern (grünes Rechteck) nach Abschluss der Schritte 10.1-10.2 an. Dieses Fenster (A) bietet Informationen (schwarzes Rechteck) über jede Zelle in der Linie Spur (ABa, in blau hervorgehoben), die den Nutzern helfen, die Linienspuren zu verfolgen und zu bearbeiten. Es wird empfohlen, dass Nutzer lineare Zellen und ihre Positionen mit der C. elegans embryonalen Zelllinie, die zuvor von Sulston et al. Dekonalisierte Datenreihen (siehe unten, B) geben den Systemnamen C. elegans in der Suchleiste ein (orangefarbenes Rechteck). (B) Die dekonvertierte Datenreihe des Nutzers öffnet sich auch automatisch nach Abschluss der Schritte 10.1-10.2. Gezeigt wird hier ein isotropical verschmolztes Bild eines Vier-Zellen-Stadionembryos mit in Rot beschrifteten Kernen. Während der Verfolgung eines Kerls sollten die Benutzer die Intensität des Bildes ändern (rotes Quadrat) und mit den Pfeiltasten auf ihrer Tastatur durch die Zeit und z navigieren (time-left/right, z-up/down). (C) 3D-Zeichentrickfilm des Zeitrahmenin (B) mit bestimmten Funktionalitäten (lila Rechteck), die eine drehbare 3D-Visualisierung ermöglicht. Eine Übersicht über AceTree und seine 3D-Funktionalität finden Sieunter Referenzen 5,6,8. Bitte klicken Sie hier, um eine größere Version dieser Figur zu sehen.

Abbildung 6: Zeitgemäße Entwicklungsdynamik von C. eleganischem Embryonen auf der DiSPIM. Top-Panel, DiSPIM-Bilder, die die erste Hälfte der Embryonalentwicklung für einen der abgebildeten Embryonen zeigen (Strain BV514 ujIs113 24). Embryonen wurden kontinuierlich abgebildet, jede Minute für 7,5 Stunden (bei 20 ° C). Die ersten beiden Bilder der oberen Tafel repräsentieren 4-und 8-Zellen-Embryonen mit Kernen (rot) und Positionen polarer Körper (dichte rote Kugeln, markiert mit blauen Sternchen). Jedes Bild stellt eine Einbliw-Maximalintensitätsprojektion des abgebildeten Embryos dar. Skalierbalken = 10 μm. Die Zeitachse (horizontaler Balken) stellt eine minder postbefruchtete (m.p.f.) der Entwicklung von C. elegans Embryonen dar. Wir bestätigten, dass die Parameter unseres Protokolls für die DiSPIM-Übernahme keine nachweisbare Phototoxizität zu den abgebildeten Embryonen veranlasst haben, wie sie durch die Lebensfähigkeit, den Zeitpunkt der Zellteilung, den Zeitpunkt des Schraffings und den Zeitpunkt der Entwicklung von Meilensteinen bewertet wurden (siehe Referenzen). 1 , 25 , Wirstellen fest, dass der Zeitpunkt der Meilensteine der Entwicklung über Embryonen hinweg mit unseren Bildungsparametern reproduzierbar war (SEM ± 8,174 Minuten für 6,4 Stunden lange bildgebende Sitzungen; N = 10). Bitte klicken Sie hier, um eine größere Version dieser Figur zu sehen.

Bild 7. Zelluläre Identifikation und Einzelzellcharakterisierung der Neurit-Auswuchdynamik bei der Entwicklung von C. eleganen Embryonen. Duale Farbdarstellung einer Sorte, die durch die Überquerung von BV514 ujIs113 24 (für Lineaging) und DCR7692 (olaex4655), einer transgenen Nematoden-Stamm, die GFP von der Neuroptid-Flp-19-Promoter in einer Untergruppe von nicht identifizierten Zellen ausdrückt. (A-H) Nach den Schritten des hier skizzierten Protokolls stellten wir fest, dass die nicht identifizierten Zellen den motorischen Neuronen RMDDL und RMDDR (gelbe Pfeile), der Ausscheidungskanalzelle (blaue Pfeile) und zwei Muskelzellen (weiße Pfeile) entsprechen. I) Quantifizierung der Auswuchdynamik der RMDDL und RMDDR-Neuronen durch die Verwendung von FIJI-Plugin "einfache Neuritverfolgung" und die Anwendung auf 3D-Rekonstruktionen isotropical verschmolzenen Volumina. Beachten Sie, dass sowohl RMDDL als auch RMDDR eine stereotype Auswuchsdynamik aufweisen, die sich synchron auf eine Gesamtlänge von 11,0 ± 0,6 μm ausdehnt (Mittelwert ± SEM; n = 12 Neuriten) und Treffen an der dorsalen Spitze des Nervenrings (siehe auch Film S1). Bitte klicken Sie hier, um eine größere Version dieser Figur zu sehen.

Abbildung 8: Untersuchung von Isotropen DiSPIM-Bildern neuronaler Morphologien in C. eleganischem Embryonen. Isotropische Visualisierung von AVHL und AVHR Neuronen (gelbe Pfeile). Mit Hilfe des DiSPIM lassen sich neuronale Morphologien erfassen, die vierdimensionale (4D) Bilder mit einer isotropen Raumauflösung von etwa 330 nm ergeben. Das diSPIM ermöglicht es dem Benutzer, Bildvolumen mit identischer Auflösung in alle Richtungen virtuell zu drehen. Bilder in A-D sind Projektionen mit maximaler Intensität des gleichen isotropical geschmolzenen DiSPIM-Bildvolumens aus unterschiedlichen Rotationen um die lange Achse des Embryos. Skalierbalken = 5 μm.
Ergänzende Film S1 : C. elegans Embryo entwickelt sich von 280 bis 434 Minuten nach der Befruchtung. Isotropen Film der Sorte DCR7692 (olaex4655), die ujIs113 allgegenwärtig mit DACR2819 spärlich beschrifteten RMDD-Neuriten (Abbildung 7A-D, gelbe Pfeile) ausdrückt. DACR2819 kennzeichnet auch zwei Muskelzellen(Abbildung 7A-D, weiße Pfeile) und Ausscheidungskanalzellen (Abbildung7A-D, blauer Pfeil) während der embryonalen Entwicklung (Abbildung 7A-D). Skalierbalken = 10 μm.
Discussion
C. elegans sticht als der einzige Organismus mit den letzten Positionen und der Konnektivität jedes erwachsenenNeuronsbekannt 27. Die Entwicklungsdynamik, die zur Organisation der Arbeitskreise und Netzwerke führt, aus denen sich die C. elegans connectome zusammensetzt, ist jedoch unbekannt. Basierend auf den Chancen, die sich aus den Fortschritten in der Lichtmikroskopie ergeben, können wir nun Zellpositionen, Morphogenese und Neurogenese in der gesamten Embryonalentwicklung von C. elegans erfassen und analysieren.
Die Prozedur, die wir beschrieben haben und die wir routinemäßig im Labor verwenden, liefert 4D-isotrope Bilder von beschrifteten Neuronen und Kernen für die Zelllineare in C. eleganischem Embryonen. Noch wichtiger ist, dass wir mit den DiSPIM und den gekoppelten semi-automatischen Linienbildungsfunktionen langfristige Bildgebungsbedingungen optimiert haben, um die Geschwindigkeit und Präzision der Analyse von C. elegans Embryogenese zu verbessern. Dieses integrierte Protokoll wird es den Nutzern ermöglichen, Zellen zu visualisieren und zu identifizieren und dreidimensionale Merkmale wie Neuriten-Migration und Axonfaschisierung durch den Beginn des frühen Twitching zu quantitieren. Dieses Verfahren lässt sich mit einem ASI-DiSPIM-System problemlos in jede Anlage integrieren, und wir empfehlen dieses System speziell für dieses Protokoll. Andere SPIM-Formulierungen, die kommerziell angeboten werden, können sich von der ASI-Konfiguration in der Probenkammer und den optischen Eigenschaften unterscheiden. Daten, die von anderen Plattformen exportiert werden, können aber auch über unsere Datenpipeline übertragen werden. Daher ist eine Bewertung ihres Wertes in der Linienführung, einem anspruchsvollen Test der Bildqualität und der Stabilität des Instruments, machbar. Auch wenn wir die DiSPIM aktiv nutzen, um regelmäßig andere Exemplare (wie Drosophila und Zebrafisch-Embryonen) abzubilden, ist die beschriebene und umfassende lineätische Analyse von Embryonen derzeit noch auf die Nematoden-Arten beschränkt. Bei größeren oder dicken Proben entscheiden wir uns für die Verwendung von Bühnenscannern, die die Proben durch ein stationäres Lichtblech scannen. Kumar et al. haben zuvor diese verbesserte DiSPIM-Sparung demonstriert, um qualitativ hochwertige Bilder aus dicken Proben ohne zusätzliche Änderungen an der DiSPIM10zu erzeugen.
Zu den kritischen Schritten innerhalb des Protokolls gehören die Montage von C. elegans Embryonen an der poly-Lysin-lysin-beschichteten Coverslip, Datenerfassung und Datenverarbeitung. Die Ernte und Montage von C. eleganischem Embryonen an den Glaskappe kann für unerfahrene Anwender eine Herausforderung sein, aber hier stellen wir ein detailliertes Protokoll der wichtigsten Schritte zur Verfügung, um das Lernen zu erleichtern. Wenn eine langfristige Bildgebung gewünscht ist, erhalten wir die besten Ergebnisse bei der Ernte von Vierzellen oder früheren Embryonenvon 8-10 jungen Erwachsenen 28. Es ist zu beachten, dass ältere Erwachsene weniger wünschenswert sind, Embryonen im Frühstadium zu ernten, da sie dazu neigen, ältere Embryonen in der Gebärmutter und unbefruchtete Eizellen zu enthalten. Bei der Montage von Embryonen können Probleme wie die Blockade des montierten Aspirators (Mundpipette) oder eine zu große Öffnung in der Mikrokapillarpipette eine korrekte Montage und Orientierung der Embryonen verhindern. Um sich auf eine optimale Bildgebung vorzubereiten, führen wir Vorbeschaffungstests an frühen und späten Pre-twittifenden Embryonen durch, um die Leistung der Lichtblätter, Kameras, Ziele und Autofokus zu überprüfen. Wir erzielen beste Ergebnisse, wenn alle diese Operationen getestet werden und liefern qualitativ hochwertige Bilder während unserer Vorbeschaffungstests. Dies ist vor allem für die Erzeugung von Bildern mit isotropher Raumauflösung von Bedeutung, für die Rohbilder, die aus beiden Blickwinkeln (Ziele) aufgenommen wurden, von hoher Qualität sein müssen. Nach der Akquisition werden die für jede Ansicht erworbenen Volumina zu isotropen Bildern verarbeitet. Es ist wichtig, eine entsprechende Grafikverarbeitungseinheit (GPU) zu verwenden, wie in diesem Protokoll beschrieben (siehe unten). Dadurch wird die Verarbeitungsgeschwindigkeit verbessert, mit der die isotropical verschmolzenen Bilder erzeugt werden, was die Zeit für Datenanalysen verkürzt. Es ist auch zwingend erforderlich, dass die Benutzer die neueste Version von CytoSHOW verwenden und die mit unserem Download-Bundle für StarryNite Auto-Linaging zur Verfügung gestellten Parameter verwenden. Wenn Nutzer daran interessiert sind, die automatische Linie für andere Proben (z.B. Zebrafische, Drosophila etc.) zu verwenden, ist eine zusätzliche Optimierung der in StarryNite verwendeten Parameter erforderlich (siehe Referenzen3,4).
Obwohl unser integriertes Protokoll Bilder und Linienresultate im Pre-twitching-Embryo liefert, sollten sich die Nutzer darüber im Klaren sein, dass eine automatisierte Linienführung im Post-twittiching-Embryo derzeit nicht machbar ist: Die nuklearen Positionen ändern sich in der Reihenfolge der Sekunden in der Nachzwitschern Embryo, zu schnell, um Abstammung zu ermöglichen. In der Tat hat die DiSPIM in der Tateinevielversprechende Fähigkeit gezeigt, neuroentwicklungsbedingte Ereignisse zu erfassen und einige Zellpositionen in den nachtwitternden Stadien der Embryogenese 23,29zu verfolgen. Wenn Nutzer daran interessiert sind, den nachtwitternden Embryo zu untersuchen, bietet das diSPIM die Geschwindigkeit an, um volumetrische Schnappschüsse zu erhalten und feine neuroentwicklungsbedingte Ereignisse, wie Neuriten-Auswuchs, in schnell bewegten Embryonen zu verfolgen.
Dieses Protokoll wird für die Zell-by-Zell-Fertigstellung des WormGUIDES Atlas 30 grundlegend sein,da es einen integrierten Ansatz mit hochauflösenden isotropen Bildern bieten wird, um 3D-Morphologien von beschrifteten Neuronen zu identifizieren und zu erfassen. Die ersten 430 Minuten der Embryogenese. Der Prototyp WormGUIDES Atlas liefert derzeit nukleare Positionen von Zellen im sich entwickelnden Embryo und zielt darauf ab, die Entwicklungsdynamik einer Teilmenge von embryonalen Neuronen zu erfassen. Dieses Protokoll wird ein Schlüssel für die Integration von zusätzlichen sich entwickelnden Neuronen in den WormGUIDES Atlas30 sein.
Unser integriertes Protokoll wird auch die Erforschung neuer Genexpressionsprofile im C. eleganischen Embryo vereinfachen. In transgenen C. elegans steuernviele zellspezifische Promoter räumlich und zeitlich die Transgenexpression. Während die Expressionsmuster der meisten Gene beim erwachsenenTier 31,32, 33,34ausgiebig charakterisiert wurden, müssen in der Entwicklung fast alle noch charakterisiert werden (insbesondere Spätstadium) Embryo. Das Promoterom C. elegans war eine nützliche Ressource für die Wurmgemeinschaft, um die zellspezifische Transgene-Expression zu fördern und festzustellen, ob die G-Funktion zellautonom oder nicht autonom ist. Die Erfassung isotropes hochauflösender und dynamischer Expressionsmuster von Genen und die präzise Identifizierung von Expressionsgegenständen über Linaging wird für viele in der wissenschaftlichen Gemeinschaft wertvoll sein.
Die Embryogenese besteht aus zwei miteinander verknüpften Hauptprozessen, der zellulären Differenzierung und der Gewebemorphogenese. Über die Mechanismen und Moleküle, die bei der Entwicklung von C. elegansverschiedene Zelltypen definieren, ist viel bekannt. Über die Mechanismen, die für die Zellmigration, die Zellhaftung und die Zellform im C. eleganischem Embryo wichtig sind, ist jedoch wenig bekannt. Mit der invarianten Zelllinie C . elegans können wir mit unserem Protokoll die katalogisierte 3D-Mikroanatomie des Embryos während der Morphogenese auf neuen Detailebenen erkennen: z.B. Axonfaschisierung, Synaptogenese und neuronale Aktivität. Ardiel et al. haben zuvor die Macht des DiSPIM demonstriert, Kalziumvergängiger auf der Ebene eines einzelnen Neurons in C. elegans Embryonen23 zu erfassen. Viele andere Aspekte der Entwicklungsphysiologie sind mit diesen Methoden reif für die Untersuchung.
Schließlich ist dieses Protokoll weitgehend automatisiert und reduziert systematisch die Zeit, die es braucht, um Dekonvolution-Bilder zu erzeugen und Zell-Linienführung über StarryNite und Acetree durchzuführen. Die in diesem Protokoll verwendeten Softwarestrategien können auf viele Fragen der Biologie angewendet werden, die weit von den ganz spezifischen Bereichen entfernt sind, in denen wir sie hier demonstriert haben.
Details zur Software-Kompatibilität und zum Download-Zugang
Informationen zu Micro-Manager und Plugins für die DiSPIM-Bildgebung gibt es bei http://dispim.org/software/micro-manager und https://micro-manager.org/wiki/ASIdiSPIM_Plugin.
Die Datenverarbeitungs-Pipeline benötigt derzeit ein Windows-Betriebssystem. Wir haben eine einzelne Archivdatei gebündelt, um die Installation aller erforderlichen Datenverarbeitungsprogramme und Support-Dateien zu vereinfachen. Sie steht zum Download auf http://dispimlineage.wormguides.org zur Verfügung.
CytoSHOW (http://run.cytoshow.org/) basiert auf der weit verbreiteten und Open-Source-Bildanalyseplattform ImageJ (v1). Java muss auf dem Computer installiert und auf dem neuesten Stand sein, um CytoSHOW zu verwenden, und Updates zu CytoSHOW werden automatisch über Java Web Start eingesetzt. Viele ImageJ-basierte Funktionen von CytoSHOW sind wie beschrieben und auf https://imagej.nih.gov/ij/docs/examples/index.html illustriert. CytoSHOW wurde angepasst, um mehrdimensionale Rohdaten des ASI diSPIM sowie anderer Bildbearbeitungssoftware anzuzeigen, die TIFF-Ausgabe erzeugt. Im Prinzip könnten andere SPIM-Bildgebungssysteme mit mehreren Ansichten durch kleinere Modifikationen von CytoSHOW unterstützt werden, um dieses Protokoll auf verschiedenen Mikroskopsystemen durchführen zu können.
SpimFusion wurde in CUDA/C + + mit Visual Studio 2013 mit CUDA Toolkit v7.5 geschrieben. Das Laufen von SpimFusion erfordert eine spezielle Computer-Hardware: Eine NVIDIA-Grafikverarbeitungseinheit (GPU) mit CUDA-Komputationsfähigkeit 1,0 oder höher und mindestens 2 GB Grafikkarten-Speicher. Zum Zeitpunkt der Veröffentlichung unseres Protokolls ist SpimFusion unveröffentlicht (Min Guo und Hari Shroff), aber im oben genannten Software-Bündel-Archiv verfügbar.
Eine speziell gebaute Kommandozeilenversion von StarryNite verlangt, dass der frei verfügbare MATLAB Compiler Runtime installiert ist, aber keine Lizenz für kommerzielle MATLAB-Software benötigt. Der MATLAB Compiler Runtime ist im oben genannten Software-Bundle-Archiv enthalten. Der Code für StarryNite, wie er in diesem Protokoll verwendet wird, ist im Wesentlichen unverändert als der für konfokaleBilder 6. Allerdings wurden hier mehrere operative Fragen bei der Erstellung von Eingangsbildern für die StarryNite-Verarbeitung und dem Umgang mit StarryNit-Ergebnissen mit Methoden in CytoSHOW befasst, die eine kontinuierliche Datenverarbeitungs-Pipeline für verschmolzene isotrope diSPIM ermöglichen. Volumes. Diese Änderungen werden durch den CytoSHOW-Code automatisiert, der diese Vor-und Nachbearbeitungsschritte bearbeitet. CytoSHOW editiert auch eine voroptimierte diSPIM-spezifische Vorlage StarryNite Parameter gesetzt, um automatisch den Segmentierungsalgorithmus auf die Fluoreszenz-Intensität von Kernen in den abgebauten Daten zu stimmen. Die einzigartigen Parameter, die StarryNite auf jedem DiSPIM-Datensatz verwendet, werden dann zusammen mit dem Ausgabebild und den Liniendaten in einer Datei gespeichert.
Eine benutzerdefinierte Version von AceTree, die mit 16-Bit-Bildern arbeitet und die Kompatibilität mit dem Joom3D-Rendering beibehält, ist für dieses Protokoll am besten geeignet. Es ist auch in dem oben genannten Software-Paketarchiv enthalten.
Disclosures
Die Autoren haben nichts zu offenbaren.
Acknowledgments
Wir danken John Murray für die integrierte Sorte, ujIs113, für die Erzeugung von Linienstrakt BV514; Brandon Harvey (NIBIB) für die Hilfe beim Testen des Protokolls; Jon Daniels und Gary Rondeau (Angewandte wissenschaftliche Instrumente) für die Unterstützung von Micro-Manager und DiSPIM-Instrument; Und Andrew York und Hank Eden für ihr kritisches Feedback zum DiSPIM-System. Wir danken auch dem Forschungszentrum für Minderheitseinrichtungen und dem Instituto de Neurobiología Jose del Castillo (Universidad de Puerto Rico) für die Bereitstellung einer Tagungs-und Brainstorming-Plattform. Ein großer Teil dieser Arbeit wurde im Rahmen des Whitman Programms am Marine Biological Laboratory in Woods Hole durchgeführt. Diese Arbeit wurde unterstützt durch die intramuralen Forschungsprogramme des NIH-Nationalen Instituts für Biomedizinische Bildgebung und Bioengineering und durch das NIH-Stipendium Nr. U01-HD075602 und Nr. R24-OD016474. Mark W. Moyle wurde von F32-NS098616 und Leighton H. Duncan unterstützt von einem Diversity Supplement to R24-OD016474.
Materials
| Name | Company | Catalog Number | Comments |
| Steps 1-4 | |||
| Concavity slides | ThermoFisher Scientific | 1519006 | 5-18mm diameter, 0.6-0.8mm deep, 1.2-1.5mm |
| Dissecting microscope with 10×–50× zoom range | Motic | SMZ-171 | |
| E. coli (OP50) | Caenorhabditis Genetics Center (CGC) | OP50 | |
| Glass coverslips, no. 1.5, 24 × 50 mm | VWR International | 48393-241 | |
| M9 Buffer | Stiernagle, T. Maintenance of C. elegans. WormBook. 1-11, doi:10.1895/wormbook.1.101.1, (2006). | ||
| Methyl cellulose | Sigma-Aldrich | H7509-25G | |
| Microcapillary pipette aspirator tube | Sigma-Aldrich | A5177 | |
| Microcapillary pipettes, 0.4-mm i.d | Drummond Scientific | 1-000-800 | |
| Needle, no. 18G x 1 ½ (1.2mm x 40mm) | BD Precision Glide | 305196 | |
| NGM plates | prepared as described by Brenner (1974) | ||
| O-ring for imaging chamber | O-Rings West | M1.5X40 | |
| Pasteur pipette | Corning/Sigma-Aldrich | CLS7095D5X | |
| Platinum wire, 0.5-mm diameter | Sigma-Aldrich | 267201 | |
| Poly-L-lysine | Sigma-Aldrich | P1524 | |
| Stainless steel rectangular chamber (76.0 mm x 50.5 mm) | Applied Scientifics Instrumentations (ASI) | I2450 | |
| Worm Eyelash Pick | Hart, A. C. Behavior. WormBook. (2006). | ||
| Worm Pick | Stiernagle, T. Maintenance of C. elegans. WormBook. 1-11, doi:10.1895/wormbook.1.101.1, (2006). | ||
| Name | Company | Catalog Number | Comments |
| Steps 5-6 | |||
| 488 nm long-pass filter | Semrock | LP02-488 RU-2 | |
| 561-nm notch filter | Semrock | NF03-561E-25 | |
| BLP02-561R-25, quantity 2 | Semrock | 561 nm EdgeBasic best-value long-pass edge filter | |
| Control software for bottom camera | Jenoptik | ProgRes CapturePro | |
| diSIPM assembly video | Applied Scientifics Instrumentations (ASI) | https://youtu.be/TAgbr6IrTqw ; http://www.asiimaging.com | |
| diSPIM alignment video | Applied Scientifics Instrumentations (ASI) | https://youtu.be/qnOrg30NNuE | |
| diSPIM imaging PC | Intel | Intel Xeon CPU E5-2630 2.6GHz, 12 cores in total, 64 GB memory, Windows 7 | |
| FF01-525/45-25, quantity 2 | Semrock | 525/45 nm BrightLine single-band bandpass filter | |
| FF555-DI03-25X36, quantity 2 | Semrock | 555 nm edge BrightLine single-edge dichroic beamsplitter | |
| Imaging PC Graphics Card | NVIDIDA | NVIDIA GeForce GTX 1080 Ti graphics cards | |
| Kumar et al diSPIM Setup | Applied Scientifics Instrumentations (ASI) | Instrument setup for this protocol is identical to Kumar et al 10,11 diSPIM, which makes use of 40x 0.8NA water immersion lenses for imaging. (See steps 5.1 and note) | |
| Micro Manager | Micro-Manager | https://micro-manager.org/ | |
| Modifications to Kumar et al diSPIM Setup (see below) | |||
| Optical table with isolators, 4 feet × 6 feet × 12 inches | TMC | 784-651-02DR and 14-416-34 | |
| Name | Company | Catalog Number | Comments |
| Steps 7-10 | |||
| Analysis PC | Intel | Intel Core i7-8700K CPU 3.70GHz, 6 cores in total, 64 GB memory, Windows 10 | |
| Analysis PC Graphics Card | NVIDIDA | NVIDIA GeForce GTX 1080 Ti graphics cards | |
| Installation instructions | Software bundle | http://dispimlineage.wormguides.org/diSPIMlineaging_InstallationInstructions.htm | |
| Software bundle | Software bundle | http://dispimlineage.wormguides.org |
References
- Sulston, J. E., Schierenberg, E., White, J. G., Thomson, J. N. The embryonic cell lineage of the nematode Caenorhabditis elegans. Developmental Biology. 100 (1), 64-119 (1983).
- Bao, Z., et al. Automated cell lineage tracing in Caenorhabditis elegans. Proceedings of the National Academy of Sciences of the United States of America. 103 (8), 2707-2712 (2006).
- Santella, A., Du, Z., Bao, Z. A semi-local neighborhood-based framework for probabilistic cell lineage tracing. BMC Bioinformatics. 15, 217 (2014).
- Santella, A., Du, Z., Nowotschin, S., Hadjantonakis, A. K., Bao, Z. A hybrid blob-slice model for accurate and efficient detection of fluorescence labeled nuclei in 3D. BMC Bioinformatics. 11, 580 (2010).
- Boyle, T. J., Bao, Z., Murray, J. I., Araya, C. L., Waterston, R. H. AceTree: a tool for visual analysis of Caenorhabditis elegans embryogenesis. BMC Bioinformatics. 7, 275 (2006).
- Katzman, B., Tang, D., Santella, A., Bao, Z. AceTree: a major update and case study in the long term maintenance of open-source scientific software. BMC Bioinformatics. 19 (1), 121 (2018).
- Murray, J. I., et al. Automated analysis of embryonic gene expression with cellular resolution in C. elegans. Nature Methods. 5 (8), 703-709 (2008).
- Murray, J. I., Bao, Z., Boyle, T. J., Waterston, R. H. The lineaging of fluorescently-labeled Caenorhabditis elegans embryos with StarryNite and AceTree. Nature Protocols. 1 (3), 1468-1476 (2006).
- Wu, Y., et al. Spatially isotropic four-dimensional imaging with dual-view plane illumination microscopy. Nature Biotechnology. 31 (11), 1032-1038 (2013).
- Kumar, A., et al. Using Stage- and Slit-Scanning to Improve Contrast and Optical Sectioning in Dual-View Inverted Light Sheet Microscopy (diSPIM). The Biological Bulletin. 231 (1), 26-39 (2016).
- Kumar, A., et al. Dual-view plane illumination microscopy for rapid and spatially isotropic imaging. Nature Protocols. 9 (11), 2555-2573 (2014).
- Wu, Y., Christensen, R., Colon-Ramos, D., Shroff, H. Advanced optical imaging techniques for neurodevelopment. Current Opinion in Neurobiology. 23 (6), 1090-1097 (2013).
- Wu, Y., et al. Inverted selective plane illumination microscopy (iSPIM) enables coupled cell identity lineaging and neurodevelopmental imaging in Caenorhabditis elegans. Proceedings of the National Academy of Sciences of the United States of America. 108 (43), 17708-17713 (2011).
- Huisken, J., Stainier, D. Y. Selective plane illumination microscopy techniques in developmental biology. Development. 136 (12), 1963-1975 (2009).
- Stelzer, E. H. Light-sheet fluorescence microscopy for quantitative biology. Nature Methods. 12 (1), 23-26 (2015).
- Winter, P. W., Shroff, H. Faster fluorescence microscopy: advances in high speed biological imaging. Current Opinion in Chemical Biology. 20, 46-53 (2014).
- Lucy, L. B. An iterative technique for the rectification of observed distributions. Astronomical Journal. 76 (6), 745-754 (1974).
- Richardson, W. H. Bayesian-Based Iterative Method of Image Restoration. JOSA. 62 (1), 55-59 (1972).
- Stiernagle, T.
- Hart, A. C.
- Edelstein, A., Amodaj, N., Hoover, K., Vale, R., Stuurman, N. Computer control of microscopes using microManager. Current Protocols in Molecular Biology. Chapter 14, Unit14 20 (2010).
- Gualda, E. J., et al. SPIM-fluid: open source light-sheet based platform for high-throughput imaging. Biomedical Optics Express. 6 (11), 4447-4456 (2015).
- Ardiel, E. L., et al. Visualizing Calcium Flux in Freely Moving Nematode Embryos. Biophysical Journal. 112 (9), 1975-1983 (2017).
- Walton, T., et al. The Bicoid class homeodomain factors ceh-36/OTX and unc-30/PITX cooperate in C. elegans embryonic progenitor cells to regulate robust development. PLoS Genetics. 11 (3), e1005003 (2015).
- Altun, Z. F. WormAtlas. , (2002).
- Wood, W. B. Embryology: In the nematode C. elegans. Cold Spring Harbor Laboratory Press. Chapter 8, 215-241 (1988).
- White, J. G., Southgate, E., Thomson, J. N., Brenner, S. The structure of the nervous system of the nematode Caenorhabditis elegans. Philosophical Transactions of the Royal Society B: Biological Sciences. 314 (1165), (1986).
- Bao, Z., Murray, J. I. Mounting Caenorhabditis elegans embryos for live imaging of embryogenesis. Cold Spring Harb Protoc. (9), (2011).
- Christensen, R. P., et al.
- Santella, A., et al. WormGUIDES: an interactive single cell developmental atlas and tool for collaborative multidimensional data exploration. BMC Bioinformatics. 16, 189 (2015).
- Dupuy, D., et al. A first version of the Caenorhabditis elegans Promoterome. Genome Research. 14 (10B), 2169-2175 (2004).
- Reece-Hoyes, J. S., et al. Insight into transcription factor gene duplication from Caenorhabditis elegans Promoterome-driven expression patterns. BMC Genomics. 8 (27), (2007).
- WormBase. , Available from: https://www.wormbase.org (2019).
- Lee, R. Y. N., et al. WormBase 2017: molting into a new stage. Nucleic Acids Research. 46 (D1), D869-D874 (2018).
