

Summary
Ce protocole fournit aux chercheurs une méthode rapide et indirecte de mesure de l'activité du facteur de transcription NF-B/AP-1, dépendante de TLR, dans une lignée de cellules macrophages murines en réponse à une variété de surfaces polymères et de couches protéiques adsorbées qui modélisent le microenvironnement de l'implant biomatériau.
Abstract
La réponse inflammatoire persistante d'hôte à un biomatériau implanté, connu sous le nom de réaction de corps étranger, est un défi significatif dans le développement et la mise en œuvre des dispositifs biomédicaux et des constructions d'ingénierie de tissu. Les macrophages, une cellule immunitaire innée, sont des acteurs clés dans la réaction du corps étranger parce qu'ils restent au site de l'implant pendant toute la durée de vie de l'appareil, et sont couramment étudiés pour acquérir une compréhension de cette réponse néfaste de l'hôte. De nombreux chercheurs en biomatériaux ont montré que les couches de protéines adsorbées sur les matériaux implantés influencent le comportement des macrophages, et par la suite ont un impact sur la réponse de l'hôte. Les méthodes de cet article décrivent un modèle in vitro utilisant des couches de protéines adsorbed contenant des molécules de dommages cellulaires sur les surfaces de biomatériaux polymères pour évaluer les réponses macrophages. Une lignée de cellules macrophages de journaliste de NF-B/AP-1 et l'essai colorimétrique associé de phosphatase alkaline ont été employés comme méthode rapide pour examiner indirectement l'activité de facteur de transcription de NF-B/AP-1 en réponse aux couches complexes de protéine adsorbée contenant des protéines sanguines et des modèles moléculaires dommages-associés, comme modèle des couches complexes de protéine adsorbed formées sur des surfaces de protéine de biomatériau in vivo.
Introduction
La réaction du corps étranger (FBR) est une réponse chronique de l'hôte qui peut avoir un impact négatif sur la performance d'un matériau ou d'un dispositif implanté (p. ex. dispositifs d'administration de médicaments, biocapteurs), par la libération persistante de médiateurs inflammatoires et en empêchant l'intégration entre le matériau implanté et le tissu environnant1. Cette réponse immunitaire innée est initiée par la procédure d'implantation et se caractérise par la présence à long terme de cellules immunitaires innées et la formation de capsules fibreuses autour de l'implant1. Dans le contexte des réponses matérielles de l'hôte, les interactions macrophage-matérielle ont un impact significatif sur la progression de la réponse de l'hôte et le développement d'un FBR1. Les macrophages sont une population de cellules immunitaires innées diversifiées, recrutées au site de l'implant soit à partir de populations de macrophages résident de tissus ou du sang comme macrophages dérivés de monocytes. Ils commencent à s'accumuler sur le site de l'implant peu de temps après l'implantation, et en quelques jours deviennent la population cellulaire prédominante dans le microenvironnement implantaire. Les macrophages matériels, ainsi que les cellules géantes du corps étranger (FBGC) formées par la fusion des macrophages, peuvent persister à la surface du matériau pendant toute la durée de vie de l'implant2,3. Par conséquent, les macrophages sont considérés comme des acteurs clés dans la réponse du corps étranger en raison de leurs rôles orchestrant les étapes caractéristiques de la FBR: réponse inflammatoire aigue, remodelage des tissus, et la formation de tissu fibrotique1.
Les récepteurs de péage-comme (TLRs) sont une famille des récepteurs de reconnaissance de modèle qui sont exprimés par beaucoup de cellules immunitaires, y compris des macrophages, et ont été montrés pour jouer un rôle significatif dans l'inflammation et la guérison de blessure. En plus des ligands dérivés d'agents pathogènes, les TLR sont capables de lier les molécules endogènes, connues sous le nom de modèles moléculaires associés aux dommages (DAMP), qui sont libérés pendant la nécrose cellulaire et d'activer les voies de signalisation inflammatoires entraînant la production de cytokines proinflammatoires4. Nous et d'autres avons proposé que les dommages encourus pendant les procédures d'implantation de biomatériaux mous libèrent des DAMPs, qui adsorbent alors aux surfaces de biomatériaux en plus des protéines de sang et modulent les interactions suivantes de cellules-matériel5,6. Lorsque les macrophages interagissent avec la couche protéique adsorbed sur un implant, leurs TLR de surface peuvent reconnaître les DAMP adsorbés et activer les cascades de signalisation pro-inflammatoire, conduisant à nf-B et AP-1 transcription facteur d'activation et de production de cytokines proinflammatoires. Nous avons précédemment montré que les macrophages murines ont considérablement augmenté l'activité de NF-B/AP-1 et le facteur de nécrose de tumeur (TNF-, cytokine proinflammatoire) sécrétion en réponse à DAMP-contenant des couches de protéines adsorbed sur une variété de surfaces polymères par rapport aux surfaces avec sérum adsorbed ou plasma seulement (c.-à-d., pas de DAMPs présents), et que cette réponse est largement médiée par TLR2, tandis que TLR4 joue un rôle moindre5.
La lignée de cellules macrophages de reporter NF-B/AP-1 (Tableau des matériaux) utilisée dans ce protocole est une méthode pratique pour mesurer l'activité relative de NF-B et d'AP-1 dans les macrophages5,7,8. En combinaison avec les inhibiteurs de voie de TLR, cette ligne de cellules est un outil utile pour étudier l'activation de TLR et son rôle dans l'inflammation en réponse à une série de stimulus5,7,8. Les cellules de journaliste sont une ligne cellulaire macrophage-like de souris modifiée qui peut produire de façon stable la phosphatase alcaline embryonnaire sécrétée (SEAP) sur l'activation de facteur de transcription NF-B et AP-19. L'analyse colorimétrique enzymatique de phosphatase alcaline(tableau des matériaux)peut alors être utilisée pour quantifier les quantités relatives d'expression SEAP comme mesure indirecte de l'activité NF-B/AP-1. Comme nf-B et AP-1 sont en aval de nombreuses voies de signalisation cellulaire, neutraliser les anticorps et les inhibiteurs ciblant des TLR spécifiques (p. ex. TLR2) ou des molécules adaptatrices TLR (p. ex. MyD88) peuvent être utilisés pour vérifier le rôle d'une voie spécifique. La méthodologie décrite dans cet article fournit une approche simple et rapide pour évaluer la contribution de la signalisation de TLR dans les réponses de macrophage murine à une série de surfaces polymères avec des couches de protéine adsorbées contenant des protéines de sang et des DAMPs comme modèle in vitro des biomatériaux implantés.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. Préparation des médias et des réactifs
- Préparer les médias fibroblastes. Combinez 450 ml du milieu Eagle modifié de Dulbecco (DMEM), 50 ml de sérum bovin fœtal (FBS) et 5 ml de pénicilline/streptomycine. Conserver à 4 oC jusqu'à 3 mois.
- Préparer les médias de croissance macrophage journaliste en 50 ml aliquots. Combiner 45 mL de DMEM, 5 mL de FBS, 5 g/mL de réactif d'élimination du mycoplasme(tableau des matériaux),et 200 g/mL de phléomycine D1(Tableau des matériaux). Conserver à 4 oC jusqu'à 3 mois.
- Préparer les médias d'exemple de macrophage de journaliste dans 50 mL aliquots. Combinez 45 ml de DMEM, 5 ml de FLeomycine inactivée par la chaleur (HI-FBS), 5 réagents d'élimination du mycoplasme de g/mL et 200 x g/mL de phléomycine D1. Conserver à 4 oC jusqu'à 3 mois.
2. Revêtement des surfaces de culture cellulaire avec poly (méthyle méthacrylate)
- Dissoudre le poly (méthyle méthacrylate) (PMMA) dans le chloroforme à 20 mg/mL (p. ex., 100 mg de PMMA dans 5 ml de chloroforme) dans une fiole de scintillation en verre de 20 ml. Placer une barre magnétique dans le flacon et laisser remuer pendant au moins 2 h, jusqu'à ce que tous les solides soient dissous.
MISE EN GARDE: Le chloroforme est nocif s'il est inhalé. Assurez-vous d'utiliser du solvant dans une hotte de fumée tout en portant des gants PVA. - Pipette 400 'L de solution PMMA sur le centre d'une lame de microscope en verre borosilicate dans un couchette de spin, et tourner à 3000 tr/min pendant 2 min. Préparer le nombre de diapositives nécessaires pour l'analyse, ainsi que 3 à 5 supplémentaires pour la mesure de l'angle de contact de l'eau. Rangez les glissières dans une boîte propre (pulvérisée et essuyée avec 70 % d'éthanol) pour une utilisation future.
REMARQUE: Le revêtement de spin est souvent employé pour déposer un revêtement mince et uniforme sur une surface plane. Un douilleur de spin fait pivoter un substrat à haute vitesse, en utilisant la force centrifuge pour étendre la solution de revêtement sur la surface.- Mesurer l'angle de contact de l'eau à deux positions aléatoires à la surface des toboggans supplémentaires enduits (c.-à-d. pas les diapositives utilisées pour la culture cellulaire) avec un goniomètre pour s'assurer que la surface de verre était complètement recouverte du polymère.
REMARQUE: Seule l'eau de la plus grande pureté (p. ex., verre triple distillé) doit être utilisée pour mesurer l'angle de contact de l'eau.
- Mesurer l'angle de contact de l'eau à deux positions aléatoires à la surface des toboggans supplémentaires enduits (c.-à-d. pas les diapositives utilisées pour la culture cellulaire) avec un goniomètre pour s'assurer que la surface de verre était complètement recouverte du polymère.
- Dans un coffret de sécurité biologique (BSC), attachez des puits collants à 8 chambres à des lames enduites PMMA à l'aide de forceps stériles et en suivant une technique aseptique. Appuyez fermement sur le dessus des puits collants pour s'assurer qu'ils sont fortement attachés. Incuber les lames avec des puits collants attachés à 37 oC pendant la nuit pour fixer le joint.
- Testez le joint des puits collants en ajoutant 200 L d'eau de qualité de culture cellulaire (sans endotoxine) à chaque puits. Incuber à température ambiante (RT) pendant 60 min et ne pas éviter de fuite avant de procéder. Aspirer l'eau, en prenant soin de ne pas déranger le revêtement PMMA.
- Effectuer des lavages d'eau sans endotoxine en ajoutant 300 l'eau sans endotoxine à chaque puits et en couve pendant 1 h (trois fois), 12 h et 24 h avant d'être utilisé pour enlever tout solvant restant.
- Testez la concentration d'endotoxines des diapositives à utiliser pour la culture cellulaire. Incuber 200 l d'eau de réactif sans endotoxine(Tableau des matériaux) dans un puits de chaque toboggan pendant 1 h. Mesurer la concentration d'endotoxinedanse dans l'extrait à l'aide d'un point de terminaison d'endotoxine chromogénique(Tableau des matériaux).
REMARQUE: Le protocole suivant est spécifique au kit d'analyse de l'endotoxine figurant dans le Tableau des matériaux. - N'utilisez que de l'eau et des consommables (c.-à-d. des pointes de pipette, des tubes de microcentrifuge et des plaques de puits) qui sont certifiés sans pyrogène (c.-à-d. sans endotoxine) pour ce travail. En outre, toute verrerie utilisée dans la préparation des surfaces recouvertes de polymères doit être dépyrogénéisée à l'aide de la stérilisation à la chaleur sèche (250 oC pendant 30 min) avant d'utiliser10. La mesure de l'endotoxine dans la solution d'extrait, comme décrit ici, peut avoir comme conséquence une sous-estimation de l'endotoxine sur la surface matérielle11,12. Par conséquent, il est recommandé que lors de l'élaboration d'un protocole de revêtement polymère, effectuer la réaction d'analyse de l'endotoxine (c.-à-d., étapes 2.5.4-2.5.6 pour les échantillons d'essai [l'eau de réactif] ou les contrôles de pointe) directement dans les puits contenant l'échantillon enduit pour s'assurer qu'aucune source d'endotoxine n'est introduite par inadvertance dans le système pendant le processus de revêtement.
- Apportez tous les échantillons d'essai (c.-à-d. extraits) et les réactifs d'analyse d'endotoxine à RT. Reconstituer le réactif chromogénique dans la norme de tampon d'essai et d'endotoxine dans l'eau de réactif, permettent de se dissoudre pendant 5 min et tourbillonnent doucement avant d'utiliser. Couvrir toutes les bouteilles de film de paraffine lorsqu'elle n'est pas utilisée.
- Créez une courbe de dilution standard de 5 à 8 points de la norme d'endotoxine allant de la limite inférieure à la limite supérieure de l'analyse en effectuant une dilution sérielle de la norme d'endotoxine dans l'eau de réactif.
- Pour contrôler l'amélioration ou l'inhibition de l'analyse de l'endotoxine dans les échantillons d'essai, préparez un contrôle positif (également appelé un contrôle de pointe ou un échantillon à pointes) en diluant une quantité connue d'endotoxine dans la solution d'échantillon d'essai inutilisée.
REMARQUE: La concentration du contrôle positif doit être la même concentration qu'une norme au milieu de la courbe standard. Si la quantité récupérée du pic d'endotoxine (c.-à-d., concentration du contrôle positif moins la concentration de l'échantillon d'essai non épilé) est dans 50-200% de la concentration nominale du pic d'endotoxine, la solution d'extraction peut être considérée pour ne pas interférer de manière significative avec l'essai. - Ajoutez 50 L de normes, d'échantillons ou de commandes de pointe à chaque puits d'une plaque de 96 puits en double ou en triple. Utilisez l'eau de réactif comme un contrôle négatif.
- Ajouter 50 l de réactif chromogénique à chaque puits. Ajouter le réactif rapidement à tous les puits. Utilisez une minuterie pour enregistrer le temps qu'il faut pour ajouter du réactif à tous les puits. Couvrir la plaque d'un joint adhésif et incuber à 37 oC (le temps d'incubation dépend du lot et indiqué sur le certificat d'analyse inclus dans le kit de réactif chromogénique). Vous pouvez également vérifier sur la plaque toutes les 15 min pendant l'incubation jusqu'à ce que le changement de couleur soit observé dans tous les puits standard.
- Après l'incubation, ajouter 25 ll d'acide acétique à 50 % de l'acide acétique à chaque puits (concentration finale de 10 % d'acide acétique par puits) pour arrêter la réaction. Ajouter de l'acide acétique dans le même ordre que le réactif chromogénique a été ajouté. Lire l'absorption de la plaque à l'aide d'un lecteur de plaque à 405 nm. Aspirer le liquide et jeter la plaque.
REMARQUE: L'ajout d'acide acétique devrait prendre le même temps à ajouter à chaque puits que le réactif chromogénique a pris (30 s).
- Ultraviolet (UV) stériliser les diapositives pendant 30 minutes avant les expériences de culture cellulaire.
3. Surfaces de culture cellulaire de revêtement avec polydimethylsiloxane
- Mélanger l'élastomère polydiméthylsiloxane (PDMS) dans un rapport de poids de 10:1 (agent de base : agent de durcissement). Dans un coffret de sécurité biologique, pipette approximativement 10 ml de base de polydiméthylsiloxane dans un tube stérile. Peser le tube et ajouter lentement l'agent de durcissement jusqu'à ce que 10% a été ajouté.
MISE EN GARDE: Utilisez des réactifs PDMS dans un endroit bien aéré et évitez le contact visuel en portant des lunettes de sécurité. - Mélanger soigneusement l'élastomère en remuant avec une pointe de pipette sérologique stérile et en faisant du vaillant de haut en bas. Ajouter environ 200 l de la solution à chaque puits d'une plaque de 48 puits. Inclinez la plaque de puits lentement pour assurer une couverture complète des puits avec une solution d'élastomère.
- Placer la plaque de puits avec de l'élastomère dans un four à vide réglé à 50 cmHg, 40 oC. Retirez le couvercle et couvrez-le d'une lingette à une seule pile pour empêcher d'autres débris de tomber dans les puits. Laisser incuber pendant au moins 48 h.
- Confirmer les puits sont complètement enduits par l'inspection visuelle. Assurez-vous que l'élastomère est entièrement guéri en appuyant doucement avec une pointe de pipette stérile avant d'enlever.
- Ajouter 300 l'éthanol de 70 % (fabriqué avec de l'éthanol absolu et de l'eau sans endotoxine) et couver à RT pendant 1 h. Enlever l'éthanol et effectuer des lavages d'eau sans endotoxine en ajoutant 300 l d'eau sans endotoxine à chaque puits et en couve pendant 1 h (trois fois), 12 h , et 24 h avant l'utilisation pour enlever tout solvant restant.
- Incuber 200 l'eau sans endotoxine dans trois puits de chaque assiette pendant 1 h. Mesurer la concentration endotoxine des extraits d'eau à l'aide d'un point de terminaison d'endotoxine chromogénique (étapes 2.5.1-2.5.6).
4. Surfaces de culture cellulaire de revêtement avec Poly fluoré (tétrafluoroethylene)
- Faire une solution de 1 mg/mL de poly fluoré (tétrafluoroéthylène) (fPTFE) (p. ex., ajouter 10 mg de fPTFE à 10 ml de solvant fluoré [Tableau des matériaux]) dans un flacon de scintillation en verre de 20 ml. Placer une barre magnétique dans le flacon et laisser remuer pendant au moins 24 h, jusqu'à ce que tous les solides soient dissous.
- Ajouter environ 150 l de la solution de polymère à chaque puits d'une plaque de 48 puits de polystyrène (c.-à-d. non traitée par la culture tissulaire). Inclinez la plaque de puits lentement pour assurer une couverture complète de tous les puits avec une solution de polymère. Remplacer le couvercle.
- Afin d'assurer une enrobage fPTFE efficace des puits, les couvercles en verre doivent être enduits dans le fPTFE et utilisés pour la mesure de l'angle de contact avec l'eau (étape 4.3.1). Placer les couvertures à l'intérieur des puits d'une plaque de 24 puits. Ajouter environ 400 L de la solution de polymère à chaque puits contenant un bordereau. Poussez les couvertures vers le bas à l'aide de forceps stériles, en vous assurant qu'ils sont complètement recouverts de solution de polymère, et couvrez la plaque de puits avec un couvercle.
- Placer la plaque de puits avec une solution de polymère et/ou des plaques d'eau dans un four à vide réglé à 50 cmHg, 40 oC. Retirez le couvercle et couvrez-le d'une lingette à une seule pile pour empêcher d'autres débris de tomber dans les puits. Laisser incuber pendant au moins 48 h.
- Mesurez l'angle de contact avec l'eau des couvertures enduites de fPTFE à l'aide d'un goniomètre afin d'assurer un revêtement efficace.
REMARQUE: Seule l'eau de la plus grande pureté (p. ex., verre triple distillé) doit être utilisée pour mesurer l'angle de contact de l'eau.
- Mesurez l'angle de contact avec l'eau des couvertures enduites de fPTFE à l'aide d'un goniomètre afin d'assurer un revêtement efficace.
- Ajouter 300 l'éthanol de 70 % (fabriqué avec de l'éthanol absolu et de l'eau sans endotoxine) et couver à RT pendant 1 h. Enlever l'éthanol et effectuer des lavages d'eau sans endotoxine en ajoutant 300 l d'eau sans endotoxine à chaque puits et en couve pendant 1 h (trois fois), 12 h , et 24 h avant l'utilisation pour enlever tout solvant restant.
- Incuber 200 l'eau sans endotoxine dans trois puits de chaque assiette pendant 1 h. Mesurer la concentration d'endotoxines d'extraits d'eau à l'aide d'un point de terminaison d'endotoxine chromogénique (étapes 2.5.1-2.5.6).
- Les UV stérilisent les plaques de puits pendant 30 minutes avant les expériences de culture cellulaire.
5. Fabrication de Lysate à partir de cellules 3T3
- Cultivez des cellules 3T3 dans plusieurs flacons T150 à 70% de confluence. Pour détacher les cellules, aspirer les médias, laver la surface avec 5 ml de PBS et aspirer le PBS. Ajouter 5 mL d'enzyme de dissociation cellulaire recombinante sans origine animale(tableau des matériaux)et incuber à 37 oC pendant 3 à 5 min.
- Détachez les cellules en inclinant doucement le flacon d'avant en arrière. Ajouter 5 ml de PBS pour neutraliser l'enzyme recombinante utilisée pour la dissociation cellulaire. Transférer les cellules détachées des flacons dans un tube de centrifugeuse et mélanger par pipetting. Effectuez un compte de cellules vivantes à l'aide d'un hémocytomètre et d'un colorant de viabilité cellulaire.
REMARQUE: Une enzyme de dissociation cellulaire qui peut être neutralisée par dilution dans PBS a été choisie pour éviter l'introduction de protéines à base de sérum dans la préparation du lysate. Si la trypsine est utilisée pour dissocier les cellules, elle doit être neutralisée avec une solution contenant du sérum, et un lavage PBS supplémentaire devrait être effectué pour réduire la quantité de protéines sériques transportées dans la préparation de lysate. - Centrifuger les cellules à 200 x g pendant 5 min. Aspirer les cellules supernatant et resuspendre les cellules en volume d'origine (c.-à-d. 10 mL x nombre de flacons) de PBS pour laver les supports restants. Répéter.
- Centrifuger les cellules à nouveau à 200 x g pendant 5 min et aspirer le supernatant. Ajouter le volume de PBS nécessaire pour atteindre une concentration cellulaire finale de 1 x 106 cellules/mL. Placer la solution cellulaire dans un congélateur de -80 oC jusqu'à ce que l'échantillon soit complètement congelé (au moins 2 h).
- Décongeler la solution cellulaire dans un bain d'eau de 37 oC. Une fois complètement décongelée, remettre la solution dans le congélateur de -80 oC jusqu'à ce qu'elle soit entièrement congelée. Répéter l'opération pour un total de 3 cycles de gel-dégel.
- Effectuer un test d'acide micro-biinchoninique (BCA) sur le lysate cellulaire à une variété de dilutions (p. ex. 1/100, 1/200, 1/500, 1/1000) pour déterminer la concentration en protéines. Diluer le lysate cellulaire à une concentration protéique de 468,75 g/mL, aliquot, et stocker à -80 oC pour une utilisation future.
REMARQUE: La concentration finale de protéines dans une plaque de 48 puits est de 125 g/cm2 (sur la base de la surface d'un puits, 0,75 cm2). - Effectuer une tache occidentale pour évaluer la présence de DAMP dans le lysate (p. ex., protéine de choc thermique 60 [HSP60], boîte de groupe à haute mobilité 1 [HMGB1]) en chargeant 40 à 60 g de protéines lysate dans un tampon de chargement sur un gel de polyacrylamide de 1,5 mm d'épaisseur et suivez la tache occidentale standard Procédures.
6. Évaluer l'effet des couches de protéines adsorbed et des récepteurs de péage sur l'activité NF-B des macrophages
REMARQUE: Pour un schéma du flux de travail expérimental et de la disposition des plaques, consultez la figure 1A et la figure 1 supplémentaire,respectivement.
- Cultivez des macrophages de reporter dans un flacon de taille appropriée à la confluence de 70%. Aspirez les supports, lavez la surface avec PBS et aspirez PBS. Ajouter l'enzyme de dissociation cellulaire recombinante et incuber à 37 oC pendant 8 min.
- Détachez les cellules en tapant fermement sur les côtés du flacon. Inactiver l'enzyme de dissociation cellulaire recombinante en ajoutant un volume égal de support de croissance (contenant 10% de FBS). Effectuez un compte de cellules vivantes à l'aide d'un hémocytomètre et d'un colorant de viabilité cellulaire.
REMARQUE: La viabilité prévue pour les macrophages de journaliste suivant une incubation de 8 min dans l'enzyme de dissociation de cellules est 90%. - Cellules centrifugeuses à 200 x g pendant 5 min. Aspirate supernatant et resuspendre dans le volume original de PBS pour laver les cellules. Centrifugeà encore et resuspendre les cellules à 7,3 x 105 cellules/mL dans les supports d'avertissement (contenant de la chaleur inactivée FBS).
- Suspension cellulaire séparée en 3 tubes différents : inhibiteur TLR4, anti-TLR2 et non traité. Incuber les cellules avec un inhibiteur TLR4 de 1 g/mL pendant 60 min à RT ou avec un anti-TLR2 de 50 g/mL pendant 30 min à RT.
- Ajouter 200 l de lysate, 10 % de FBS, 10 % de plasma commercial de souris(tableau des matériaux),ou un mélange des solutions protéiques à une plaque de 48 puits (ou équivalent) et laisser la protéine à l'adsorbe à 37 oC pour la quantité désirée de temps (c.-à-d., 30 min, 60 min, ou 24 h). Aspirez les solutions protéiques des puits, en utilisant une pipette Pasteur fraîche pour chaque solution protéique, et lavez les surfaces avec 250 L de PBS pendant 5 min. Aspirate PBS. Répéter l'opération pour un total de 3 lavages.
REMARQUE: Cette étape peut devoir être commencée plus tôt dans le protocole en fonction du temps d'adsorption désiré. Ajuster le protocole en conséquence. - Après la période d'incubation avec l'inhibiteur TLR4 ou anti-TLR2, les cellules pipettes à resuspendre. Ajouter 200 L de solution cellulaire à chaque puits.
- Pour l'état de contrôle positif de TLR2, ajoutez Pam3CSK4 à une concentration finale de 150 ng/mL. Pour l'état de contrôle positif tLR4, ajouter le lipopolysaccharide (LPS) à une concentration finale de 1,5 g/mL. Incuber les cellules à 37 oC pendant 20 h.
- Échantillondez 20 ll de supernatant de chaque puits et plaque en double dans une plaque de 96 puits. Inclure trois puits de 20 médias d'analyse ll comme un contrôle de fond. Ajouter 200 l de réagent d'assay de journaliste SEAP à chaque puits. Couvrir la plaque d'un joint adhésif et couver pendant 2,5 h à 37 oC.
REMARQUE: Le temps d'incubation peut varier en fonction des conditions expérimentales, et devrait être optimisé pour une forte différence d'absorption entre les puits de contrôle positifs et négatifs.- Transférer le reste du supernatant dans un tube de 1,5 ml (par puits). Centrifugeuse à 1 000 x g pendant 10 min pour granuler les débris. Transférer le supernatant dans un nouveau tube de 1,5 ml et entreposer à -80 oC. Analyser le supernatant pour la présence de cytokines proinflammatoires (p. ex., TNF-MD, interleukine 6) par l'intermédiaire de l'analyse immunosorbent enzymatique (ELISA).
- Retirer le joint adhésif. Lire l'absorption de la plaque à l'aide d'un lecteur de plaque à 635 nm. Aspirer le liquide et jeter la plaque.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Des méthodes de nettoyage des surfaces recouvertes de polymères ont été testées pour s'assurer qu'il n'y avait pas de perturbation du revêtement, ce qui serait considéré comme un changement dans l'angle de contact avec l'eau à un bordereau de verre non couché (Figure 2). Des lames de microscope recouvertes de PMMA dans 70 % d'éthanol pendant 1 h ont été trouvés pour enlever le revêtement PMMA(figure 2, panneau gauche), probablement en raison de la solubilité de PMMA dans 80 wt% d'éthanol13, donc les surfaces enduites de PMMA ont été nettoyées en utilisant 30 min de stérilisation UV seule. La concentration de PMMA pour le revêtement a été optimisée précédemment5. Un trempage d'éthanol de 1 h 70% a été utilisé pour nettoyer le PDMS, et la stérilisation UV a été négligée puisque la lumière UV peut causer la scission de chaîne et influencer les propriétés d'humidification de surface de PDMS14. Le trempage à l'éthanol à 70 % et la stérilisation UV n'ont pas influencé l'angle de contact avec l'eau des couvertures enduites de fPTFE(figure 2, panneau droit), de fait, les deux méthodes, successivement, ont été utilisées pour nettoyer les revêtements fPTFE. La méthode de revêtement fPTFE a déjà été décrite par le groupe Grainger15.
Une tache occidentale a été exécutée sur le lysate 3T3 pour s'assurer que les espèces de DAMP étaient présentes dans le mélange moléculaire complexe. Les résultats ont montré que les deux HMGB1 et HSP60, deux DAMP bien documentés16,17, étaient présents dans le lysate (Figure 1B). L'adsorption des ligands TLR du lysate sur les surfaces de polymère a été confirmée par la culture des macrophages de journaliste (non traités, TLR2 neutralisé, ou TLR4 inhibé) pendant 20 h sur les surfaces de polymères adsorbed protéine (c.-à-d., polystyrène traité par culture tissulaire [TCPS], PMMA, PDMS, fPTFE), puis évaluant indirectement l'activité NF-B/AP-1 basée sur la production de SEAP à l'aide d'un test enzymatique(figure 1C et figure 3). En outre, les macrophages de journaliste avaient considérablement augmenté l'activité de NF-B/AP-1 sur le lysate adsorbed comparé au FBS ou au plasma adsorbé et à aucune protéine pré-adsorbée (médias) (figure 4). TLR ligands synthétiques lipopeptide triacylated (Pam3CSK4, TLR2 ligand) et lipopolysaccharide (LPS, TLR4 ligand) ont été inclus comme contrôles positifs pour confirmer l'anticorps ou l'inhibiteur et l'essai fonctionnait correctement. La neutralisation de TLR2 a eu une réduction sensiblement plus forte de la réponse de NF-B/AP-1 des macrophages de journaliste au lysate adsorbed comparé à l'inhibition de TLR4. De plus, de petites quantités de lysate dilué dans le sérum (à base de protéines totales) ont entraîné une augmentation significative de la réponse nf-B/AP-1 par rapport au sérum seul, la plus faible dilution effective dépendant de la surface du polymère (figure 5). Ces résultats démontrent la puissance des molécules adsorbed lysate-dérivées sur induire TLR-dépendant NF-B/AP-1 activité dans les macrophages de reporter sur une variété de surfaces polymères.

Figure 1 : Méthodes et résultats pour l'analyse de phosphatase alcaline des macrophages de journaliste de NF-B/AP-1 sur TCPS, PMMA, PDMS, et fPTFE. (A) Diagramme du flux de travail pour l'analyse de phosphatase alcaline de macrophage de journaliste. (B) Tache occidentale de lysate confirmant la présence des espèces DAMP HMGB1 et HSP60, avec l'actine comme contrôle de chargement. (C) Activité NF-B/AP-1 (représentée par absorption) des macrophages de reporter cultivés sur les médias (contrôle négatif), 10% FBS, lysate, et Pam3CSK4 (TLR2 ligand, contrôle positif) pendant 20 h. Les données montrent les résultats d'une expérience et sont représentatifs des résultats d'au moins 2 expériences distinctes, présentées comme une déviation standard moyenne (SD). Chaque expérience utilisait des puits séparés n '3 par condition, et chaque puits était en double pour l'assay enzymatique. Analyse à l'aide d'un test post-hoc ANOVA et Tukey. p lt; 0,001. Ce chiffre a été adapté avec la permission de McKiel et Fitzpatrick5. Copyright 2018 American Chemical Society. Veuillez cliquer ici pour voir une version plus grande de ce chiffre.

Figure 2 : Optimisation des méthodes de nettoyage des surfaces enduites de PMMA et de fPTFE, évaluées à l'aide de l'angle de contact avec l'eau (WCA). Des mesures ont été prises sur 2 spots distincts d'au moins 3 couvertures. Les données sont présentées comme moyennes et SD. Analysées à l'aide d'un test anodin ANOVA et Tukey. p lt; 0,05. Ce chiffre a été adapté avec la permission de McKiel et Fitzpatrick5. Copyright 2018 American Chemical Society. Veuillez cliquer ici pour voir une version plus grande de ce chiffre.

Figure 3 : Activité NF-B/AP-1 (représentée par absorption) des macrophages de reporter cultivés sur 10% FBS (contrôle), lysate, et contrôle positif pendant 20 h. (A) Influence de la neutralisation de TLR2 sur des réponses de macrophage de journaliste au lysate adsorbed. Le contrôle positif est Pam (Pam3CSK4, TLR2 ligand). (B) Influence de l'inhibition de TLR4 sur la réponse de macrophage de journaliste au lysate adsorbed. Le contrôle positif est LPS (TLR4 ligand). Les données montrent les résultats d'une expérience et sont représentatives des résultats d'au moins 2 expériences distinctes, présentées comme moyennes - SD. Chaque expérience utilisait des puits séparés n '3 par condition, et chaque puits était en double pour l'assay enzymatique. Analyse à l'aide d'un test post-hoc ANOVA et Tukey. p 'lt; 0.01, 'p 'lt; 0.001. Ce chiffre a été adapté avec la permission de McKiel et Fitzpatrick5. Copyright 2018 American Chemical Society. Veuillez cliquer ici pour voir une version plus grande de ce chiffre.

Figure 4 : Activité NF-B/AP-1 (représentée par absorption) des macrophages de reporter cultivés sur les médias (contrôle négatif), les couches de protéines adsorbées de 30 min et 24 h, et pam3CSK4 (contrôle positif) sur TCPS pendant 20 h. Les données sont combinées à partir de 3 expériences distinctes et présentées comme moyennes et SD. Chaque expérience utilisait des puits de n à 3 puits distincts par condition, et chaque puits était en double pour l'évaluation enzymatique (c.-à-d. n'9 puits de culture cellulaire non indépendants et n 18 puits d'analysés enzymatiques non indépendants). Analyse à l'aide d'un test post-hoc ANOVA et Tukey. p lt; 0,001. Veuillez cliquer ici pour voir une version plus grande de ce chiffre.
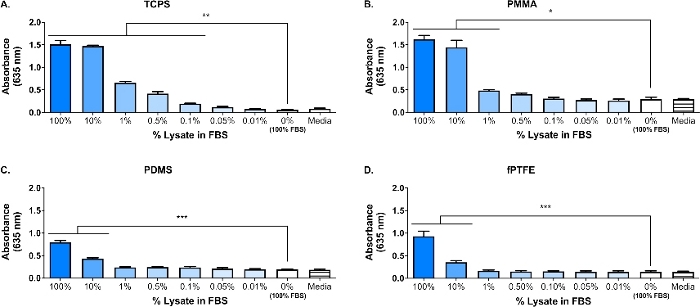
Figure 5 : Activité de macrophage reporter NF-B/AP-1 (représentée par absorption) après 20 h en réponse aux dilutions du lysate dans FBS (protéine totale de 280 g/puits) adsorbées aux surfaces polymères pendant 30 min. (A) TCPS. (B) PMMA. (C) PDMS. (D) fPTFE. Les données montrent les résultats d'une expérience et sont représentatives des résultats d'au moins 2 expériences distinctes, présentées comme moyennes - SD. Chaque expérience utilisait des puits séparés n '3 par condition, et chaque puits était en double pour l'assay enzymatique. Analyse à l'aide d'un test post-hoc ANOVA et Tukey. p 'lt; 0.05, 'p 'lt; 0.01, 'p 'lt; 0.001. Ce chiffre a été adapté avec la permission de McKiel et Fitzpatrick5. Copyright 2018 American Chemical Society. Veuillez cliquer ici pour voir une version plus grande de ce chiffre.

Figure supplémentaire 1 : Exemple s'agencent pour l'exemple de l'exemple utilisé pour l'assidui de culture de macrophage de journaliste NF-B/AP-1 dans les formats de plaque de 8-chambre et de 48 puits. Veuillez cliquer ici pour voir une version plus grande de ce chiffre.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
L'un des principaux objectifs de notre laboratoire est la réponse de l'hôte aux implants solides de tissus mous biomatériaux, et en particulier comment les dommages cellulaires encourus pendant la procédure d'implantation influe sur la réponse de l'hôte. Les travaux présentés ici décrivent des expériences préliminaires utilisant une lignée de cellules macrophages reporter et du lysate cellulaire contenant du DAMP généré sin in vitro, afin d'étudier l'influence des molécules libérées lors de dommages cellulaires (c.-à-d. de la chirurgie implantaire) sur les réponses aux macrophages aux biomatériaux. Le lysate de cellules de fibroblaste a été employé pour modéliser les dommages cellulaires et la libération des DAMP dus au placement de biomatériau. Les fibroblastes ont été choisis pour créer le lysate en raison de la prévalence des fibroblastes dans les tissus mous, ainsi que de leur capacité à sécréte une variété de protéines de matrice extracellulaire (ECM), y compris la fibronectine18. Le cycle de gel-dégel a été choisi comme méthode de lyse pour produire des DAMP intracellulaires et ECM-dérivés, semblables à ce qui serait présent dans l'environnement d'implant. Les inhibiteurs de la protéase n'ont pas été utilisés pour fabriquer ce lysate. Tandis que la lyse cellulaire incontrôlée comme le cycle de gel-dégel peut avoir comme conséquence la libération des protéases qui peuvent dégrader DESDAMP, ces enzymes seraient également probablement présentes dans l'environnement d'implant de biomatériau quand les cellules sont endommagées pendant la procédure d'implantation. La présence de DAMP dans le mélange moléculaire complexe du lysate a été confirmée par Western blot (Figure 1B; HMGB1 et HSP60) et SEAP reporter d'assay (Figure 1C; L'activité NF-B/AP-1 en réponse au lysate adsorbed). Nous avons également effectué des tests où le lysate a été dilué dans FBS basé sur la concentration totale de protéines et adsorbed sur les surfaces de culture cellulaire (Figure 5) pour mieux refléter la complexité de l'environnement implantaire, car il contiendra une abondance de protéines sanguines ainsi que DAMPs6. L'activité de macrophage de reporter NF-B/AP-1 est restée sensiblement augmentée sur les couches adsorbed du lysate dilué dans FBS, et la dilution la plus basse pour réaliser l'activation significative était dépendante de surface, s'étendant de 0.1% (TCPS) à 10% (PDMS et fPTFE).
Les polymères PMMA, PDMS, et PTFE ont été choisis pour ce travail parce qu'ils ne sont pas dégradables et ont été largement utilisés dans la littérature pour évaluer l'adsorption des protéines et la réponse macrophage aux biomatériaux19,20,21,22,23,24,25. TCPS a également été utilisé pour la comparaison car il est un substrat commun utilisé pour le macrophage in vitro et TLR travail de signalisation21,26,27,28. Les matériaux utilisés dans notre travail sont des exemples représentatifs de biomatériaux solides et non dégradables. Cependant, beaucoup d'autres matériaux pourraient être utilisés avec ce modèle, à condition que le matériel puisse être enduit sur des plaques de culture cellulaire ou des glissières de microscope et correctement décontaminé. La lignée de cellules macrophages des reporters NF-B/AP-1 a été sélectionnée pour ce modèle in vitro parce qu'elle permet une mesure rapide et indirecte de l'activité NF-B/AP-1 grâce à l'expression inductible NF-B/AP-1 du SEAP. Les macrophages de journaliste de NF-B/AP-1 exigent l'utilisation de la phléomycine D1 dans le média de culture comme antibiotique sélectif pour s'assurer que seules les cellules avec le gène SEAP inductible de NF-B/AP-1 sont présentes29. Pour l'analyse de phosphatase alcaline, il est essentiel d'utiliser HI-FBS dans le média de culture cellulaire pour éviter les résultats positifs potentiels faux générés par les phosphatases alcaline présentes dans le sérum. Nos recherches à ce jour suggèrent que les surfaces adsorbed FBS ne génèrent pas un résultat fausse ment positif détectable, probablement parce que les molécules de sérum sont fortement adsorbées à la surface de la culture et ne sont pas libérées dans le supernatant. Le point de temps de culture pour les macrophages de journaliste (20 h), le point de temps d'incubation d'essai (2,5 h), et la longueur d'onde de lecture d'absorption (635 nm) pour l'essai alcalin de phosphatase ont été optimisés avec ce système pour assurer des mesures robustes et reproductibles pour toutes les conditions.
Un point de temps initial d'adsorption de protéine de 30 min a été choisi pour ce travail dû à son utilisation commune dans la littérature d'adsorption de protéine (figure 1C)30,31,32,33,34. Cependant, nous avons également exploré des temps d'adsorption plus longs (c.-à-d., 60 min et 24 h, figure 4) pour mieux représenter la couche de protéine adsorbée que les macrophages interagiraient avec in vivo, qui est susceptible de se produire 4-24 h après l'implantation1. Il a été postulé que la majorité de l'adsorption et l'échange de protéines se produit dans les 60 premières min d'exposition à une surface26,35,36, donc un temps d'adsorption de 60 min peut être un point de temps plus pertinent. Nous sommes également passés de l'utilisation de FBS comme un contrôle négatif pour la présence de DAMP dans la couche de protéines adsorbed au plasma de souris commerciale. La raison d'être de l'utilisation du plasma au lieu du sérum est que les protéines plasmatiques sont connues pour jouer un rôle important dans l'adsorption des protéines et la réponse macrophage1, et que le plasma fournit une meilleure représentation des protéines dans l'environnement des plaies. Le plasma utilisé dans les expériences d'adsorption protéique est généralement préparé sous forme de dilution de 1 à 10 %26,36,37, ce qui a motivé notre utilisation de 10 % de plasma. Le plasma humain est couramment utilisé26,36, car il est plus facile à obtenir en grandes quantités et plus cliniquement pertinent, par rapport au plasma de souris. Cependant, nous avons choisi d'utiliser le plasma de souris commerciale pour dans ce modèle pour garder l'espèce des solutions protéiques compatibles avec celle des cellules reporter.
L'utilisation de la lignée de cellules macrophages reporter introduit certaines limites dans l'étude. Tout d'abord, l'utilisation d'une lignée de cellules macrophages leucémiques murine a des limites inhérentes, car le phénotype et le comportement peuvent varier des cultures macrophages primaires. Bien que cette limitation sera abordée dans les travaux futurs à l'aide de macrophages primaires, la lignée cellulaire macrophage parentale a été montré pour imiter étroitement macrophages de moelle osseuse de souris en termes de leurs récepteurs de surface cellulaire et la réponse aux ligands microbiens pour Les TLR 2, 3 et 438. En outre, les macrophages de journaliste de NF-B/AP-1 ont donné des résultats semblables en réponse à la stimulation de HMGB1 et de LPS comparés aux macrophages primaires péritonés39. Il convient de noter que les macrophages de journaliste NF-B/AP-1, et leur tension parentale, n'expriment pas TLR540. Les chercheurs ont démontré que HMGB1 était capable d'activer les facteurs de transcription NF-B via les voies de signalisation TLR5 dans les cellules HEK-293 poignardée transfectée avec l'homme TLR541. Par conséquent, la contribution de la signalisation HMGB1-TLR5 à l'activité globale de NF-B sur les surfaces enduites de lysate a été négligée dans ce modèle. En outre, les macrophages de journaliste et leur souche parentale n'expriment pas la protéine adaptatrice d'ASC, et par conséquent ne forment pas la plupart des types d'inflammatoires et ne peuvent pas traiter l'IL-1 ou l'IL-18 inactif inactif à leurs formes mûres42. Par conséquent, le modèle que nous avons utilisé ne tient pas compte de la contribution de l'activité inflammasome dépendante de l'ASC et de la signalisation autocrine IL-1 et IL-18 subséquente dans les réponses macrophages aux surfaces adsorbes de lysate. Par conséquent, cet analyse se veut un examen préliminaire de l'activation nf-B dépendante de TLR, et la recherche ultérieure utilisant des macrophages primaires est recommandée pour fournir une compréhension plus complète et représentative de l'activation et du phénotype de macrophage sur les surfaces matérielles d'intérêt.
L'essai alcalin de phosphatase mesure indirectement l'activité de NF-B/AP-1 des macrophages de journaliste. Cependant, il existe de nombreuses voies de signalisation autres que les TLR qui impliquent NF-B/AP-1 (par exemple, le récepteur de l'interleukine-1 [IL-1R]43 et le récepteur du facteur de nécrose tumorale [TNFR]44). Par conséquent, il était nécessaire d'évaluer la contribution de la signalisation TLR2 et TLR4 dans la réponse accrue de NF-B/AP-1 aux surfaces lysate-adsorbed utilisant des essais d'inhibition (figure 3). La raison d'être de la sélection de ces deux TLR de surface était qu'au moins 23 DAMP qui ont été montrés pour signaler par TLR2 et TLR445, y compris le HMGB1 bien caractérisé, et les deux récepteurs sont exprimés sur la surface cellulaire et peuvent interagir directement avec la surface du biomatériau6. Les essais d'inhibition de TLR2 et de TLR4 ont démontré que lorsque la signalisation de TLR2 ou de TLR4 a été bloquée, la réponse de NF-B/AP-1 des macrophages de journaliste au lysate adsorbé a été réduite, indiquant que les deux voies sont impliquées. Cependant, il y avait une réduction sensiblement plus grande de l'activité de NF-B/AP-1 quand la signalisation de TLR2 a été neutralisée, suggérant que TLR2 puisse jouer un rôle primaire dans la réponse des macrophages de journaliste au lysate adsorbed. Nous reconnaissons qu'il peut y avoir une certaine inhibition hors cible avec la voie de signalisation TLR neutralisant les anticorps et les inhibiteurs. Un anticorps neutralisant a été employé pour inhiber la voie de TLR2 puisqu'il n'y avait pas des molécules d'inhibiteur de TLR2 disponibles dans le commerce au moment de ce travail.
Les méthodes présentées ici utilisent le lysate, comme source complexe de DAMP, et les macrophages de reporter de NF-B/AP-1 comme modèle in vitro pour des réponses de macrophage aux DAMPs et à d'autres protéines adsorbées aux biomatériaux polymères (figure 1). Nous prévoyons que notre protocole peut être utilisé pour analyser rapidement les réponses NF-B/AP-1 et la signalisation en amont des macrophages des journalistes à une variété de matériaux (y compris les matériaux dégradables, les échafaudages poreux ou les hydrogels) et les couches de protéines adsorbed (figure 3). Cependant, l'utilisation de matériaux poreux et d'hydrogels introduira de la complexité au sein du système, car il peut être difficile de faire la distinction entre les molécules adsorbed et les molécules entraînées. Nous prévoyons également que ce protocole peut être facilement adapté pour étudier la contribution d'autres voies de signalisation en amont de NF-B/AP-1 (p. ex., récepteurs de lectine de type C46 et récepteurs d'oligomerisation liant les nucléotides (NOD)récepteurs 47) avec les inhibiteurs appropriés. En outre, la réponse NF-B/AP-1 des macrophages de journaliste pourrait être comparée entre différents matériaux, à condition que les réponses soient normalisées à l'activité cellulaire de base (c.-à-d., cellules dans les médias sur chaque surface sans protéine pré-adsorbée) et tous les matériaux ont des niveaux indétectables d'endotoxine.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Les auteurs n'ont rien à révéler.
Acknowledgments
Les auteurs remercient le financement opérationnel du Projet de recherche des Instituts de santé du Canada (PTJ 162251), du Comité consultatif du Sénat de l'Université Queen's et du soutien à l'infrastructure du Fonds de leadership de la Fondation canadienne pour l'innovation John Evan (projet 34137) et du Fonds de recherche du ministère de la Recherche et de l'Innovation de l'Ontario (projet 34137). L.A.M. a reçu l'appui d'une bourse R. Samuel McLaughlin de l'Université Queen's, d'une maîtrise en études supérieures du Conseil canadien des études supérieures en sciences naturelles et en génie du Canada et d'une bourse d'études supérieures de l'Ontario. Les auteurs tient à remercier le Dr Myron Szewczuk pour son généreux don de la lignée de cellules macrophages journaliste NF-B/AP-1 et les Drs Michael Blennerhassett et Sandra Lourenssen pour l'utilisation de leur système d'imagerie gel et lecteur de plaques.
Materials
| Name | Company | Catalog Number | Comments |
| Cell culture reagents | |||
| anti-mouse/human CD282 (TLR2) | Biolegend | 121802 | |
| CLI-095 (TLR4 inhibitor) | Invivogen | TLRL-CLI95 | |
| C57 complement plasma K2 EDTA 10ml, innovative grade US origin | InnovativeResearch | IGMSC57-K2 EDTA-Compl-10ml | Mouse plasma |
| Dulbecco's modified eagle medium (DMEM) | Sigma Aldrich | D6429-500ML | |
| Dulbecco's phosphate buffered saline (DPBS) | Fisher Scientific | 14190250 | No calcium, no magnesium |
| Fetal bovine serum (FBS), research grade | Wisent | 98150 | |
| LPS-EK | Invivogen | TLRL-EKLPS | Lipopolysaccharide from Escherichia coli K12 |
| NIH/3T3 fibroblasts | ATCC | CRL-1658 | |
| Pam3CSK4 | Invivogen | tlrl-pms | Synthetic triacylated lipopeptide - TLR1/2 ligand |
| Penicillin/streptomycin | Sigma Aldrich | P4333-100ML | |
| Plasmocin | Invivogen | ANT-MPP | Mycoplasma elimination reagent |
| RAW-Blue cells | Invivogen | raw-sp | NF-κB/AP-1 reporter macrophage cell line |
| Trypan blue solution, 0.4% | Fisher Scientific | 15250061 | |
| TrypLE express enzyme (1X) | Fisher Scientific | 12604021 | animal origin-free recombinant cell dissociation enzyme |
| Zeocin | Invivogen | ANT-ZN-1 | |
| Kits and assays | |||
| ELISA precoated plates, mouse IL-6 | Biolegend | B213022 | |
| ELISA precoated plates, mouse TNF-α | Biolegend | B220233 | |
| Endotoxin (Escherichia coli) - Control standard endotoxin (CSE) | Associates of Cape Cope Inc. | E0005-5 | Endotoxin for standard curve in chromogenic endotoxin assay |
| LAL water, 100 mL | Associates of Cape Cope Inc. | WP1001 | Used with chromogenic endotoxin assay |
| Micro BCA protein assay | Fisher Scientific | PI23235 | |
| Limulus amebocyte lysate (LAL) Pyrochrome endotoxin test kit | Associates of Cape Cope Inc. | C1500-5 | Chromogenic endotoxin assay reagent |
| QUANTI-Blue alkaline phosphatase detection medium | Invivogen | rep-qb2 | Alkaline phosphatase assay to indirectly measure NF-κB/AP-1 activity |
| Polymeric coating reagents | |||
| Chloroform, anhydrous | Sigma Aldrich | 288306-1L | |
| Ethyl alcohol anhydrous | Commercial Alcohols | P006EAAN | Sigma: Reagent alcohol, anhydrous, 676829-1L |
| Straight tapered fine tip forceps | Fisher Scientific | 16-100-113 | |
| Fluorinert FC-40 solvent | Sigma Aldrich | F9755-100ML | Fluorinated solvent for fPTFE |
| Cell culture grade water (endotoxin-free) | Fisher Scientific | SH30529LS | |
| Poly(methyl methacrylate) (PMMA) | Sigma Aldrich | 182230-25G | |
| Sylgard 184 elastomer kit | Fisher Scientific | 50822180 | |
| Teflon-AF (fPTFE) | Sigma Aldrich | 469610-1G | Poly[4,5-difluoro-2,2-bis(trifluoromethyl)-1,3-dioxole-co-tetrafluoroethylene] |
| Consumables | |||
| Adhesive plate seals | Fisher Scientific | AB-0580 | |
| Axygen microtubes, 1.5 mL | Fisher Scientific | 14-222-155 | |
| Borosilicate glass scintillation vials, with white polypropylene caps | Fisher Scientific | 03-337-4 | |
| Clear PS 48-well plate | Fisher Scientific | 08-772-52 | |
| Clear TCPS 96-well plate | Fisher Scientific | 08-772-2C | |
| Clear TCPS 48-well plate | Fisher Scientific | 08-772-1C | |
| Cover glasses, circles | Fisher Scientific | 12-545-81 | |
| Falcon tissue culture treated flasks, T25 | Fisher Scientific | 10-126-10 | |
| sticky-Slide 8 Well | Ibidi | 80828 | |
| Superfrost microscope slides | Fisher Scientific | 12-550-15 | |
| Tissue culture treated flasks, T150 | Fisher Scientific | 08-772-48 |
References
- Anderson, J. M., Rodriguez, A., Chang, D. T. Foreign body reaction to biomaterials. Seminars in Immunology. 20 (2), 86-100 (2008).
- Anderson, J. M., Miller, K. M.
- Collier, T. O., Anderson, J. M. Protein and surface effects on monocyte and macrophage adhesion, maturation, and survival. Journal of Biomedical Materials Research. 60 (3), 487-496 (2002).
- Bianchi, M. E. DAMPs, PAMPs and alarmins: all we need to know about danger. Journal of Leukocyte Biology. 81 (1), 1-5 (2007).
- McKiel, L. A., Fitzpatrick, L. E. Toll-like Receptor 2-Dependent NF-κB/AP-1 Activation by Damage-Associated Molecular Patterns Adsorbed on Polymeric Surfaces. ACS Biomaterials Science & Engineering. 4 (11), 3792-3801 (2018).
- Babensee, J. E. Interaction of dendritic cells with biomaterials. Seminars in Immunology. 20 (2), 101-108 (2008).
- Sintes, J., Romero, X., de Salort, J., Terhorst, C., Engel, P. Mouse CD84 is a pan-leukocyte cell-surface molecule that modulates LPS-induced cytokine secretion by macrophages. Journal of Leukocyte Biology. 88 (4), 687-697 (2010).
- Tom, J. K., Mancini, R. J., Esser-Kahn, A. P. Covalent modification of cell surfaces with TLR agonists improves and directs immune stimulation. Chemical Communications. 49 (83), 9618-9620 (2013).
- Abdulkhalek, S., et al. Neu1 sialidase and matrix metalloproteinase-9 cross-talk is essential for toll-like receptor activation and cellular signaling. Journal of Biological Chemistry. 286 (42), 36532-36549 (2011).
- Gorbet, M. B., Sefton, M. V.
- Xing, Z., Pabst, M. J., Hasty, K. A., Smith, R. A. Accumulation of LPS by polyethylene particles decreases bone attachment to implants. Journal of Orthopaedic Research. 24 (5), 959-966 (2006).
- Ding, H., et al. Comparison of the cytotoxic and inflammatory responses of titanium particles with different methods for endotoxin removal in RAW264.7 macrophages. Journal of Materials Science: Materials in Medicine. 23 (4), 1055-1062 (2012).
- Hoogenboom, R., Becer, C. R., Guerrero-Sanchez, C., Hoeppener, S., Schubert, U. S. Solubility and thermoresponsiveness of PMMA in alcohol-water solvent mixtures. Australian Journal of Chemistry. 63 (8), 1173-1178 (2010).
- Efimenko, K., Wallace, W. E., Genzer, J. Surface modification of Sylgard-184 poly(dimethyl siloxane) networks by ultraviolet and ultraviolet/ozone treatment. Journal of Colloid and Interface Science. 254 (2), 306-315 (2002).
- Godek, M. L., Sampson, J. A., Duchsherer, N. L., McElwee, Q., Grainger, D. W. Rho GTPase protein expression and activation in murine monocytes/macrophages is not modulated by model biomaterial surfaces in serum-containing in vitro cultures. Journal of Biomaterials Science. Polymer Edition. 17 (10), 1141-1158 (2006).
- Park, J. S., et al. Involvement of Toll-like Receptors 2 and 4 in Cellular Activation by High Mobility Group Box 1 Protein. Journal of Biological Chemistry. 279 (9), 7370-7377 (2004).
- Ohashi, K., Burkart, V., Flohé, S., Kolb, H. Cutting Edge: Heat Shock Protein 60 Is a Putative Endogenous Ligand of the Toll-Like Receptor-4 Complex. The Journal of Immunology. 164 (2), 558-561 (2000).
- Wong, T., McGrath, J. A., Navsaria, H. The role of fibroblasts in tissue engineering and regeneration. British Journal of Dermatology. 156 (6), 1149-1155 (2007).
- van Wachem, P. B., et al. The influence of protein adsorption on interactions of cultured human endothelial cells with polymers. Journal of Biomedical Materials Research. 21 (6), 701-718 (1987).
- Miller, K. M., Anderson, J. M. Human monocyte/macrophage activation and interleukin 1 generation by biomedical polymers. Journal of Biomedical Materials Research. 22 (8), 713-731 (1988).
- Bonfield, T. L., Colton, E., Anderson, J. M. Plasma protein adsorbed biomedical polymers: Activation of human monocytes and induction of interleukin 1. Journal of Biomedical Materials Research. 23 (6), 535-548 (1989).
- González, O., Smith, R. L., Goodman, S. B. Effect of size, concentration, surface area, and volume of polymethylmethacrylate particles on human macrophages in vitro. Journal of Biomedical Materials Research. 30 (4), 463-473 (1996).
- Anderson, J. M., et al. Protein adsorption and macrophage activation on polydimethylsiloxane and silicone rubber. Journal of Biomaterials Science. Polymer Edition. 7 (2), 159-169 (1995).
- Lord, M. S., Foss, M., Besenbacher, F. Influence of nanoscale surface topography on protein adsorption and cellular response. Nano Today. 5 (1), 66-78 (2010).
- Chen, S., et al. Characterization of topographical effects on macrophage behavior in a foreign body response model. Biomaterials. 31 (13), 3479-3491 (2010).
- Shen, M., Horbett, T. A. The effects of surface chemistry and adsorbed proteins on monocyte/macrophage adhesion to chemically modified polystyrene surfaces. Journal of Biomedical Materials Research. 57 (3), 336-345 (2001).
- Love, R. J., Jones, K. S. The recognition of biomaterials: Pattern recognition of medical polymers and their adsorbed biomolecules. Journal of Biomedical Materials Research Part A. 101 (9), 2740-2752 (2013).
- McNally, A. K., Anderson, J. M. Phenotypic expression in human monocyte-derived interleukin-4-induced foreign body giant cells and macrophages in vitro: Dependence on material surface properties. Journal of Biomedical Materials Research Part A. 103 (4), 1380-1390 (2015).
- Gambhir, V., et al. The TLR2 agonists lipoteichoic acid and Pam3CSK4 induce greater pro-inflammatory responses than inactivated Mycobacterium butyricum. Cellular Immunology. 280 (1), 101-107 (2012).
- Suzuki, O., Yagishita, H., Yamazaki, M., Aoba, T. Adsorption of Bovine Serum Albumin onto Octacalcium Phosphate and its Hydrolyzates. Cells and Materials. 5 (1), 45-54 (1995).
- Johnston, R. L., Spalton, D. J., Hussain, A., Marshall, J. In vitro protein adsorption to 2 intraocular lens materials. Journal of Cataract and Refractive Surgery. 25 (8), 1109-1115 (1999).
- Jin, J., Jiang, W., Yin, J., Ji, X., Stagnaro, P. Plasma proteins adsorption mechanism on polyethylene-grafted poly(ethylene glycol) surface by quartz crystal microbalance with dissipation. Langmuir. 29 (22), 6624-6633 (2013).
- Swartzlander, M. D., et al. Linking the foreign body response and protein adsorption to PEG-based hydrogels using proteomics. Biomaterials. 41, 26-36 (2015).
- Chamberlain, M. D., et al. Unbiased phosphoproteomic method identifies the initial effects of a methacrylic acid copolymer on macrophages. Proceedings of the National Academy of Sciences. 112 (34), 10673-10678 (2015).
- Dillman, W. J., Miller, I. F. On the adsorption of serum proteins on polymer membrane surfaces. Journal of Colloid And Interface Science. 44 (2), 221-241 (1973).
- Ishihara, K., Ziats, N. P., Tierney, B. P., Nakabayashi, N., Anderson, J. M. Protein adsorption from human plasma is reduced on phospholipid polymers. Journal of Biomedical Materials Research. 25 (11), 1397-1407 (1991).
- Warkentin, P., Wälivaara, B., Lundström, I., Tengvall, P. Differential surface binding of albumin, immunoglobulin G and fibrinogen. Biomaterials. 15 (10), 786-795 (1994).
- Berghaus, L. J., et al. Innate immune responses of primary murine macrophage-lineage cells and RAW 264.7 cells to ligands of Toll-like receptors 2, 3, and 4. Comparative Immunology, Microbiology and Infectious Diseases. 33 (5), 443-454 (2010).
- Zhang, Y., Karki, R., Igwe, O. J. Toll-like receptor 4 signaling: A common pathway for interactions between prooxidants and extracellular disulfide high mobility group box 1 (HMGB1) protein-coupled activation. Biochemical Pharmacology. 98 (1), 132-143 (2015).
- Mizel, S. B., Honko, A. N., Moors, M. A., Smith, P. S., West, A. P. Induction of macrophage nitric oxide production by Gram-negative flagellin involves signaling via heteromeric Toll-like receptor 5/Toll-like receptor 4 complexes. Journal of Immunology. 170 (12), 6217-6223 (2003).
- Das, N., et al. HMGB1 Activates Proinflammatory Signaling via TLR5 Leading to Allodynia. Cell Reports. 17 (4), 1128-1140 (2016).
- Pelegrin, P., Barroso-Gutierrez, C., Surprenant, A. P2X7 Receptor Differentially Couples to Distinct Release Pathways for IL-1β in Mouse Macrophage. The Journal of Immunology. 180 (11), 7147-7157 (2008).
- Tak, P. P., Firestein, G. S. NF-κB: A key role in inflammatory diseases. Journal of Clinical Investigation. 107 (1), 7-11 (2001).
- Ashkenazi, A., Dixit, V. M. Death receptors: signaling and modulation. Science. 281 (5381), 1305-1308 (1998).
- Erridge, C. Endogenous ligands of TLR2 and TLR4: agonists or assistants. Journal of Leukocyte Biology. 87 (6), 989-999 (2010).
- Feng, Y., et al. A macrophage-activating, injectable hydrogel to sequester endogenous growth factors for in situ angiogenesis. Biomaterials. 134, 128-142 (2017).
- Lonez, C., et al. Cationic lipid nanocarriers activate Toll-like receptor 2 and NLRP3 inflammasome pathways. Nanomedicine: Nanotechnology, Biology, and Medicine. 10 (4), 775-782 (2014).
