

Summary
Denne protokollen gir forskere en rask, indirekte metode for å måle TLR-avhengige NF-кB/AP-1 transkripsjon faktor aktivitet i en murine macrophage cellelinje som svar på en rekke polymer overflater og adsorberes protein lag som modell biomaterialet implantatet mikromiljøet.
Abstract
Den vedvarende inflammatorisk vert respons på en implantert biomaterialet, kjent som den utenlandske kroppen reaksjonen, er en betydelig utfordring i utvikling og implementering av biomedisinsk utstyr og vev engineering konstruksjoner. Makrofager, en medfødt immun celle, er sentrale aktører i den utenlandske kroppen reaksjonen fordi de forblir på implantatet området for levetiden til enheten, og er vanligvis studert for å få en forståelse av dette ødeleggende verten respons. Mange Biomaterials forskere har vist at adsorberes protein lag på implantert materiale påvirke macrophage atferd, og deretter påvirke verten respons. Metodene i dette dokumentet beskriver en in vitro-modell ved hjelp av adsorberes protein lag som inneholder cellulære skader molekyler på polymer biomaterialet overflater for å vurdere macrophage responser. En NF-кB/AP-1 reporter macrophage cellelinje og tilhørende fargemetrisk alkalisk fosfatase-analysen ble brukt som en rask metode for å indirekte undersøke NF-кB/AP-1 transkripsjon faktor aktivitet som svar på komplekse adsorberes protein lag som inneholder Blodproteiner og skade-forbundet molekylære mønstre, som en modell av komplekse adsorberes protein lag dannet på biomaterialet overflater in vivo.
Introduction
Den fremmedlegeme reaksjonen (FBR) er en kronisk vert respons som kan negativt påvirke ytelsen til et implantat materiale eller enhet (for eksempel medikamentlevering enheter, biosensors), gjennom vedvarende frigjøring av inflammatoriske meglere og ved å hindre integrasjon mellom implantert materiale og omkringliggende vev1. Denne medfødte immunresponsen er initiert av implantation prosedyren og er karakterisert ved den langsiktige tilstedeværelsen av medfødte immunceller og fiber kapsel formasjon rundt implantatet1. I forbindelse med materielle verts responser har macrophage-interaksjoner en betydelig innvirkning på progresjonen av verts responsen og utviklingen av en FBR1. Makrofager er en mangfoldig iboende immun celle befolkning, rekruttert til implantatet området enten fra vev-Resident macrophage populasjoner eller fra blodet som monocytt-avledet makrofager. De begynner å akkumulere på implantatet stedet kort tid etter implantation, og i løpet av dager blitt den dominerende cellen befolkningen i implantatet mikromiljøet. Materiale-tilhenger makrofager, sammen med fremmedlegeme gigantiske celler (FBGC) dannet gjennom macrophage fusjon, kan vedvare på materialets overflate for levetiden til implantatet2,3. Følgelig er makrofager anses å være sentrale aktører i fremmedlegeme respons på grunn av deres roller orkestrering de karakteristiske trinnene i FBR: akutt inflammatorisk respons, vev remodeling, og dannelse av antifibrotiske vev1.
Toll-lignende reseptorer (TLRs) er en familie av mønstergjenkjenning reseptorer som uttrykkes av mange immunceller, inkludert makrofager, og har vist å spille en betydelig rolle i betennelser og sår helbredelse. I tillegg til patogen-avledet ligander, TLRs er i stand til å binde endogene molekyler, kjent som skade-forbundet molekylære mønstre (demper), som er utgitt under celle nekrose og aktivere inflammatoriske signalering trasé resulterer i produksjon av proinflammatoriske cytokiner4. Vi og andre har foreslått at skaden pådratt under bløtvev biomaterialet implantation prosedyrer Release demper, som deretter adsorbere å biomaterialet overflater i tillegg til Blodproteiner og modulere påfølgende celle-materiale interaksjoner5,6. Når makrofager samhandler med adsorberes protein lag på et implantat, deres overflate TLRs kan gjenkjenne adsorberes demper og aktivere proinflammatoriske signalering kaskader, fører til NF-KB og AP-1 transkripsjon faktor aktivering og produksjon av proinflammatoriske cytokiner. Vi har tidligere vist at murine makrofager har signifikant økt NF-KB/AP-1 aktivitet og tumor nekrose faktor α (TNF-α, proinflammatoriske cytokin) sekresjon som svar på fuktig inneholder adsorberes protein lag på en rekke polymer overflater i forhold til overflater med adsorberes serum eller plasma bare (dvs. ingen demper stede), og at dette svaret er i stor grad formidlet av TLR2, mens TLR4 spiller en mindre rolle5.
NF-KB/Ap-1 reporter macrophage cellelinjen (tabell med materialer) som brukes i denne protokollen er en praktisk metode for å måle relativ NF-KB og Ap-1 aktivitet i makrofager5,7,8. I kombinasjon med TLR Pathway hemmere, denne cellen linjen er et nyttig verktøy for å undersøke TLR aktivisering og dens rolle i betennelse som svar på en rekke stimuli5,7,8. Reporteren celler er en modifisert mus macrophage-lignende cellelinje som kan stabilt produsere utskilles embryonale alkalisk fosfatase (SEAP) upon NF-KB og AP-1 transkripsjon faktor aktivering9. Den fargemetrisk enzymatisk alkalisk fosfatase analysen (tabell av materialer) kan deretter brukes til å kvantifisere RELATIVE mengder SEAP uttrykk som et indirekte mål på NF-KB/Ap-1 aktivitet. Som NF-KB og AP-1 er nedstrøms for mange celle signalering trasé, nøytralisere antistoffer og hemmere rettet mot bestemte TLRs (f. eks TLR2) eller TLR adapter molekyler (f. eks MyD88) kan brukes til å verifisere rollen til en bestemt vei. Metodikken som er beskrevet i denne artikkelen gir en enkel og rask tilnærming for å vurdere bidraget av TLR signalering i murine macrophage svar på en rekke polymer overflater med adsorberes protein lag som inneholder både Blodproteiner og demper som en in vitro modell av implantert Biomaterials.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. Media og reagens forberedelse
- Klargjør Fibroblast medium. Kombiner 450 mL av Dulbecco ' s modifiserte Eagle medium (DMEM), 50 mL av fosterets storfe serum (FBS), og 5 mL penicillin/Streptomycin. Oppbevares ved 4 ° c i opptil 3 måneder.
- Forbered reporter macrophage vekst medier i 50 mL alikvoter. Kombiner 45 mL av DMEM, 5 mL FBS, 5 μg/mL mycoplasma eliminerings reagens (tabell over materialer) og 200 μg/ml phleomycin D1 (tabell over materialer). Oppbevares ved 4 ° c i opptil 3 måneder.
- Forbered reporter macrophage analysen Media i 50 mL alikvoter. Kombiner 45 mL DMEM, 5 mL varme deaktivert FBS (HI-FBS), 5 μg/mL mycoplasma eliminerings reagens og 200 μg/mL phleomycin D1. Oppbevares ved 4 ° c i opptil 3 måneder.
2. coating Cell kultur overflater med Poly (metyl akrylat)
- Oppløsning Poly (metyl akrylat) (PMMA) i kloroform ved 20 mg/mL (f. eks, 100 mg PMMA i 5 mL kloroform) i et 20 mL glass scintillation hetteglass. Plasser en magnetisk rør i hetteglasset og la det røres i minst 2 timer, til alle faste stoffer oppløses.
Forsiktig: Kloroform er skadelig ved innånding. Sørg for å bruke løsemiddel i en avtrekksvifte mens du bruker PVA-hansker. - Pipetter 400 μL av PMMA-løsning på midten av et Borosilikatglass glas objekt lysbilde i en spin-elektrostatisk, og snurr ved 3000 RPM i 2 min. Forbered antall lysbilder som kreves for analysen, i tillegg til 3 − 5 ekstra for vann kontakt vinkel måling. Lagre lysbilder i en ren boks (sprayet og tørkes med 70% etanol) for fremtidig bruk.
Merk: Spin coating brukes ofte til å sette inn et tynt, jevnt belegg på en flat overflate. En spin elektrostatisk roterer et substrat ved høye hastigheter, ved hjelp av sentrifugalkraften til å spre belegg løsningen over overflaten.- Mål vann kontakt vinkel på to tilfeldige posisjoner på overflaten av ekstra belagt lysbilder (dvs. ikke lysbildene som brukes for cellekultur) med en goniometer for å sikre glassoverflaten var helt belagt med polymer.
Merk: Kun vann av høyeste renhet (f.eks. glass trippel destillert) skal brukes til måling av vann kontakt vinkel.
- Mål vann kontakt vinkel på to tilfeldige posisjoner på overflaten av ekstra belagt lysbilder (dvs. ikke lysbildene som brukes for cellekultur) med en goniometer for å sikre glassoverflaten var helt belagt med polymer.
- I et biologisk sikkerhetskabinett (BSC) feste 8-kammer klebrig brønner til PMMA belagt-lysbilder ved hjelp av steril tang og følgende aseptisk teknikk. Trykk godt på toppen av de klebrige brønnene for å sikre at de er sterkt festet. Ruge lysbildene med vedlagte klebrig-brønner ved 37 ° c over natten for å sikre forseglingen.
- Test forseglingen på de klebrige brønnene ved å tilsette 200 μL av cellekultur grade (endotoksin) vann til hver brønn. Ruge ved romtemperatur (RT) i 60 minutter og unngå lekkasje før du fortsetter. Aspirer vannet, være forsiktig med å forstyrre PMMA belegg.
- Utfør endotoksin vann vasker ved å tilsette 300 μL av endotoksin vann til hver brønn og incubating i 1 time (tre ganger), 12 timer og 24 timer før bruk for å fjerne gjenværende løsemiddel.
- Test endotoksin konsentrasjon av lysbildene som skal brukes for cellekultur. Ruge 200 μL av endotoksin-fritt reagens vann (tabell av materialer) i ett brønn av hvert lysbilde for 1 h. mål endotoksin konsentrasjon i ekstraktet ved hjelp av et ende punkts kromogen endotoksin-analysen (tabell med materialer).
Merk: Følgende protokoll er spesifikk for det endotoksin analyse settet som er oppført i materialfortegnelsen. - Bruk bare vann og Forbruksartikler (f.eks. pipette tips, mikrosentrifugen rør og brønn plater) som er sertifisert pyrogene (dvs. endotoksin) for dette arbeidet. Dessuten bør alle glass som brukes i utarbeidelsen av polymer-belagte overflater være depyrogenated ved hjelp av tørr varme sterilisering (250 ° c i 30 min) før bruk10. Måle endotoksin i ekstrakt løsningen, som beskrevet her, kan resultere i en undervurdering av endotoksin på materialet overflaten11,12. Følgelig er det anbefalt at ved utvikling av en polymer belegg protokoll, utføre endotoksin analysen reaksjon (dvs. trinn 2.5.4 − 2.5.6 for testprøver [reagens vann] eller spiker kontroller) direkte innenfor brønner som inneholder belagt prøven for å sikre at ingen kilder til endotoksin er utilsiktet innføres i systemet under belegget prosessen.
- Bring alle testprøver (dvs. ekstrakter) og endotoksin analysen reagenser til RT. Rekonstituer kromogen reagens i analysen buffer og endotoksin standard i reagens vann, tillate å oppløse i 5 min og forsiktig virvel før bruk. Dekk alle flasker med para fin film når den ikke er i bruk.
- Opprett en 5 − 8-punkts standard fortynning kurve av endotoksin standard som spenner fra den nedre til den øvre grensen av analysen ved å utføre en seriell fortynning av endotoksin standard i reagens vann.
- For å kontrollere for forbedring eller hemming av endotoksin analysen i testprøver, utarbeide en positiv kontroll (også kalt en spiker kontroll eller piggete utvalg) ved å fortynne en kjent mengde endotoksin i ubrukt test sample løsning.
Merk: Konsentrasjonen av den positive kontrollen bør være den samme konsentrasjonen som en standard i midten av standardkurven. Hvis den gjenopprettede mengden av endotoksin Spike (dvs. konsentrasjonen av den positive kontrollen minus konsentrasjonen av unspiked prøven) er innen 50 − 200% av den nominelle konsentrasjonen av endotoksin pigg, kan utvinnings løsningen anses å ikke signifikant forstyrre analysen. - Tilsett 50 μL av standarder, prøver eller Spike kontroller til hver brønn av en 96-brønn plate i duplikat eller tre eksemplarer. Bruk reagens vann som en negativ kontroll.
- Tilsett 50 μL av kromogen reagens til enhver brønn. Legg til reagens raskt på alle brønner. Bruk en tidtaker til å registrere hvor mye tid det tar å legge til reagens i alle brønner. Dekkplaten med en selvklebende tetning og ruge ved 37 ° c (inkubasjonstid er mye avhengig og oppgitt på analysesertifikat inkludert i kromogen reagenssett). Alternativt, sjekk på tallerkenen hver 15 min under inkubasjons til fargeendring er observert i alle standard brønner.
- Etter inkubasjons, tilsett 25 μL av 50% eddiksyre til hver brønn (endelig konsentrasjon på 10% eddiksyre per brønn) for å stoppe reaksjonen. Tilsett eddiksyre i samme rekkefølge som kromogen reagens ble tilsatt. Les absorbansen av platen ved hjelp av en plate leser ved 405 NM. Aspirer væske og kast plate.
Merk: Eddiksyre tillegg bør ta like lang tid å legge til hver brønn som kromogen reagens tok (± 30 s).
- Ultrafiolett (UV) sterilisere lysbildene i 30 minutter før cellekultur eksperimenter.
3. coating Cell kultur overflater med Polydimethylsiloxan
- Bland Polydimethylsiloxan (PDMS) elastomer i et 10:1 vekt forhold (Base: herding agent). I et biologisk sikkerhetskabinett kan du Pipetter ca. 10 mL Polydimethylsiloxan base i et sterilt rør. Veie røret og langsomt legge herding agent til 10% er lagt til.
Forsiktig: Bruk PDMS-reagenser i et godt ventilert område, og unngå øyekontakt ved å bruke vernebriller. - Bland elastomer grundig ved å røre med en steril, serologisk pipette spiss og ved å pipettering opp og ned. Tilsett ca 200 μL av løsningen på hver brønn av en 48-brønn plate. Vipp brønn platen langsomt for å sikre fullstendig dekning av brønner med elastomer løsning.
- Plasser brønn platen med elastomer i en vakuum ovn satt til 50 cmHg, 40 ° c. Fjern lokket og dekk med en enkelt-lags tørk for å hindre at andre rusk faller inn i brønnene. La det ruge i minst 48 h.
- Bekreft at brønnene er helt belagt via visuell inspeksjon. Sørg for at elastomer er fullstendig kurert ved å forsiktig prodding med en steril pipette før du tar den ut.
- Tilsett 300 μL av 70% etanol (laget med absolutt etanol og endotoksin vann) og ruge ved RT for 1 h. Fjern etanol og utfør endotoksin vann vasker ved å tilsette 300 μL av endotoksin vann til hver brønn og incubating i 1 time (tre ganger), 12 timer , og 24 timer før bruk for å fjerne eventuelle gjenværende løsemiddel.
- Ruge 200 μL av endotoksin vann i tre brønner på hver plate for 1 h. mål endotoksin konsentrasjon av vann ekstrakter ved hjelp av en ende punkts kromogen endotoksin-analyse (trinn 2.5.1 − 2.5.6).
4. coating Cell kultur overflater med Fluoriserte Poly (tetrafluoroethylene)
- Lag en 1 mg/mL oppløsning av fluoriserte Poly (tetrafluoroethylene) (fPTFE) (f. eks, tilsett 10 mg fPTFE til 10 mL fluoriserte løsemiddel [tabell over materialer]) i et 20 ml glass scintillation hetteglasset. Plasser et magnetisk rør i hetteglasset og la det røre i minst 24 timer, til alle faste stoffer oppløses.
- Tilsett ca 150 μL av polymer løsningen til hver brønn av en polystyren 48-brønn plate (dvs. ikke vevs kultur behandlet). Vipp brønn platen langsomt for å sikre fullstendig dekning av alle brønner med polymer løsning. Skift lokk.
- For å sikre effektiv fPTFE-belegg av brønner, bør glass coverslips være belagt i fPTFE og brukes til vann kontakt vinkel måling (trinn 4.3.1). Plasser coverslips inne i brønnene på en 24-brønn plate. Tilsett ca 400 μL av polymer løsningen til hver brønn som inneholder en dekkglass. Skyv coverslips ned ved hjelp av steril tang, slik at de er helt dekket i polymer løsning, og dekker brønnen plate med et lokk.
- Plasser brønn platen med polymer oppløsning og/eller coverslips i en vakuum ovn satt til 50 cmHg, 40 ° c. Fjern lokket og dekk med en enkelt-lags tørk for å hindre at andre rusk faller inn i brønnene. La det ruge i minst 48 h.
- Mål vann kontakt vinkelen på fPTFE-belagt coverslips med en goniometer for å sikre effektiv belegg.
Merk: Kun vann av høyeste renhet (f.eks. glass trippel destillert) skal brukes til måling av vann kontakt vinkel.
- Mål vann kontakt vinkelen på fPTFE-belagt coverslips med en goniometer for å sikre effektiv belegg.
- Tilsett 300 μL av 70% etanol (laget med absolutt etanol og endotoksin vann) og ruge ved RT for 1 h. Fjern etanol og utfør endotoksin vann vasker ved å tilsette 300 μL av endotoksin vann til hver brønn og incubating i 1 time (tre ganger), 12 timer , og 24 timer før bruk for å fjerne eventuelle gjenværende løsemiddel.
- Ruge 200 μL av endotoksin vann i tre brønner på hver plate for 1 h. mål endotoksin konsentrasjon av vann ekstrakter ved hjelp av en ende punkts kromogen endotoksin-analyse (trinn 2.5.1 − 2.5.6).
- UV sterilisere brønn platene i 30 minutter før cellekultur eksperimenter.
5. Making Lysat fra 3T3 celler
- Grow 3T3 celler i flere T150 flasker til 70% samløpet. Å løsne celler, aspirer medier, vaske overflaten med 5 mL PBS, og aspirer PBS. Tilsett 5 mL animalsk opprinnelse-gratis, rekombinant Cell dissosiasjon enzym (tabell av materialer) og ruge ved 37 ° c for 3 − 5 min.
- Løsne cellene ved å vippe flasken forsiktig frem og tilbake. Tilsett 5 mL PBS å nøytralisere rekombinant enzymet som brukes for celle dissosiasjon. Overfør de frittliggende cellene fra flaskene til et sentrifugerør og bland dem via pipettering. Utfør en live celle telling ved hjelp av en hemocytometer og celle levedyktighet fargestoff.
Merk: En celle dissosiasjon enzym som kan nøytralisert gjennom fortynning i PBS ble valgt for å unngå innføring av serum-baserte proteiner i lysat forberedelse. Hvis Trypsin brukes til å distansere celler, bør det være nøytralisert med en serum-inneholdende løsning, og en ekstra PBS vask bør utføres for å redusere mengden av serumproteiner overført til lysat forberedelse. - Sentrifuger cellene ved 200 x g i 5 min. aspirer supernatanten og resuspend celler i original volum (dvs. 10 ml x antall flasker) av PBS å vaske av eventuelle gjenværende medier. Gjenta.
- Sentrifuger cellene igjen ved 200 x g i 5 min og aspirer supernatanten. Legg volumet av PBS nødvendig for å oppnå en endelig celle konsentrasjon på 1 x 106 celler/ml. Plasser celle løsningen i en-80 ° c fryser til prøven er helt frosset (minst 2 timer).
- Tin celle løsning i et vannbad på 37 ° c. Når den er fullstendig tint, plasser løsningen tilbake i-80 ° c fryseren til helt frosset. Gjenta for totalt 3 fryse-tine sykluser.
- Utfør en mikro bicinchoninic syre (BCA) analysen på cellen lysat på en rekke fortynninger (f. eks, 1/100, 1/200, 1/500, 1/1000) for å bestemme proteinkonsentrasjon. Fortynne celle lysat til en proteinkonsentrasjon på 468,75 μg/mL, alikvot og oppbevares ved-80 ° c for fremtidig bruk.
Merk: Endelig proteinkonsentrasjon i en 48-brønn er 125 μg/cm2 (basert på overflatearealet på en brønn, 0,75 cm2). - Utfør en Western Blot for å vurdere tilstedeværelsen av demper i lysat (for eksempel varme sjokk protein 60 [HSP60], høy mobilitet gruppeboks 1 [HMGB1]) ved lasting 40 − 60 mikrogram lysat protein i lasting buffer på en 1,5 mm tykk 10% polyakrylamid gel og følger standard Western Blot Prosedyrer.
6. vurdere effekten av adsorberes protein lag og toll-lignende reseptorer på NF-KB aktivitet av makrofager
Merk: For en skjematisk av eksperimentell arbeidsflyt og plate layout, se figur 1a og supplerende figur 1, henholdsvis.
- Grow reporter makrofager i en passende størrelse kolbe til 70% samløpet. Aspirer Media, vaske overflate med PBS, og aspirer PBS. Tilsett rekombinant Cell dissosiasjon enzym og ruge ved 37 ° c i 8 min.
- Løsne celler ved fast trykke på sidene av flasken. Deaktiver rekombinant Cell dissosiasjon enzym ved å legge til et lik volum av vekst medier (som inneholder 10% FBS). Utfør en live celle telling ved hjelp av en hemocytometer og celle levedyktighet fargestoff.
Merk: Forventet levedyktighet for reporteren makrofager etter en 8 min inkubasjons i cellen dissosiasjon enzymet er 90%. - Sentrifuger celler ved 200 x g for 5 min. aspirer supernatanten og resuspend i original volumet av PBS å vaske celler. Sentrifuger igjen og resuspend celler på 7,3 x 105 celler/ml i analysen Media (som inneholder varme deaktivert FBS).
- Separat celle suspensjon i 3 forskjellige rør: TLR4 inhibitor, anti-TLR2, og ubehandlet. Ruge celler med 1 μg/mL TLR4-hemmer for 60 min ved RT eller med 50 μg/mL anti-TLR2 i 30 minutter ved RT.
- Tilsett 200 μL av lysat, 10% FBS, 10% kommersiell mus plasma (tabell av materialer), eller en blanding av protein løsninger til en 48-brønn plate (eller tilsvarende) og tillate protein å adsorbere ved 37 ° c for den ønskede tiden (dvs. 30 min, 60 min, eller 24 h). Aspirer protein løsninger fra brønner, ved hjelp av en frisk, Pasteur-pipette for hver protein løsning, og vask overflater med 250 μL av PBS i 5 min. aspirer PBS. Gjenta for totalt 3 vasker.
Merk: Dette trinnet må kanskje startes tidligere i protokollen, avhengig av ønsket absorpsjons tid. Juster protokollen i henhold til dette. - Etter inkubasjonsperiode med TLR4-hemmere eller anti-TLR2, Pipetter celler til resuspend. Tilsett 200 μL av celle løsning i hver brønn.
- For TLR2 positiv kontroll tilstand, tilsett Pam3CSK4 til en endelig konsentrasjon på 150 ng/mL. For TLR4 positiv kontroll tilstand, Legg lipopolysakkarid (LPS) til en endelig konsentrasjon av 1,5 μg/mL. Ruge celler ved 37 ° c i 20 timer.
- Eksempel på 20 μL av supernatanten fra hver brønn og plate i duplikat i en 96 brønn plate. Inkluder tre brønner på 20 μL-analyse medier som en bakgrunns kontroll. Tilsett 200 μL av SEAP reporter analysen reagens til hver brønn. Dekkplaten med en selvklebende forsegling og ruge for 2,5 h ved 37 ° c.
Merk: Inkubasjonstiden kan variere avhengig av eksperimentelle forhold, og bør optimaliseres for en sterk forskjell i absorbansen mellom positive og negative kontroll brønner.- Overfør resten av supernatanten til en 1,5 mL slange (per brønn). Sentrifuger på 1 000 x g for 10 min til pellets noe rusk. Overfør supernatanten til en ny 1,5 mL tube og oppbevares ved-80 ° c. Analyser supernatanten for tilstedeværelsen av proinflammatoriske cytokiner (f.eks. TNF-α, interleukin 6) via enzym koblet immunosorbentanalyse-analyse (ELISA).
- Fjern lim plate forseglingen. Les absorbansen av platen ved hjelp av en plate leser ved 635 NM. Aspirer væske og kast plate.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Rengjøringsmetoder for polymer belagte overflater ble testet for å sikre at det ikke var noen forstyrrelse av belegget, noe som ville bli sett på som en endring i vann kontakt vinkelen til en dekkglass i glass (figur 2). Soaking PMMA-belagt mikroskop lysbilder i 70% etanol for 1 h ble funnet å fjerne PMMA belegg (figur 2, venstre panel), sannsynligvis på grunn av LØSELIGHET av PMMA i 80 WT% etanol13, derfor PMMA-belagte overflater ble rengjort med 30 min av UV sterilisering alene. Konsentrasjonen av PMMA for belegget var optimalisert tidligere5. A 1 t 70% etanol suge ble brukt til å rengjøre PDMS, og UV sterilisering ble neglisjert siden UV-lys kan forårsake kjede scission og påvirke overflaten fukting egenskaper av PDMS14. Begge 70% etanol suge og UV sterilisering ikke påvirke vann kontakt vinkelen fPTFE-belagt coverslips (figur 2, høyre panel), derfor de to metodene, på rad, ble brukt til å rense fPTFE belegg. Metoden for fPTFE belegg ble tidligere beskrevet av Grainger gruppe15.
En Western Blot ble utført på 3T3 lysat for å sikre at FUKTIGE arter var til stede i den komplekse molekylære blandingen. Resultatene viste at både HMGB1 og HSP60, to godt dokumentert demper16,17, var til stede i lysat (figur 1B). Absorpsjon av TLR ligander fra lysat på polymer overflater ble bekreftet av dyrking reporter makrofager (ubehandlet, TLR2 nøytralisert, eller TLR4 hemmet) for 20 h på protein-adsorberes polymer overflater (dvs. vev kultur-behandlet polystyren [TCPS], PMMA, PDMS, fPTFE), og deretter indirekte vurdere NF-кB/AP-1 aktivitet basert på SEAP produksjon ved hjelp av en enzymatisk analysen (figur 1C og Figur 3). Videre hadde reporteren makrofager signifikant økt NF-кB/AP-1 aktivitet på adsorberes lysat sammenlignet med adsorberes FBS eller plasma og ingen pre-adsorberes protein (Media) (Figur 4). TLR ligander syntetisk triacylated lipopeptide (Pam3CSK4, TLR2 ligand) og lipopolysakkarid (LPS, TLR4 ligand) ble inkludert som positive kontroller for å bekrefte antistoff eller hemmer og analysen ble fungerer som den skal. TLR2 nøytralisering hadde en merkbart sterkere reduksjon i NF-кB/AP-1 respons av reporter makrofager å adsorberes lysat sammenlignet med TLR4 hemming. I tillegg, små mengder lysat fortynnet i serum (basert på total protein) indusert betydelig økt NF-кB/AP-1 respons sammenlignet med serum alene, med lavest effektiv fortynning avhengig av polymer overflaten (figur 5). Disse resultatene viser styrken på adsorberes lysat-avledede molekyler på inducing TLR-avhengige NF-кB/AP-1 aktivitet i reporter makrofager på en rekke polymer overflater.

Figur 1: metoder og resultater for alkalisk fosfatase analysen av NF-кB/Ap-1 reporter makrofager på TCPS, PMMA, PDMS og fPTFE. (A) diagram av arbeidsflyten for reporteren macrophage alkalisk fosfatase analysen. (B) Western Blot av lysat bekrefter tilstedeværelsen av damp arter HMGB1 og HSP60, med β-utgangen som lasting kontroll. (C) NF-КB/Ap-1 aktivitet (representert ved absorbansen) av reporter makrofager kultivert på Media (negativ kontroll), 10% FBS, lysat, og PAM3CSK4 (TLR2 ligand, positiv kontroll) for 20 h. data viser resultatene av ett eksperiment og er representativt for resultater fra minst 2 separate eksperimenter, vist som gjennomsnittet ± standardavvik (SD). Hvert eksperiment brukes n = 3 separate brønner per tilstand, og hver brønn ble belagt i duplikat for enzymatisk analysen. Analysert ved hjelp av enveis ANOVA og Tukey post-hoc-test. p < 0,001. Dette tallet er tilpasset med tillatelse fra McKiel og Fitzpatrick5. Copyright 2018 American Chemical Society. Vennligst klikk her for å se en større versjon av dette tallet.

Figur 2: optimalisering av rengjøringsmetoder for PMMA-og fPTFE-belagte overflater, vurdert ved hjelp av vann kontakt vinkel (WCA). Målingene ble tatt på 2 separate punkter av minst 3 coverslips. Data vises som gjennomsnittet ± SD. analysert ved hjelp av enveis ANOVA og Tukey post-hoc-test. * p < 0,05. Dette tallet er tilpasset med tillatelse fra McKiel og Fitzpatrick5. Copyright 2018 American Chemical Society. Vennligst klikk her for å se en større versjon av dette tallet.

Figur 3: TLR-MEDIERT NF-кB/Ap-1 aktivitet (representert ved absorbansen) av reporter makrofager kultivert på 10% FBS (kontroll), lysat, og positiv kontroll for 20 t. (A) påvirkning av TLR2 nøytralisering på reporter macrophage svar på adsorberes lysat. Positiv kontroll er Pam (Pam3CSK4, TLR2 ligand). (B) påvirkning av TLR4 hemming på reporter macrophage respons på adsorberes lysat. Positiv kontroll er LPS (TLR4 ligand). Data viser resultatene av ett eksperiment og er representativt for resultater fra minst 2 separate eksperimenter, vist som gjennomsnittet ± SD. Hvert eksperiment brukes n = 3 separate brønner per tilstand, og hver brønn ble belagt i duplikat for enzymatisk analysen. Analysert ved hjelp av enveis ANOVA og Tukey post-hoc-test. * * p < 0,01, * * * p < 0,001. Dette tallet er tilpasset med tillatelse fra McKiel og Fitzpatrick5. Copyright 2018 American Chemical Society. Vennligst klikk her for å se en større versjon av dette tallet.

Figur 4: NF-кB/Ap-1 aktivitet (representert ved absorbansen) av reporter makrofager kultivert på Media (negativ kontroll), 30 min og 24 h adsorberes protein lag, og Pam3CSK4 (positiv kontroll) på TCPS for 20 h. Data kombineres fra 3 separate eksperimenter og vises som gjennomsnittet ± SD. Hvert eksperiment brukes n = 3 separate brønner per tilstand, og hver brønn ble belagt i duplikat for enzymatisk analysen (dvs. n = 9 ikke-uavhengige cellekultur brønner og n = 18 ikke-uavhengige enzymatisk analysen brønner). Analysert ved hjelp av enveis ANOVA og Tukey post-hoc-test. p < 0,001. Vennligst klikk her for å se en større versjon av dette tallet.
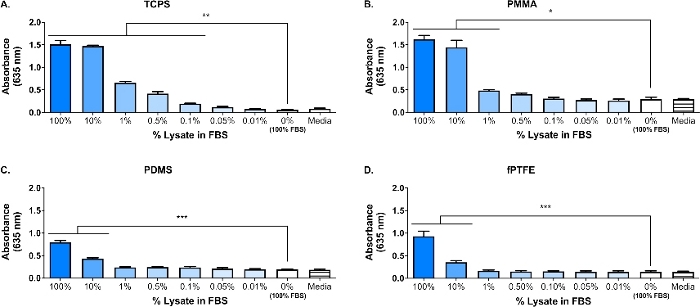
Figur 5: reporter MACROPHAGE NF-кB/Ap-1 aktivitet (representert ved absorbansen) etter 20 timer som svar på fortynninger av lysat i FBS (totalt protein = 280 μg/brønn) adsorberes til polymer overflater i 30 min. (A) TCPS. (B) PMMA. (C) PDMS. (D) fPTFE. Data viser resultatene av ett eksperiment og er representativt for resultater fra minst 2 separate eksperimenter, vist som gjennomsnittet ± SD. Hvert eksperiment brukes n = 3 separate brønner per tilstand, og hver brønn ble belagt i duplikat for enzymatisk analysen. Analysert ved hjelp av enveis ANOVA og Tukey post-hoc-test. * p < 0,05, * * p < 0,01, * * * p < 0,001. Dette tallet er tilpasset med tillatelse fra McKiel og Fitzpatrick5. Copyright 2018 American Chemical Society. Vennligst klikk her for å se en større versjon av dette tallet.

Supplerende figur 1: eksempel oppsett som brukes for NF-кB/Ap-1 reporter macrophage cellekultur analysen i 8-kammer og 48-brønn plateformater. Vennligst klikk her for å se en større versjon av dette tallet.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Hovedfokuset til laboratoriet vårt er å svare på solide biomaterialet bløt vevs implantater, og spesielt hvordan den cellulære skaden som oppstår under implantat prosessen påvirker verts responsen. Arbeidet som presenteres her beskriver foreløpige eksperimenter ved hjelp av en reporter macrophage cellelinje og in vitro-genererte DAMP-inneholdende cellulære lysat, for å undersøke påvirkningen av molekyler som ble utgitt under cellulær skade (dvs. fra implantat operasjonen) på macrophage responser til Biomaterials. Fibroblast celle lysat ble brukt til å modellere cellulære skader og frigjøring av demper på grunn av biomaterialet plassering. Fibroblaster ble valgt til å skape lysat på grunn av utbredelsen av fibroblaster i bløtvev, så vel som deres evne til å skille ut en rekke ekstracellulære matrise (ECM) proteiner, inkludert fibronektin18. Freeze-Tin sykling ble valgt som metode for lyse å produsere både intracellulære og ECM-avledet demper, ligner på hva som ville være til stede i implantatet miljøet. Protease hemmere ble ikke brukt til å lage denne lysat. Mens ukontrollert cellelyse som fryse-tine sykling kan føre til frigjøring av proteaser som kan svekke demper, disse enzymene vil også trolig være tilstede i biomaterialet implantat miljø når cellene er skadet under implantation prosedyren. Tilstedeværelsen av demper i den komplekse molekylære blandingen av lysat ble bekreftet av Western Blot (figur 1B; HMGB1 og HSP60) og SEAP reporter analysen (figur 1C; NF-кB/AP-1-aktivitet som svar på adsorberes lysat). Vi har også utført analyser der lysat ble fortynnet i FBS basert på total proteinkonsentrasjon og adsorberes på cellekultur overflater (figur 5) for å bedre gjenspeile kompleksiteten i implantat miljøet, siden det vil inneholde en overflod av Blodproteiner så vel som demper6. Reporter macrophage NF-кB/AP-1 aktivitet forble betydelig økt på adsorberes lag fra lysat fortynnet i FBS, og den laveste fortynning for å oppnå betydelig aktivering var overflaten avhengig, alt fra 0,1% (TCPS) til 10% (PDMS og fPTFE).
Polymerer PMMA, PDMS og PTFE ble valgt for dette arbeidet fordi de er nondegradable og har blitt brukt mye i litteraturen for å vurdere absorpsjon av protein og macrophage respons på Biomaterials19,20,21,22,23,24,25. TCPS ble også brukt til sammenligning siden det er et vanlig substrat brukes for in vitro macrophage og TLR signalering arbeid21,26,27,28. Materialene som brukes i vårt arbeid er representative eksempler på ikke-nedbrytbart, solid Biomaterials. Imidlertid kan mange andre materialer brukes med denne modellen, forutsatt at materialet kan være belagt på cellekultur plater eller mikroskop lysbilder og riktig dekontaminert. NF-KB/AP-1 reporter macrophage cellelinjen ble valgt for denne in vitro-modellen fordi den muliggjør rask, indirekte måling av NF-KB/AP-1-aktivitet gjennom NF-KB/AP-1 induserbart uttrykk for SEAP. NF-KB/AP-1 reporter makrofager krever bruk av phleomycin D1 i kultur Media som en selektiv antibiotika for å sikre at bare celler med NF-кB/AP-1 induserbart SEAP genet er til stede29. For alkalisk fosfatase analysen, er det avgjørende å bruke HI-FBS i cellen kultur Media for å unngå potensielle falske positive resultater generert av alkaliske phosphatases stede i serum. Vår forskning hittil tyder på at FBS-adsorberes overflater ikke genererer et synlig falskt positivt resultat, sannsynligvis fordi serum molekylene er sterkt adsorberes til kulturen overflaten og er ikke sluppet inn i supernatanten. Kulturen tidspunkt for reporteren makrofager (20 h), analysen inkubasjons timepoint (2,5 h), og absorbansen lesing bølgelengde (635 NM) for alkalisk fosfatase analysen ble optimalisert med dette systemet for å sikre robust og reproduserbar målinger for alle forhold.
En innledende absorpsjons timepoint på 30 minutter ble valgt for dette arbeidet på grunn av den vanlige bruken i protein absorpsjons litteraturen (figur 1C)30,31,32,33,34. Vi har imidlertid også utforsket lengre absorpsjons tider (dvs. 60 min og 24 h, Figur 4) for å bedre representere det adsorberes protein laget som makrofager ville samhandle med in vivo, som sannsynligvis vil forekomme 4 − 24 timer etter implantation1. Det har blitt postulert at flertallet av protein absorpsjon og utveksling skjer i de første 60 min av eksponering til en overflate26,35,36, derfor en 60 min absorpsjons tid kan være en mer relevant timepoint. Vi har også flyttet fra å bruke FBS som en negativ kontroll for tilstedeværelsen av demper i adsorberes protein laget til kommersiell mus plasma. Begrunnelsen for å bruke plasma i stedet for serum er at plasmaproteiner er kjent for å spille viktige roller i protein absorpsjon og macrophage respons1, og at plasma gir en bedre representasjon av proteiner i såret miljøet. Plasma som brukes i protein absorpsjons eksperimenter, fremstilles vanligvis som en 1 − 10% fortynning26,36,37, som motiverte vår bruk av 10% plasma. Humant plasma brukes vanligvis26,36, da det er lettere å få i store mengder og mer klinisk relevant, sammenlignet med mus plasma. Men, valgte vi å bruke kommersiell mus plasma for i denne modellen for å holde arter av protein løsninger i samsvar med at av reporteren celler.
Bruken av reporteren macrophage cellelinje innført noen begrensninger i studien. For det første, benytter en murine Leukemic macrophage cellen Line har iboende begrensninger, idet det fenotype og opptreden kanskje variere fra primære macrophage kulturen. Selv om denne begrensningen vil bli behandlet i fremtidig arbeid ved hjelp av primære makrofager, ble foreldrenes macrophage cellelinje vist å tett etterligne mus benmarg-avledet makrofager i form av deres celle overflate reseptorer og respons på mikrobiell ligander for TLRs 2, 3 og 438. Videre, NF-KB/AP-1 reporter makrofager gitt lignende resultater som svar på HMGB1 og LPS stimulering i forhold til bukhulen primære murine makrofager39. Det bør bemerkes at NF-KB/AP-1 reporter makrofager, og deres foreldrenes belastning, ikke uttrykke TLR540. Forskere har vist at HMGB1 var i stand til å aktivere NF-KB transkripsjon faktorer via TLR5 signalering trasé i HEK-293 celler stabilt transfekterte med menneskelig TLR541. Derfor er bidraget fra HMGB1-TLR5 signaliserer til samlet NF-KB aktivitet på lysat overflater ble neglisjert i denne modellen. I tillegg reporter makrofager og foreldrenes belastning ikke uttrykke ASC adapter protein, og følgelig ikke danne de fleste typer inflammasomes og kan ikke behandle inaktive IL-1β eller inaktiv IL-18 til sine eldre former42. Derfor har modellen vi har brukt ikke står for bidraget av ASC-avhengige inflammasome aktivitet og påfølgende autocrine IL-1β og IL-18 signalering i macrophage svar på lysat-adsorberes overflater. Følgelig, denne analysen er ment som en foreløpig undersøkelse av TLR-avhengige NF-KB aktivisering, og påfølgende forskning bruker primære makrofager anbefales å gi en mer komplett og representativ forståelse av macrophage aktivisering og fenotype på materielle overflater av interesse.
Alkalisk fosfatase analysen indirekte måler NF-кB/AP-1 aktivitet av reporteren makrofager. Men det er mange signalering veier enn TLRs som involverer NF-кB/AP-1 (f. eks interleukin-1 reseptor [IL-1R]43 og tumor nekrose faktor RESEPTOR [TNFR]44). Derfor var det nødvendig å vurdere bidraget av TLR2 og TLR4 signalering i den økte NF-кB/AP-1 respons på lysat-adsorberes overflater ved hjelp av hemming analyser (Figur 3). Begrunnelsen for å velge disse to overflaten TLRs var at minst 23 demper som har vist å signalisere gjennom TLR2 og TLR445, inkludert godt karakterisert HMGB1, og begge reseptorer er uttrykt på celleoverflaten og kan samhandle direkte med biomaterialet overflate6. Den TLR2 og TLR4 hemming analysene viste at når TLR2 eller TLR4 signalering ble blokkert, NF-кB/AP-1 respons av reporteren makrofager til adsorberes lysat ble redusert, noe som indikerer at begge veier er involvert. Det var imidlertid en merkbart større reduksjon i NF-кB/AP-1 aktivitet når TLR2 signalering ble nøytralisert, noe som tyder på at TLR2 kan spille en primær rolle i responsen fra reporter makrofager til adsorberes lysat. Vi erkjenner at det kan være noe utenfor målet hemming med TLR signalering veien nøytralisere antistoffer og hemmere. En nøytralisere antistoff ble brukt til å hemme TLR2 veien siden det ikke var kommersielt tilgjengelig TLR2 inhibitor molekyler på tidspunktet for dette arbeidet.
Metodene som presenteres her bruker lysat, som en kompleks kilde til demper, og NF-KB/AP-1 reporter makrofager som en in vitro-modell for macrophage svar på demper og andre proteiner adsorberes til polymer Biomaterials (figur 1). Vi forventer vår protokoll kan brukes til å raskt analysere NF-KB/AP-1 svar og oppstrøms TLR signalering av reporter makrofager til en rekke materialer (inkludert nedbrytbart materialer, porøse stillaser eller hydrogeler) og adsorberes protein lag (Figur 3). Bruken av porøse materialer og hydrogeler vil imidlertid innføre kompleksitet i systemet, da det kan være utfordrende å skille mellom adsorberes molekyler og entrained molekyler. Vi forventer at denne protokollen kan enkelt tilpasses for å undersøke bidraget fra andre signalnettverk sti oppstrøms av NF-кB/AP-1 (f. eks C-type Lektiner reseptorer46 og nukleotid-binding oligomerisering domene (Nod)-lignende reseptorer47) med riktig hemmere. Videre, NF-KB/AP-1 respons av reporter makrofager kan sammenlignes mellom ulike materialer, forutsatt at svarene er normalisert til Baseline celle aktivitet (dvs. celler i Media på hver overflate uten pre-adsorberes protein) og alle materialer har undetectable endotoksin nivåer.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Forfatterne har ingenting å avsløre.
Acknowledgments
Forfatterne takknemlig erkjenner operasjonelle midler fra Canadian Institutes of Health Research Project (PTJ 162251), Queen ' s University Senatet Advisory Research Committee og infrastruktur støtte fra Canadian Foundation for Innovation John Evan ' s Leadership Fund (Project 34137) og departementet for forskning og innovasjon Ontario Research Fund (Project 34137). L.A.M. ble støttet av en Queen ' s University R. Samuel McLaughlin Fellowship, en Natural Sciences og engineering Research Council of Canada kanadiske Graduate Scholarship Master ' s Award og en Ontario Graduate Scholarship. Forfatterne vil gjerne takke Dr. Myron Szewczuk for sin sjenerøse gave NF-KB/AP-1 reporter macrophage cellelinje og DRS. Michael Blennerhassett og Sandra Lourenssen for bruk av deres gel Imaging system og plate leser.
Materials
| Name | Company | Catalog Number | Comments |
| Cell culture reagents | |||
| anti-mouse/human CD282 (TLR2) | Biolegend | 121802 | |
| CLI-095 (TLR4 inhibitor) | Invivogen | TLRL-CLI95 | |
| C57 complement plasma K2 EDTA 10ml, innovative grade US origin | InnovativeResearch | IGMSC57-K2 EDTA-Compl-10ml | Mouse plasma |
| Dulbecco's modified eagle medium (DMEM) | Sigma Aldrich | D6429-500ML | |
| Dulbecco's phosphate buffered saline (DPBS) | Fisher Scientific | 14190250 | No calcium, no magnesium |
| Fetal bovine serum (FBS), research grade | Wisent | 98150 | |
| LPS-EK | Invivogen | TLRL-EKLPS | Lipopolysaccharide from Escherichia coli K12 |
| NIH/3T3 fibroblasts | ATCC | CRL-1658 | |
| Pam3CSK4 | Invivogen | tlrl-pms | Synthetic triacylated lipopeptide - TLR1/2 ligand |
| Penicillin/streptomycin | Sigma Aldrich | P4333-100ML | |
| Plasmocin | Invivogen | ANT-MPP | Mycoplasma elimination reagent |
| RAW-Blue cells | Invivogen | raw-sp | NF-κB/AP-1 reporter macrophage cell line |
| Trypan blue solution, 0.4% | Fisher Scientific | 15250061 | |
| TrypLE express enzyme (1X) | Fisher Scientific | 12604021 | animal origin-free recombinant cell dissociation enzyme |
| Zeocin | Invivogen | ANT-ZN-1 | |
| Kits and assays | |||
| ELISA precoated plates, mouse IL-6 | Biolegend | B213022 | |
| ELISA precoated plates, mouse TNF-α | Biolegend | B220233 | |
| Endotoxin (Escherichia coli) - Control standard endotoxin (CSE) | Associates of Cape Cope Inc. | E0005-5 | Endotoxin for standard curve in chromogenic endotoxin assay |
| LAL water, 100 mL | Associates of Cape Cope Inc. | WP1001 | Used with chromogenic endotoxin assay |
| Micro BCA protein assay | Fisher Scientific | PI23235 | |
| Limulus amebocyte lysate (LAL) Pyrochrome endotoxin test kit | Associates of Cape Cope Inc. | C1500-5 | Chromogenic endotoxin assay reagent |
| QUANTI-Blue alkaline phosphatase detection medium | Invivogen | rep-qb2 | Alkaline phosphatase assay to indirectly measure NF-κB/AP-1 activity |
| Polymeric coating reagents | |||
| Chloroform, anhydrous | Sigma Aldrich | 288306-1L | |
| Ethyl alcohol anhydrous | Commercial Alcohols | P006EAAN | Sigma: Reagent alcohol, anhydrous, 676829-1L |
| Straight tapered fine tip forceps | Fisher Scientific | 16-100-113 | |
| Fluorinert FC-40 solvent | Sigma Aldrich | F9755-100ML | Fluorinated solvent for fPTFE |
| Cell culture grade water (endotoxin-free) | Fisher Scientific | SH30529LS | |
| Poly(methyl methacrylate) (PMMA) | Sigma Aldrich | 182230-25G | |
| Sylgard 184 elastomer kit | Fisher Scientific | 50822180 | |
| Teflon-AF (fPTFE) | Sigma Aldrich | 469610-1G | Poly[4,5-difluoro-2,2-bis(trifluoromethyl)-1,3-dioxole-co-tetrafluoroethylene] |
| Consumables | |||
| Adhesive plate seals | Fisher Scientific | AB-0580 | |
| Axygen microtubes, 1.5 mL | Fisher Scientific | 14-222-155 | |
| Borosilicate glass scintillation vials, with white polypropylene caps | Fisher Scientific | 03-337-4 | |
| Clear PS 48-well plate | Fisher Scientific | 08-772-52 | |
| Clear TCPS 96-well plate | Fisher Scientific | 08-772-2C | |
| Clear TCPS 48-well plate | Fisher Scientific | 08-772-1C | |
| Cover glasses, circles | Fisher Scientific | 12-545-81 | |
| Falcon tissue culture treated flasks, T25 | Fisher Scientific | 10-126-10 | |
| sticky-Slide 8 Well | Ibidi | 80828 | |
| Superfrost microscope slides | Fisher Scientific | 12-550-15 | |
| Tissue culture treated flasks, T150 | Fisher Scientific | 08-772-48 |
References
- Anderson, J. M., Rodriguez, A., Chang, D. T. Foreign body reaction to biomaterials. Seminars in Immunology. 20 (2), 86-100 (2008).
- Anderson, J. M., Miller, K. M.
- Collier, T. O., Anderson, J. M. Protein and surface effects on monocyte and macrophage adhesion, maturation, and survival. Journal of Biomedical Materials Research. 60 (3), 487-496 (2002).
- Bianchi, M. E. DAMPs, PAMPs and alarmins: all we need to know about danger. Journal of Leukocyte Biology. 81 (1), 1-5 (2007).
- McKiel, L. A., Fitzpatrick, L. E. Toll-like Receptor 2-Dependent NF-κB/AP-1 Activation by Damage-Associated Molecular Patterns Adsorbed on Polymeric Surfaces. ACS Biomaterials Science & Engineering. 4 (11), 3792-3801 (2018).
- Babensee, J. E. Interaction of dendritic cells with biomaterials. Seminars in Immunology. 20 (2), 101-108 (2008).
- Sintes, J., Romero, X., de Salort, J., Terhorst, C., Engel, P. Mouse CD84 is a pan-leukocyte cell-surface molecule that modulates LPS-induced cytokine secretion by macrophages. Journal of Leukocyte Biology. 88 (4), 687-697 (2010).
- Tom, J. K., Mancini, R. J., Esser-Kahn, A. P. Covalent modification of cell surfaces with TLR agonists improves and directs immune stimulation. Chemical Communications. 49 (83), 9618-9620 (2013).
- Abdulkhalek, S., et al. Neu1 sialidase and matrix metalloproteinase-9 cross-talk is essential for toll-like receptor activation and cellular signaling. Journal of Biological Chemistry. 286 (42), 36532-36549 (2011).
- Gorbet, M. B., Sefton, M. V.
- Xing, Z., Pabst, M. J., Hasty, K. A., Smith, R. A. Accumulation of LPS by polyethylene particles decreases bone attachment to implants. Journal of Orthopaedic Research. 24 (5), 959-966 (2006).
- Ding, H., et al. Comparison of the cytotoxic and inflammatory responses of titanium particles with different methods for endotoxin removal in RAW264.7 macrophages. Journal of Materials Science: Materials in Medicine. 23 (4), 1055-1062 (2012).
- Hoogenboom, R., Becer, C. R., Guerrero-Sanchez, C., Hoeppener, S., Schubert, U. S. Solubility and thermoresponsiveness of PMMA in alcohol-water solvent mixtures. Australian Journal of Chemistry. 63 (8), 1173-1178 (2010).
- Efimenko, K., Wallace, W. E., Genzer, J. Surface modification of Sylgard-184 poly(dimethyl siloxane) networks by ultraviolet and ultraviolet/ozone treatment. Journal of Colloid and Interface Science. 254 (2), 306-315 (2002).
- Godek, M. L., Sampson, J. A., Duchsherer, N. L., McElwee, Q., Grainger, D. W. Rho GTPase protein expression and activation in murine monocytes/macrophages is not modulated by model biomaterial surfaces in serum-containing in vitro cultures. Journal of Biomaterials Science. Polymer Edition. 17 (10), 1141-1158 (2006).
- Park, J. S., et al. Involvement of Toll-like Receptors 2 and 4 in Cellular Activation by High Mobility Group Box 1 Protein. Journal of Biological Chemistry. 279 (9), 7370-7377 (2004).
- Ohashi, K., Burkart, V., Flohé, S., Kolb, H. Cutting Edge: Heat Shock Protein 60 Is a Putative Endogenous Ligand of the Toll-Like Receptor-4 Complex. The Journal of Immunology. 164 (2), 558-561 (2000).
- Wong, T., McGrath, J. A., Navsaria, H. The role of fibroblasts in tissue engineering and regeneration. British Journal of Dermatology. 156 (6), 1149-1155 (2007).
- van Wachem, P. B., et al. The influence of protein adsorption on interactions of cultured human endothelial cells with polymers. Journal of Biomedical Materials Research. 21 (6), 701-718 (1987).
- Miller, K. M., Anderson, J. M. Human monocyte/macrophage activation and interleukin 1 generation by biomedical polymers. Journal of Biomedical Materials Research. 22 (8), 713-731 (1988).
- Bonfield, T. L., Colton, E., Anderson, J. M. Plasma protein adsorbed biomedical polymers: Activation of human monocytes and induction of interleukin 1. Journal of Biomedical Materials Research. 23 (6), 535-548 (1989).
- González, O., Smith, R. L., Goodman, S. B. Effect of size, concentration, surface area, and volume of polymethylmethacrylate particles on human macrophages in vitro. Journal of Biomedical Materials Research. 30 (4), 463-473 (1996).
- Anderson, J. M., et al. Protein adsorption and macrophage activation on polydimethylsiloxane and silicone rubber. Journal of Biomaterials Science. Polymer Edition. 7 (2), 159-169 (1995).
- Lord, M. S., Foss, M., Besenbacher, F. Influence of nanoscale surface topography on protein adsorption and cellular response. Nano Today. 5 (1), 66-78 (2010).
- Chen, S., et al. Characterization of topographical effects on macrophage behavior in a foreign body response model. Biomaterials. 31 (13), 3479-3491 (2010).
- Shen, M., Horbett, T. A. The effects of surface chemistry and adsorbed proteins on monocyte/macrophage adhesion to chemically modified polystyrene surfaces. Journal of Biomedical Materials Research. 57 (3), 336-345 (2001).
- Love, R. J., Jones, K. S. The recognition of biomaterials: Pattern recognition of medical polymers and their adsorbed biomolecules. Journal of Biomedical Materials Research Part A. 101 (9), 2740-2752 (2013).
- McNally, A. K., Anderson, J. M. Phenotypic expression in human monocyte-derived interleukin-4-induced foreign body giant cells and macrophages in vitro: Dependence on material surface properties. Journal of Biomedical Materials Research Part A. 103 (4), 1380-1390 (2015).
- Gambhir, V., et al. The TLR2 agonists lipoteichoic acid and Pam3CSK4 induce greater pro-inflammatory responses than inactivated Mycobacterium butyricum. Cellular Immunology. 280 (1), 101-107 (2012).
- Suzuki, O., Yagishita, H., Yamazaki, M., Aoba, T. Adsorption of Bovine Serum Albumin onto Octacalcium Phosphate and its Hydrolyzates. Cells and Materials. 5 (1), 45-54 (1995).
- Johnston, R. L., Spalton, D. J., Hussain, A., Marshall, J. In vitro protein adsorption to 2 intraocular lens materials. Journal of Cataract and Refractive Surgery. 25 (8), 1109-1115 (1999).
- Jin, J., Jiang, W., Yin, J., Ji, X., Stagnaro, P. Plasma proteins adsorption mechanism on polyethylene-grafted poly(ethylene glycol) surface by quartz crystal microbalance with dissipation. Langmuir. 29 (22), 6624-6633 (2013).
- Swartzlander, M. D., et al. Linking the foreign body response and protein adsorption to PEG-based hydrogels using proteomics. Biomaterials. 41, 26-36 (2015).
- Chamberlain, M. D., et al. Unbiased phosphoproteomic method identifies the initial effects of a methacrylic acid copolymer on macrophages. Proceedings of the National Academy of Sciences. 112 (34), 10673-10678 (2015).
- Dillman, W. J., Miller, I. F. On the adsorption of serum proteins on polymer membrane surfaces. Journal of Colloid And Interface Science. 44 (2), 221-241 (1973).
- Ishihara, K., Ziats, N. P., Tierney, B. P., Nakabayashi, N., Anderson, J. M. Protein adsorption from human plasma is reduced on phospholipid polymers. Journal of Biomedical Materials Research. 25 (11), 1397-1407 (1991).
- Warkentin, P., Wälivaara, B., Lundström, I., Tengvall, P. Differential surface binding of albumin, immunoglobulin G and fibrinogen. Biomaterials. 15 (10), 786-795 (1994).
- Berghaus, L. J., et al. Innate immune responses of primary murine macrophage-lineage cells and RAW 264.7 cells to ligands of Toll-like receptors 2, 3, and 4. Comparative Immunology, Microbiology and Infectious Diseases. 33 (5), 443-454 (2010).
- Zhang, Y., Karki, R., Igwe, O. J. Toll-like receptor 4 signaling: A common pathway for interactions between prooxidants and extracellular disulfide high mobility group box 1 (HMGB1) protein-coupled activation. Biochemical Pharmacology. 98 (1), 132-143 (2015).
- Mizel, S. B., Honko, A. N., Moors, M. A., Smith, P. S., West, A. P. Induction of macrophage nitric oxide production by Gram-negative flagellin involves signaling via heteromeric Toll-like receptor 5/Toll-like receptor 4 complexes. Journal of Immunology. 170 (12), 6217-6223 (2003).
- Das, N., et al. HMGB1 Activates Proinflammatory Signaling via TLR5 Leading to Allodynia. Cell Reports. 17 (4), 1128-1140 (2016).
- Pelegrin, P., Barroso-Gutierrez, C., Surprenant, A. P2X7 Receptor Differentially Couples to Distinct Release Pathways for IL-1β in Mouse Macrophage. The Journal of Immunology. 180 (11), 7147-7157 (2008).
- Tak, P. P., Firestein, G. S. NF-κB: A key role in inflammatory diseases. Journal of Clinical Investigation. 107 (1), 7-11 (2001).
- Ashkenazi, A., Dixit, V. M. Death receptors: signaling and modulation. Science. 281 (5381), 1305-1308 (1998).
- Erridge, C. Endogenous ligands of TLR2 and TLR4: agonists or assistants. Journal of Leukocyte Biology. 87 (6), 989-999 (2010).
- Feng, Y., et al. A macrophage-activating, injectable hydrogel to sequester endogenous growth factors for in situ angiogenesis. Biomaterials. 134, 128-142 (2017).
- Lonez, C., et al. Cationic lipid nanocarriers activate Toll-like receptor 2 and NLRP3 inflammasome pathways. Nanomedicine: Nanotechnology, Biology, and Medicine. 10 (4), 775-782 (2014).
