

Summary
Este protocolo proporciona a los investigadores un método rápido e indirecto para medir la actividad del factor de transcripción NF--B/AP-1 dependiente de TLR en una línea celular de macrófagomur en respuesta a una variedad de superficies poliméricas y capas de proteínas adsorbidas que modelan el microambiente de implantes de biomaterial.
Abstract
La persistente respuesta inflamatoria del huésped a un biomaterial implantado, conocido como reacción del cuerpo extraño, es un desafío significativo en el desarrollo e implementación de dispositivos biomédicos y construcciones de ingeniería de tejidos. Los macrófagos, una célula inmune innata, son actores clave en la reacción del cuerpo extraño porque permanecen en el sitio del implante durante la vida útil del dispositivo, y se estudian comúnmente para obtener una comprensión de esta respuesta perjudicial del huésped. Muchos investigadores de biomateriales han demostrado que las capas de proteínas adsorbidas en materiales implantados influyen en el comportamiento de los macrófagos y, posteriormente, afectan la respuesta del huésped. Los métodos de este artículo describen un modelo in vitro utilizando capas de proteínas adsorbidas que contienen moléculas de daño celular en superficies de biomaterial espolimérico para evaluar las respuestas de los macrófagos. Se utilizaron como método rápido para examinar indirectamente la actividad del factor de transcripción NF-B/AP-1 en respuesta a complejas capas de proteínas absorbentes que contienen proteínas sanguíneas y patrones moleculares asociados al daño, como modelo de las complejas capas de proteínas adsorbidas formadas en superficies biomateriales in vivo.
Introduction
La reacción del cuerpo extraño (FBR) es una respuesta crónica del huésped que puede afectar negativamente el rendimiento de un material o dispositivo implantado (por ejemplo, dispositivos de administración de medicamentos, biosensores), a través de la liberación persistente de mediadores inflamatorios y al impedir la integración entre el material implantado y el tejido circundante1. Esta respuesta inmune innata se inicia por el procedimiento de implantación y se caracteriza por la presencia a largo plazo de células inmunitarias innatas y la formación de cápsulas fibrosas alrededor del implante1. En el contexto de las respuestas materiales del huésped, las interacciones entre macrófagos y materiales tienen un impacto significativo en la progresión de la respuesta del huésped y el desarrollo de un FBR1. Los macrófagos son una población de células inmunitarias innatas diversas, reclutadas en el sitio del implante, ya sea de poblaciones de macrófagos residentes en tejidos o de la sangre como macrófagos derivados de monocitos. Comienzan a acumularse en el sitio del implante poco después de la implantación, y en cuestión de días se convierten en la población celular predominante en el microambiente del implante. Los macrófagos adherentes a los materiales, junto con las células gigantes de cuerpo extraño (FBGC) formadas a través de la fusión de macrófagos, pueden persistir en la superficie del material durante la vida útil del implante2,3. En consecuencia, los macrófagos se consideran actores clave en la respuesta del cuerpo extraño debido a sus funciones orquestando los pasos característicos de la FBR: respuesta inflamatoria aguda, remodelación tisular y formación de tejido fibroso1.
Los receptores similares a los peajes (TLR) son una familia de receptores de reconocimiento de patrones que son expresados por muchas células inmunitarias, incluyendo macrófagos, y se ha demostrado que juegan un papel importante en la inflamación y la cicatrización de heridas. Además de los ligandos derivados de patógenos, los TLR son capaces de unir moléculas endógenas, conocidas como patrones moleculares asociados al daño (DAPP), que se liberan durante la necrosis celular y activan las vías de señalización inflamatoria que resultan en la producción de citoquinas proinflamatorias4. Nosotros y otros hemos propuesto que el daño incurrido durante los procedimientos de implantación de biomateriales de tejido blando libere a los DAMPs, que luego se adsorbe a las superficies biomateriales además de las proteínas de la sangre y modulalas las interacciones entre células y materiales subsiguientes5,6. Cuando los macrófagos interactúan con la capa de proteína adsorbida en un implante, sus TDR de superficie pueden reconocer los DPR adsorbidas y activar cascadas de señalización proinflamatoria, lo que conduce a la activación del factor de transcripción NF-B y AP-1 y a la producción de citoquinas proinflamatorias. Hemos demostrado anteriormente que los macrófagos murinos han aumentado significativamente la actividad de NF-B/AP-1 y el factor de necrosis tumoral (TNF-- secreción proinflamatoria de citoquinas) en respuesta a las capas de proteína adsorbida que contienen DAMP en una variedad de superficies poliméricas en comparación con superficies con suero o plasma adsorbida solamente (es decir, no hay DAPP presentes), y que esta respuesta está mediada en gran medida por TLR2, mientras que TLR4 desempeña un papel menor5.
La línea celular de macrófagos de reportero NF-B/AP-1utilizadaen este protocolo es un método conveniente para medir la actividad relativa NF--B y AP-1 en macrófagos5,7,8. En combinación con los inhibidores de la vía TLR, esta línea celular es una herramienta útil para investigar la activación de TLR y su papel en la inflamación en respuesta a una variedad de estímulos5,7,8. Las células reporteras son una línea celular modificada similar a un macróforo de ratón que puede producir establemente fosfatasa alcalina embrionaria secreta (SEAP) sobre la activación del factor de transcripción NF-B y AP-19. El ensayo fosfatasa alcalina colorimétrica enzimática(Tabla de materiales) se puede utilizar para cuantificar las cantidades relativas de expresión SEAP como una medida indirecta de la actividad NF-B/AP-1. Como NF-B y AP-1 son aguas abajo de muchas vías de señalización celular, se pueden utilizar anticuerpos e inhibidores neutralizantes dirigidos a TlR específicos (por ejemplo, TLR2) o moléculas de adaptador TLR (por ejemplo, MyD88) para verificar el papel de una vía específica. La metodología descrita en este artículo proporciona un enfoque sencillo y rápido para evaluar la contribución de la señalización TLR en las respuestas de macrófagos murinos a una variedad de superficies poliméricas con capas de proteínas adsorbidas que contienen proteínas de la sangre y DAPP como modelo in vitro de biomateriales implantados.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. Preparación de medios y reactivos
- Prepare los medios de fibroblastos. Combinar 450 ml del medio águila modificado de Dulbecco (DMEM), 50 ml de suero bovino fetal (FBS) y 5 ml de penicilina/estreptomicina. Conservar a 4oC durante un máximo de 3 meses.
- Prepare medios de crecimiento de macrófagos reporteros en alícuotas de 50 ml. Combinar 45 ml de DMEM, 5 ml de FBS, 5 g/ml de reactivo de eliminación de micoplasma(Tabla de materiales)y 200 g/ml de fleomicina D1(Tabla de materiales). Conservar a 4oC durante un máximo de 3 meses.
- Prepare los medios de ensayo de macrófagos de reportero en alícuotas de 50 ml. Combinar 45 ml de DMEM, 5 ml de FBS inactivado por calor (HI-FBS), 5 g/ml de reactivo de eliminación de micoplasma y 200 g/ml de fleleomicina D1. Conservar a 4oC durante un máximo de 3 meses.
2. Recubrimiento superficies de cultivo celular con poli(metil metacrilato)
- Disolver poli(metil metacrilato) (PMMA) en cloroformo a 20 mg/ml (por ejemplo, 100 mg de PMMA en 5 ml de cloroformo) en un vial de centelleo de vidrio de 20 ml. Coloque una barra de agitación magnética en el vial y deje que se revuelva durante al menos 2 horas, hasta que todos los sólidos se disuelvan.
PRECAUCION: El cloroformo es dañino si se inhala. Asegúrese de usar disolvente en una campana de humos mientras usa guantes de PVA. - Pipetear 400 l de solución de PMMA en el centro de un portaobjetos de microscopio de borosilicato en una capa de centrifugado, y girar a 3000 rpm durante 2 min. Prepare el número de diapositivas necesarias para el ensayo, así como 3 x 5 adicionales para la medición del ángulo de contacto con el agua. Almacene los portaobjetos en una caja limpia (pulverizadas y limpiadas con 70% de etanol) para su uso futuro.
NOTA: El recubrimiento de espín se utiliza a menudo para depositar un recubrimiento delgado y uniforme en una superficie plana. Un revestimiento de espín gira un sustrato a altas velocidades, utilizando la fuerza centrífuga para extender la solución de recubrimiento sobre la superficie.- Mida el ángulo de contacto del agua en dos posiciones aleatorias en la superficie de los portaobjetos recubiertos adicionales (es decir, no los portaobjetos que se utilizan para el cultivo celular) con un goniómetro para garantizar que la superficie de vidrio esté completamente recubierta con el polímero.
NOTA: Sólo se debe utilizar agua de la más alta pureza (por ejemplo, vidrio triple destilado) para mediciones de ángulo de contacto con el agua.
- Mida el ángulo de contacto del agua en dos posiciones aleatorias en la superficie de los portaobjetos recubiertos adicionales (es decir, no los portaobjetos que se utilizan para el cultivo celular) con un goniómetro para garantizar que la superficie de vidrio esté completamente recubierta con el polímero.
- En un armario de seguridad biológica (BSC) conecte los pozos pegajosos de 8 cámaras a los portaobjetos recubiertos de PMMA utilizando fórceps estériles y siguiendo la técnica aséptica. Presione firmemente en la parte superior de los pozos pegajosos para asegurarse de que están fuertemente unidos. Incubar los portaobjetos con pozos pegajosos unidos a 37 oC durante la noche para asegurar el sello.
- Pruebe el sello de los pozos pegajosos añadiendo 200 ml de agua de grado de cultivo celular (libre de endotoxinas) a cada pocillo. Incubar a temperatura ambiente (RT) durante 60 minutos y asegurar se quede sin fugas antes de proceder. Aspirar el agua, teniendo cuidado de no molestar el recubrimiento de PMMA.
- Realice lavados de agua sin endotoxinas añadiendo 300 ml de agua libre de endotoxinas a cada poca y incubando durante 1 h (tres veces), 12 h y 24 horas antes de su uso para eliminar cualquier disolvente restante.
- Pruebe la concentración de endotoxinas de las diapositivas que se utilizarán para el cultivo celular. Incubar 200 l de agua reactivo libre de endotoxinas(Tabla de materiales) en un pozo de cada diapositiva durante 1 h. Medir la concentración de endotoxinas en el extracto utilizando un ensayo de endotoxina cromogénica de punto final (Tabla de materiales).
NOTA: El siguiente protocolo es específico del kit de ensayo de endotoxinas que figura en la Tabla de materiales. - Utilice únicamente agua y consumibles (es decir, puntas de pipeta, tubos de microcentrífuga y placas de pozos) que estén certificados libres de pirógenos (es decir, libres de endotoxinas) para este trabajo. Además, cualquier cristalería utilizada en la preparación de las superficies recubiertas de polímero debe despirarse utilizando esterilización por calor seco (250 oC durante 30 minutos) antes de utilizar10. La medición de la endotoxina en la solución de extracto, como se describe aquí, puede dar lugar a una subestimación de la endotoxina en la superficie del material11,12. En consecuencia, se recomienda que al desarrollar un protocolo de recubrimiento de polímeros, realice la reacción de ensayo de endotoxina (es decir, los pasos 2.5.4-2.5.6 para las muestras de ensayo [agua reactiva] o controles de espiga) directamente dentro de los pozos que contienen la muestra recubierta para asegurarse de que no se introducen inadvertidamente en el sistema fuentes de endotoxinas durante el proceso de recubrimiento.
- Lleve todas las muestras de ensayo (es decir, extractos) y los reactivos de ensayo de endotoxinas a RT. Reconstituya el reactivo cromogénico en el tampón de ensayo y la norma de endotoxina en agua reactiva, permita disolverse durante 5 minutos y revuelva suavemente antes de usarlo. Cubra todas las botellas con película de parafina cuando no esté en uso.
- Cree una curva de dilución estándar de 5 x 8 puntos del estándar de endotoxina que va desde el límite inferior al límite superior del ensayo mediante la realización de una dilución en serie del estándar de endotoxina en agua reactivo.
- Para controlar la mejora o inhibición del ensayo de endotoxina en muestras de prueba, prepare un control positivo (también llamado control de picos o muestra salpicada) diluyendo una cantidad conocida de endotoxina en la solución de muestra de prueba no utilizada.
NOTA: La concentración del control positivo debe ser la misma concentración que una norma en el centro de la curva estándar. Si la cantidad recuperada del pico de la endotoxina (es decir, la concentración del control positivo menos la concentración de la muestra de ensayo sin pico) está dentro del 50-200% de la concentración nominal del pico de la endotoxina, se puede considerar que la solución de extracción no interfiere significativamente con el ensayo. - Agregue 50 s de estándares, muestras o controles de púas a cada pocto de una placa de 96 pocillos en duplicado o triplicado. Utilice agua reactivo como control negativo.
- Añadir 50 ml de reactivo cromogénico a cada pocómetro. Agregue el reactivo rápidamente a todos los pozos. Utilice un temporizador para registrar la cantidad de tiempo que se tarda en agregar reactivo a todos los pozos. Cubra la placa con un sello adhesivo e incubar a 37 oC (el tiempo de incubación depende del lote y se indica en el certificado de análisis incluido en el kit de reactivos cromogénicos). Alternativamente, compruebe la placa cada 15 minutos durante la incubación hasta que se observe el cambio de color en todos los pozos estándar.
- Después de la incubación, añadir 25 s l de ácido acético 50% a cada poca (concentración final de 10% ácido acético por poca) para detener la reacción. Añadir ácido acético en el mismo orden en que se añadió el reactivo cromogénico. Lea la absorbancia de la placa utilizando un lector de placas a 405 nm. Aspirar líquido y desechar la placa.
NOTA: La adición de ácido acético debe tardar el mismo tiempo en añadira a cada uno bien que el reactivo cromogénico (30 s).
- Ultravioleta (UV) esteriliza las diapositivas durante 30 minutos antes de los experimentos de cultivo celular.
3. Recubrimiento superficies de cultivo celular con polidimetilsiloxano
- Mezclar el elastómero de polidimetilsiloxano (PDMS) en una relación de peso de 10:1 (base:agente de curado). En un armario de seguridad biológica, pipeta aproximadamente 10 ml de base de polidimetilsiloxano en un tubo estéril. Pesar el tubo y añadir lentamente el agente de curado hasta que se haya añadido un 10%.
PRECAUCION: Utilice reactivos PDMS en un área bien ventilada y evite el contacto visual con gafas de seguridad. - Mezcle bien el elastómero revolviendo con una punta de pipeta serológica estéril y pipeteando hacia arriba y hacia abajo. Agregue aproximadamente 200 s de la solución a cada pocal de una placa de 48 pocillos. Incline la placa de pozo lentamente para asegurar una cobertura completa de los pozos con solución de elastómero.
- Coloque la placa de pozo con elastómero en un horno al vacío a 50 cmHg, 40 oC. Retire la tapa y la cubierta con una toallita de una sola hoja para evitar que otros residuos caigan en los pozos. Dejar incubar durante al menos 48 h.
- Confirme que los pozos están completamente recubiertos a través de inspección visual. Asegúrese de que el elastómero esté completamente curado mediante una proddesción suave con una punta de pipeta estéril antes de retirarlo.
- Añadir 300 l de etanol 70% (hecho con etanol absoluto y agua libre de endotoxinas) e incubar a RT durante 1 h. Retire el etanol y realice lavados de agua libres de endotoxinas añadiendo 300 ol de agua libre de endotoxinas a cada pozo e incubando durante 1 h (tres veces), 12 h , y 24 h antes de su uso para eliminar cualquier disolvente restante.
- Incubar 200 l de agua libre de endotoxinas en tres pocillos de cada placa durante 1 h. Medir la concentración de endotoxinas de los extractos de agua utilizando un ensayo de endotoxina cromogénica de punto final (pasos 2.5.1-2.5.6).
4. Superficies de cultivo celular de recubrimiento con poli fluorado (tetrafluoroetileno)
- Hacer una solución de 1 mg/ml de polifluorado (tetrafluoroetileno) (fPTFE) (por ejemplo, añadir 10 mg de fPTFE a 10 ml de disolvente fluorado [Tabla de materiales]) en un vial de centelleo de vidrio de 20 ml. Coloque una barra de agitación magnética en el vial y deje que se revuelva durante al menos 24 horas, hasta que todos los sólidos se disuelvan.
- Añadir aproximadamente 150 l de la solución de polímero a cada pocal de una placa de 48 pocillos de poliestireno (es decir, no el cultivo tisular tratado). Incline la placa de pozo lentamente para asegurar una cobertura completa de todos los pozos con solución de polímero. Vuelva a colocar la tapa.
- Para garantizar un recubrimiento eficaz de fPTFE de los pozos, los revestimientos de vidrio deben revestirse en fPTFE y utilizarse para la medición del ángulo de contacto del agua (paso 4.3.1). Coloque los labios de las cubiertas dentro de los pocillos de una placa de 24 pocillos. Añadir aproximadamente 400 l de la solución de polímero a cada pocal que contenga un cubreobjetos. Empuje los labios de las cubiertas hacia abajo usando fórceps estériles, asegurándose de que estén completamente cubiertos de solución de polímero, y cubra la placa del pozo con una tapa.
- Coloque la placa de pozo con solución de polímero y/o cubiertas en un horno al vacío a 50 cmHg, 40 oC. Retire la tapa y la cubierta con una toallita de una sola hoja para evitar que otros residuos caigan en los pozos. Dejar incubar durante al menos 48 h.
- Mida el ángulo de contacto con el agua de los revestimientos recubiertos de fPTFE con un goniómetro para garantizar un recubrimiento eficaz.
NOTA: Sólo se debe utilizar agua de la más alta pureza (por ejemplo, vidrio triple destilado) para mediciones de ángulo de contacto con el agua.
- Mida el ángulo de contacto con el agua de los revestimientos recubiertos de fPTFE con un goniómetro para garantizar un recubrimiento eficaz.
- Añadir 300 l de etanol 70% (hecho con etanol absoluto y agua libre de endotoxinas) e incubar a RT durante 1 h. Retire el etanol y realice lavados de agua libres de endotoxinas añadiendo 300 ol de agua libre de endotoxinas a cada pozo e incubando durante 1 h (tres veces), 12 h , y 24 h antes de su uso para eliminar cualquier disolvente restante.
- Incubar 200 l de agua libre de endotoxinas en tres pozos de cada placa durante 1 h. Medir la concentración de endotoxinas de extractos de agua utilizando un ensayo de endotoxina cromogénica de punto final (pasos 2.5.1 a 2.5.6).
- LOS rayos UV esterilizan las placas de pozo durante 30 minutos antes de los experimentos de cultivo celular.
5. Hacer lisado a partir de células 3T3
- Cultivar células 3T3 en múltiples matraces T150 a 70% de confluencia. Para separar las células, aspirar medios, lavar la superficie con 5 ml de PBS y aspirar PBS. Añadir 5 ml de enzima de disociación celular recombinante libre de origen animal(Tabla de materiales)e incubar a 37 oC durante 3 x 5 min.
- Separe las células inclinando suavemente el matraz hacia adelante y hacia atrás. Añadir 5 ml de PBS para neutralizar la enzima recombinante utilizada para la disociación celular. Transfiera las células separadas de los matraces a un tubo centrífugo y mezcle a través de pipeteo. Realice un recuento de células vivas utilizando un hemocitómetro y un tinte de viabilidad celular.
NOTA: Se seleccionó una enzima disociación celular que puede ser neutralizada a través de la dilución en PBS para evitar la introducción de proteínas a base de suero en la preparación de lisato. Si se utiliza tripsina para disociar las células, debe neutralizarse con una solución que contenga suero, y se debe realizar un lavado adicional de PBS para reducir la cantidad de proteínas séricas transportadas a la preparación de lisado. - Centrifugar las células a 200 x g durante 5 min. Aspirar el sobrenadante y resuspender las células en el volumen original (es decir, 10 ml x número de matraces) de PBS para lavar cualquier medio restante. Repetir.
- Centrifugar las células de nuevo a 200 x g durante 5 min y aspirar el sobrenadante. Agregue el volumen de PBS necesario para lograr una concentración celular final de 1 x 106 células/ml. Colocar la solución celular en un congelador de -80 oC hasta que la muestra esté completamente congelada (al menos 2 h).
- Descongelar la solución celular en un baño de agua a 37 oC. Una vez descongelado completamente, vuelva a colocar la solución en el congelador de -80 oC hasta que esté totalmente congelada. Repita para un total de 3 ciclos de congelación-descongelación.
- Realizar un ensayo de ácido micro bicininicónico (BCA) en el lisado celular en una variedad de diluciones (por ejemplo, 1/100, 1/200, 1/500, 1/1000) para determinar la concentración de proteínas. Diluir el lisado celular a una concentración proteica de 468,75 g/ml, alícuota y almacenar a -80 oC para su uso futuro.
NOTA: La concentración final de proteína en una placa de 48 pocillos es de 125 g/cm2 (basada en la superficie de un pozo, 0,75 cm2). - Realizar una mancha occidental para evaluar la presencia de DPP en el lisato (por ejemplo, proteína de choque térmico 60 [HSP60], caja de grupo de alta movilidad 1 [HMGB1]) cargando 40 x 60 g de proteína de lisado en el búfer de carga en un gel de poliacrilamida de 1,5 mm de espesor y 10% y siga la blotdemida occidental estándar en un gel de poliacrilamida occidental de 1,5 mm de espesor y siga la blota occidental estándar y siga la blota occidental estándar en un gel de poliacrilamida occidental estándar y siga la blot western estándar Procedimientos.
6. Evaluación del efecto de las capas de proteínas absorbidas y los receptores similares al peaje en la actividad NF-B de los macrófagos
NOTA: Para obtener un esquema del flujo de trabajo experimental y el diseño de placas, consulte la Figura 1A y la Figura suplementaria 1, respectivamente.
- Cultiva los macrófagos de reportero en un matraz de tamaño adecuado a un 70% de confluencia. Aspirar medios, lavar la superficie con PBS y aspirar PBS. Añadir la enzima de disociación de células recombinantes e incubar a 37oC durante 8 min.
- Separe las células tocando firmemente los lados del matraz. Inactivar la enzima disociación celular recombinante añadiendo un volumen igual de medios de crecimiento (que contiene 10% FBS). Realice un recuento de células vivas utilizando un hemocitómetro y un tinte de viabilidad celular.
NOTA: La viabilidad esperada para los macrófagos del reportero después de una incubación de 8 minutos en la enzima de disociación celular es del 90%. - Células centrífugas a 200 x g durante 5 min. Aspirate sobrenadant y resuspenden en volumen original de PBS para lavar las células. Centrifugar de nuevo y resuspender las células a 7,3 x 105 células/ml en medios de ensayo (que contienen FBS inactivado por calor).
- Separe la suspensión celular en 3 tubos diferentes: inhibidor TLR4, anti-TLR2 y sin tratar. Incubar células con 1 inhibidor de TLR4 g/ml durante 60 min a RT o con 50 g/ml anti-TLR2 durante 30 min a RT.
- Añadir 200 ml de lisato, 10% FBS, 10% plasma de ratón comercial(Tabla de materiales),o una mezcla de las soluciones proteicas a una placa de 48 pocillos (o equivalente) y permitir que la proteína se absorba a 37 oC durante el tiempo deseado (es decir, 30 min, 60 min o 24 h). Aspirar las soluciones proteicas de los pozos, utilizando una pipeta Pasteur fresca para cada solución proteica, y lavar superficies con 250 ml de PBS durante 5 min. Repita el proceso para un total de 3 lavados.
NOTA: Es posible que este paso deba iniciarse antes en el protocolo en función del tiempo de adsorción deseado. Ajuste el protocolo en consecuencia. - Después del período de incubación con el inhibidor TLR4 o anti-TLR2, las células de pipeta se resuspenden. Añadir 200 l de solución celular a cada poca.
- Para la condición de control positivo TLR2, agregue Pam3CSK4 a una concentración final de 150 ng/mL. Para la condición de control positivo TLR4, agregue lipopolisacárido (LPS) a una concentración final de 1,5 g/ml. Incubar células a 37oC durante 20 h.
- Muestre 20 ml de sobrenadante de cada pocal y placa en duplicado en una placa de 96 pocillos. Incluya tres pozos de medios de ensayo de 20 l como control de fondo. Añadir 200 ol de reactivo de ensayo de reportero SEAP a cada pozo. Cubrir la placa con un sello adhesivo e incubar durante 2,5 h a 37 oC.
NOTA: El tiempo de incubación puede variar dependiendo de las condiciones experimentales, y debe optimizarse para una fuerte diferencia en la absorción entre los pozos de control positivos y negativos.- Transfiera el resto del sobrenadante a un tubo de 1,5 ml (por poca). Centrífuga a 1.000 x g durante 10 minutos para peletizar cualquier escombro. Transfiera el sobrenadante a un nuevo tubo de 1,5 ml y guárdelo a -80 oC. Analizar el sobrenadante para detectar la presencia de citoquinas proinflamatorias (p. ej., TNF-, interleucina 6) a través del ensayo inmunoabsorbente ligado a enzimas (ELISA).
- Retire el sello de la placa adhesiva. Lea la absorbancia de la placa utilizando un lector de placas a 635 nm. Aspirar líquido y desechar la placa.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Se probaron métodos de limpieza para las superficies recubiertas de polímero para garantizar que no hubiera interrupción del recubrimiento, lo que se vería como un cambio en el ángulo de contacto del agua a un revestimiento de vidrio sin recubrimiento(Figura 2). Se encontró que el remojo de los portaobjetos recubiertos con PMMA en etanol 70% durante 1 h elimina el recubrimiento de PMMA(Figura 2, panel izquierdo), probablemente debido a la solubilidad de PMMA en 80 wt% etanol13, por lo tanto, las superficies recubiertas de PMMA se limpiaron utilizando 30 minutos de esterilización UV solamente. La concentración de PMMA para recubrimiento se optimizó previamente5. Se utilizó un remojo de etanol de 1 h 70% para limpiar PDMS, y se descuidó la esterilización UV, ya que la luz UV puede causar escisiones en cadena e influir en las propiedades de humectación de la superficie de PDMS14. Tanto el 70% de remojo de etanol como la esterilización UV no influyeron en el ángulo de contacto con el agua de los revestimientos recubiertos de fPTFE(Figura 2, panel derecho), por lo tanto, los dos métodos, sucesivamente, se utilizaron para limpiar recubrimientos de fPTFE. El método de recubrimiento fPTFE fue descrito previamente por el grupo15de Grainger.
Una mancha occidental se realizó en el lisado 3T3 para asegurar que las especies de DAMP estuvieran presentes en la compleja mezcla molecular. Los resultados mostraron que tanto HMGB1 como HSP60, dos DAP bien documentados16,17, estaban presentes en el isla (Figura 1B). La adsorción de ligandos TLR del lisado sobre las superficies de polímeros se confirmó mediante el cultivo de macrófagos de reportero (sin tratar, TLR2 neutralizado, o TLR4 inhibido) durante 20 h en superficies de polímeros adsorbidos por proteínas (es decir, poliestireno tratado con cultivo de tejidos [TCPS], PMMA, PDMS, fPTFE), y luego evaluar indirectamente la actividad nf-B/AP-1 en función de la producción seDE mediante un ensayo enzimático(Figura 1C y Figura 3). Además, los macrófagos de los reporteros habían aumentado significativamente la actividad de NF-B/AP-1 en el iscaso adsorbido en comparación con el FBS adsorbido o el plasma y ninguna proteína pre-adsorbida (medios)(Figura 4). TlR ligandos lipopéptido triacilado sintético (Pam3CSK4, ligando TLR2) y lipopolisacárido (LPS, ligando TLR4) se incluyeron como controles positivos para confirmar el anticuerpo o inhibidor y el ensayo funcionaba correctamente. La neutralización de TLR2 tuvo una reducción notablemente más fuerte en la respuesta NF-B/AP-1 de los macrófagos de reporteros al iscaso adsorbida en comparación con la inhibición de TLR4. Además, pequeñas cantidades de lisato diluidos en suero (basado en proteína total) indujeron significativamente mayor respuesta NF-B/AP-1 en comparación con el suero solo, con la dilución efectiva más baja dependiente de la superficie del polímero(Figura 5). Estos resultados demuestran la potencia de las moléculas derivadas del lisado adsorbida en la inducción de la actividad NF--B/AP-1 dependiente de TLR en macrófagos de reportero en una variedad de superficies poliméricas.

Figura 1: Métodos y resultados para el ensayo de fosfatasa alcalina de macrófagos de reporteroNF-B/AP-1 en TCPS, PMMA, PDMS y fPTFE. (A) Diagrama del flujo de trabajo para el ensayo de fosfatasa alcalina de macrófagos reportero. (B) Mancha occidental de lisato que confirma la presencia de las especies DAMP HMGB1 y HSP60, con la actina como control de carga. (C) actividad NF--B/AP-1 (representada por la absorbancia) de macrófagos reporteros cultivados en medios (control negativo), 10% FBS, lisado y Pam3CSK4 (ligando TLR2, control positivo) durante 20 h. Los datos muestran los resultados de un experimento y son representativos de los resultados de al menos 2 experimentos separados, mostrados como media de desviación estándar (SD). Cada experimento utilizaba n x 3 pozos separados por condición, y cada pozo estaba chapado en duplicado para el ensayo enzimático. Analizado mediante la prueba post-hoc de ANOVA unidireccional y Tukey. p < 0.001. Esta figura ha sido adaptada con el permiso de McKiel y Fitzpatrick5. Copyright 2018 American Chemical Society. Haga clic aquí para ver una versión más grande de esta figura.

Figura 2: Optimización de los métodos de limpieza para superficies recubiertas de PMMA y fPTFE, evaluadas mediante ángulo de contacto con agua (WCA). Las mediciones se tomaron en 2 puntos separados de al menos 3 tapas. Los datos se muestran como medias: SD. Analizados utilizando la prueba post-hoc de ANOVA unidireccional y Tukey. * p < 0.05. Esta figura ha sido adaptada con el permiso de McKiel y Fitzpatrick5. Copyright 2018 American Chemical Society. Haga clic aquí para ver una versión más grande de esta figura.

Figura 3: Actividad DE NF--B/AP-1 mediada por TLR (representada por la absorbancia) de macrófagos de reportero cultivados en 10% FBS (control), licuado y control positivo durante 20 h. (A) Influencia de la neutralización de TLR2 en las respuestas de los macrófagos de reportero al islato adsorbido. El control positivo es Pam (Pam3CSK4, ligando TLR2). (B) Influencia de la inhibición de TLR4 en la respuesta de los macrófagos del reportero al islato adsorbido. El control positivo es LPS (ligando TLR4). Los datos muestran los resultados de un experimento y son representativos de los resultados de al menos 2 experimentos separados, mostrados como medias : SD. Cada experimento utilizaba n x 3 pozos separados por condición, y cada pozo estaba chapado en duplicado para el ensayo enzimático. Analizado mediante la prueba post-hoc de ANOVA unidireccional y Tukey. ** p < 0.01, *** p < 0.001. Esta figura ha sido adaptada con el permiso de McKiel y Fitzpatrick5. Copyright 2018 American Chemical Society. Haga clic aquí para ver una versión más grande de esta figura.

Figura 4: Actividad NF--B/AP-1 (representada por la absorbancia) de macrófagos reportero cultivados en medios (control negativo), capas de proteína adsorbida de 30 min y 24 h, y Pam3CSK4 (control positivo) en TCPS durante 20 h. Los datos se combinan a partir de 3 experimentos separados y se muestran como media sd. Cada experimento utilizaba n x 3 pozos separados por condición, y cada pozo estaba chapado en duplicado para el ensayo enzimático (es decir, n 9 pozos de cultivo celular no independientes y n a 18 pozos de ensayo enzimático no independientes). Analizado mediante la prueba post-hoc de ANOVA unidireccional y Tukey. p < 0.001. Haga clic aquí para ver una versión más grande de esta figura.
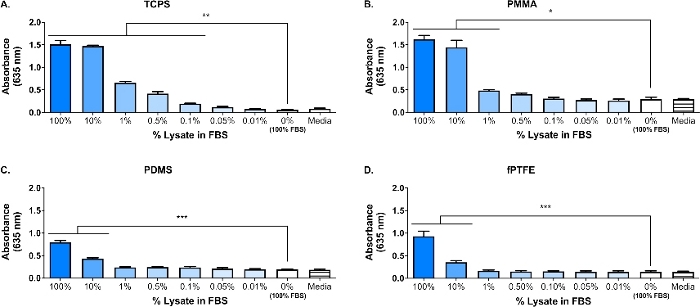
Figura 5: Actividad de macrófagos de reportero NF--B/AP-1 (representada por absorbancia) después de 20 h en respuesta a diluciones de lisato en FBS (proteína total de 280 g/pozo) adsorbida a superficies de polímero durante 30 min. (A) TCPS. (B) PMMA. (C) PDMS. (D) fPTFE. Los datos muestran los resultados de un experimento y son representativos de los resultados de al menos 2 experimentos separados, mostrados como medias : SD. Cada experimento utilizaba n x 3 pozos separados por condición, y cada pozo estaba chapado en duplicado para el ensayo enzimático. Analizado mediante la prueba post-hoc de ANOVA unidireccional y Tukey. * p < 0.05, ** p < 0.01, *** p < 0.001. Esta figura ha sido adaptada con el permiso de McKiel y Fitzpatrick5. Copyright 2018 American Chemical Society. Haga clic aquí para ver una versión más grande de esta figura.

Figura suplementaria 1: Diseños de ejemplo utilizados para el ensayo de cultivo de células de macrófagos de reportero NF-B/AP-1 en formatos de placa de 8 y 48 pocillos. Haga clic aquí para ver una versión más grande de esta figura.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Un enfoque principal de nuestro laboratorio es la respuesta del huésped a los implantes de tejido blando biomaterial sólido, y en particular cómo el daño celular incurrido durante el procedimiento de implantación afecta la respuesta del huésped. El trabajo presentado aquí describe experimentos preliminares utilizando una línea celular de macrófagos de reportero y un lisato celular que contiene DAMP generado in vitro, para investigar la influencia de las moléculas liberadas durante el daño celular (es decir, de la cirugía de implante) en las respuestas de los macrófagos a los biomateriales. El lisato de células fibroblastas se utilizó para modelar el daño celular y la liberación de los DAPP debido a la colocación del biomaterial. Los fibroblastos fueron elegidos para crear el lisado debido a la prevalencia de fibroblastos en el tejido blando, así como su capacidad para secretar una variedad de proteínas de matriz extracelular (ECM), incluyendo la fibronectina18. El ciclo de congelación de savia fue elegido como el método de lisis para producir DAPP intracelulares y derivados de ECM, similar a lo que estaría presente en el entorno del implante. Los inhibidores de la proteasa no se utilizaron para hacer este isla. Mientras que la lisis celular no controlada como el ciclo de congelación de descongelación puede resultar en la liberación de proteasas que pueden degradar los DAPS, estas enzimas también probablemente estarían presentes en el entorno del implante biomaterial cuando las células se dañan durante el procedimiento de implantación. La presencia de DMIP en la compleja mezcla molecular del islao fue confirmada por Western blot(Figura 1B; HMGB1 y HSP60) y el ensayo de reportero SEAP(Figura 1C; Actividad NF--B/AP-1 en respuesta al islate adsorbido). También hemos realizado ensayos en los que el islato se diluyó en FBS a base de la concentración total de proteínas y se adsorbicó en superficies de cultivo celular(Figura 5)para reflejar mejor la complejidad del entorno del implante, ya que contendrá una abundancia de proteínas sanguíneas, así como DAPP6. La actividad de los macrófagos de reportero NF-B/AP-1 se mantuvo significativamente incrementada en las capas adsorbidas a partir del lisado diluido en FBS, y la dilución más baja para lograr una activación significativa fue dependiente de la superficie, que oscilaba entre el 0,1% (TCPS) y el 10% (PDMS y fPTFE).
Los polímeros PMMA, PDMS y PTFE fueron elegidos para este trabajo porque no son degradables y se han utilizado ampliamente en la literatura para evaluar la adsorción de proteínas y la respuesta de macrófagos a biomateriales19,20,21,22,23,24,25. TCPS también se utilizó para la comparación ya que es un sustrato común utilizado para macrófagos in vitro y trabajos de señalización TLR21,26,27,28. Los materiales utilizados en nuestro trabajo son ejemplos representativos de biomateriales sólidos y no degradables. Sin embargo, muchos otros materiales podrían ser utilizados con este modelo, siempre que el material se pueda recubrir en placas de cultivo celular o diapositivas de microscopio y descontaminado adecuadamente. Para este modelo in vitro se seleccionó la línea celular de macrófagodea NF-B/AP-1 porque permite una medición rápida e indirecta de la actividad NF-B/AP-1 a través de la expresión inducible NF-B/AP-1 del SEAP. Los macrófagos de reporteroNF-B/AP-1 requieren el uso de fleomicina D1 en los medios de cultivo como un antibiótico selectivo para garantizar que sólo las células con el gen SEAP inducible NF-B/AP-1 estén presentes29. Para el ensayo de fosfatasa alcalina, es fundamental utilizar HI-FBS en los medios de cultivo celular para evitar posibles resultados falsos positivos generados por fosfatasas alcalinas presentes en el suero. Nuestra investigación hasta la fecha sugiere que las superficies adsorvistas de FBS no generan un resultado falso positivo detectable, probablemente porque las moléculas séricas se absorben fuertemente a la superficie de cultivo y no se liberan en el sobrenadante. El punto de tiempo de cultivo para los macrófagos del reportero (20 h), el punto de tiempo de incubación del ensayo (2,5 h) y la longitud de onda de lectura de absorbancia (635 nm) para el ensayo de fosfatasa alcalina se optimizaron con este sistema para garantizar mediciones robustas y reproducibles para todas las condiciones.
Para este trabajo se eligió un plazo inicial de adsorción proteica de 30 min debido a su uso común en la literatura de adsorción proteica(Figura 1C)30,31,32,33,34. Sin embargo, también hemos explorado tiempos de adsorción más largos (es decir, 60 min y 24 h, Figura 4)para representar mejor la capa de proteína adsorbida con la que los macrófagos interactuarían in vivo, lo que es probable que ocurra de 4 a 24 h después de la implantación1. Se ha postulado que la mayoría de la adsorción e intercambio de proteínas se produce en los primeros 60 minutos de exposición a una superficie26,35,36, por lo tanto un tiempo de adsorción de 60 minutos puede ser un punto de tiempo más relevante. También hemos pasado de usar FBS como un control negativo para la presencia de DAPP en la capa de proteína adsorbida al plasma de ratón comercial. La razón para el uso de plasma en lugar de suero es que las proteínas plasmáticas son conocidas por desempeñar un papel significativo en la adsorción de proteínas y la respuesta a los macrófagos1, y que el plasma proporciona una mejor representación de las proteínas en el entorno de la herida. El plasma utilizado en experimentos de adsorción de proteínas se prepara comúnmente como una dilución del 1-10%26,36,37, lo que motivó nuestro uso de 10% plasma. Plasma humano se utiliza comúnmente26,36, ya que es más fácil de obtener en grandes cantidades y más clínicamente relevante, en comparación con el plasma del ratón. Sin embargo, elegimos usar plasma de ratón comercial para en este modelo para mantener las especies de las soluciones proteicas consistentes con las de las células del reportero.
El uso de la línea celular de los macrófagos del reportero introdujo algunas limitaciones dentro del estudio. En primer lugar, el uso de una línea celular de macrófago súbito murino tiene limitaciones inherentes, ya que el fenotipo y el comportamiento pueden variar de los cultivos de macrófagos primarios. Si bien esta limitación se abordará en futuros trabajos utilizando macrófagos primarios, se demostró que la línea celular de macrófagos parentales imitaba de cerca los macrófagos derivados de la médula ósea del ratón en términos de sus receptores de superficie celular y la respuesta a los ligandos microbianos para los TIR 2, 3 y 438. Además, los macrófagos de reportero NF-B/AP-1 dieron resultados similares en respuesta a la estimulación de HMGB1 y LPS en comparación con los macrófagos murinos primarios peritoneales39. Cabe señalar que los macrófagos del reportero NF-B/AP-1, y su cepa parental, no expresan TLR540. Los investigadores han demostrado que HMGB1 fue capaz de activar los factores de transcripción NF-B a través de las vías de señalización TLR5 en células HEK-293 estables infectadas con TLR541humano. Por lo tanto, en este modelo se descuidó la contribución de la señalización HMGB1-TLR5 a la actividad general nf-B en superficies recubiertas de lisado. Además, los macrófagos del reportero y su cepa parental no expresan la proteína adaptadora ASC, y por lo tanto no forman la mayoría de los tipos de inflamammasomes y no pueden procesar IL-1o inactivo o IL-18 inactivo a sus formas maduras42. Por lo tanto, el modelo que hemos utilizado no tiene en cuenta la contribución de la actividad inflammasome dependiente de ASC y la posterior señalización autocrina IL-1 y IL-18 en las respuestas de macrófagos a superficies de lisado-adsorbidos. En consecuencia, este ensayo está concebido como un examen preliminar de la activación NF-B dependiente del TLR, y se recomienda la investigación posterior utilizando macrófagos primarios para proporcionar una comprensión más completa y representativa de la activación de los macrófagos y el fenotipo en las superficies materiales de interés.
El ensayo de fosfatasa alcalina mide indirectamente la actividad NF-B/AP-1 de los macrófagos del reportero. Sin embargo, hay muchas vías de señalización distintas de los TIR que implican NF--B/AP-1 (por ejemplo, el receptor de interleucina-1 [IL-1R]43 y el receptor del factor de necrosis tumoral [TNFR]44). Por lo tanto, era necesario evaluar la contribución de la señalización TLR2 y TLR4 en la respuesta creciente de NF--B/AP-1 a superficies de lisado-adsorbida utilizando ensayos de inhibición(Figura 3). La razón para seleccionar estos dos TPR de superficie fue que al menos 23 DAPP que se han demostrado que señalan a través de TLR2 y TLR445,incluyendo el HMGB1 bien caracterizado, y ambos receptores se expresan en la superficie celular y pueden interactuar directamente con la superficie del biomaterial6. Los ensayos de inhibición TLR2 y TLR4 demostraron que cuando se bloquearon la señalización TLR2 o TLR4, se redujo la respuesta NF--B/AP-1 de los macrófagos del reportero al ispelado adsorbido, lo que indica que ambas vías están involucradas. Sin embargo, hubo una reducción notablemente mayor en la actividad de NF-B/AP-1 cuando se neutralizó la señalización TLR2, lo que sugiere que el TLR2 puede desempeñar un papel principal en la respuesta de los macrófagos de reportero según el lisado adsorbido. Reconocemos que puede haber alguna inhibición fuera del objetivo con la vía de señalización TLR neutralizando anticuerpos e inhibidores. Se utilizó un anticuerpo neutralizante para inhibir la vía TLR2 ya que no había moléculas inhibidoras TLR2 disponibles comercialmente en el momento de este trabajo.
Los métodos presentados aquí utilizan el issato, como una fuente compleja de DAPP, y los macrófagos de reporteroNF-B/AP-1 como modelo in vitro para las respuestas de macrófagos a los DAP y otras proteínas adsorbidas a biomateriales poliméricos(Figura 1). Anticipamos que nuestro protocolo se puede utilizar para analizar rápidamente las respuestas NF-B/AP-1 y la señalización TLR ascendente de los macrófagos de reporteros a una variedad de materiales (incluyendo materiales degradables, andamios porosos o hidrogeles) y capas de proteínas adsorbidas(Figura 3). Sin embargo, el uso de materiales porosos e hidrogeles introducirá complejidad dentro del sistema, ya que puede ser difícil distinguir entre moléculas adsorbidas y moléculas entrelazadas. También anticipamos que este protocolo se puede adaptar fácilmente para investigar la contribución de otras vías de señalización aguas arriba de NF--B/AP-1 (por ejemplo, receptores de lectina de tipo C46 y receptores similares a los de unión de nucleótidos (NOD)47) con los inhibidores adecuados. Además, la respuesta NF-B/AP-1 de los macrófagos de reporteros podría compararse entre diferentes materiales, siempre que las respuestas se normalizan a la actividad celular basal (es decir, células en medios en cada superficie sin proteína pre-adsorbido) y todos los materiales tienen niveles de endotoxina indetectables.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Los autores no tienen nada que revelar.
Acknowledgments
Los autores agradecen la financiación operativa de Canadian Institutes of Health Research Project (PTJ 162251), el Comité asesor de investigación del Senado de la Universidad de Queen y el apoyo a la infraestructura de la Fundación Canadiense para la Innovación del Fondo de Liderazgo john Evan (Proyecto 34137) y el Fondo de Investigación de Ontario del Ministerio de Investigación e Innovación (Proyecto 34137). L.A.M. fue apoyado por una beca R. Samuel McLaughlin de la Universidad de La Reina, un Consejo de Investigación de Ciencias Naturales e Ingeniería de Canadá Canadian Graduate Scholarship's Award y una Beca de Posgrado de Ontario. Los autores quieren agradecer al Dr. Myron Szewczuk por su generoso don de la línea celular de los macrófagos de reportero NF-B/AP-1 y a los doctores Michael Blennerhassett y Sandra Lourenssen por el uso de su sistema de imágenes de gel y lector de placas.
Materials
| Name | Company | Catalog Number | Comments |
| Cell culture reagents | |||
| anti-mouse/human CD282 (TLR2) | Biolegend | 121802 | |
| CLI-095 (TLR4 inhibitor) | Invivogen | TLRL-CLI95 | |
| C57 complement plasma K2 EDTA 10ml, innovative grade US origin | InnovativeResearch | IGMSC57-K2 EDTA-Compl-10ml | Mouse plasma |
| Dulbecco's modified eagle medium (DMEM) | Sigma Aldrich | D6429-500ML | |
| Dulbecco's phosphate buffered saline (DPBS) | Fisher Scientific | 14190250 | No calcium, no magnesium |
| Fetal bovine serum (FBS), research grade | Wisent | 98150 | |
| LPS-EK | Invivogen | TLRL-EKLPS | Lipopolysaccharide from Escherichia coli K12 |
| NIH/3T3 fibroblasts | ATCC | CRL-1658 | |
| Pam3CSK4 | Invivogen | tlrl-pms | Synthetic triacylated lipopeptide - TLR1/2 ligand |
| Penicillin/streptomycin | Sigma Aldrich | P4333-100ML | |
| Plasmocin | Invivogen | ANT-MPP | Mycoplasma elimination reagent |
| RAW-Blue cells | Invivogen | raw-sp | NF-κB/AP-1 reporter macrophage cell line |
| Trypan blue solution, 0.4% | Fisher Scientific | 15250061 | |
| TrypLE express enzyme (1X) | Fisher Scientific | 12604021 | animal origin-free recombinant cell dissociation enzyme |
| Zeocin | Invivogen | ANT-ZN-1 | |
| Kits and assays | |||
| ELISA precoated plates, mouse IL-6 | Biolegend | B213022 | |
| ELISA precoated plates, mouse TNF-α | Biolegend | B220233 | |
| Endotoxin (Escherichia coli) - Control standard endotoxin (CSE) | Associates of Cape Cope Inc. | E0005-5 | Endotoxin for standard curve in chromogenic endotoxin assay |
| LAL water, 100 mL | Associates of Cape Cope Inc. | WP1001 | Used with chromogenic endotoxin assay |
| Micro BCA protein assay | Fisher Scientific | PI23235 | |
| Limulus amebocyte lysate (LAL) Pyrochrome endotoxin test kit | Associates of Cape Cope Inc. | C1500-5 | Chromogenic endotoxin assay reagent |
| QUANTI-Blue alkaline phosphatase detection medium | Invivogen | rep-qb2 | Alkaline phosphatase assay to indirectly measure NF-κB/AP-1 activity |
| Polymeric coating reagents | |||
| Chloroform, anhydrous | Sigma Aldrich | 288306-1L | |
| Ethyl alcohol anhydrous | Commercial Alcohols | P006EAAN | Sigma: Reagent alcohol, anhydrous, 676829-1L |
| Straight tapered fine tip forceps | Fisher Scientific | 16-100-113 | |
| Fluorinert FC-40 solvent | Sigma Aldrich | F9755-100ML | Fluorinated solvent for fPTFE |
| Cell culture grade water (endotoxin-free) | Fisher Scientific | SH30529LS | |
| Poly(methyl methacrylate) (PMMA) | Sigma Aldrich | 182230-25G | |
| Sylgard 184 elastomer kit | Fisher Scientific | 50822180 | |
| Teflon-AF (fPTFE) | Sigma Aldrich | 469610-1G | Poly[4,5-difluoro-2,2-bis(trifluoromethyl)-1,3-dioxole-co-tetrafluoroethylene] |
| Consumables | |||
| Adhesive plate seals | Fisher Scientific | AB-0580 | |
| Axygen microtubes, 1.5 mL | Fisher Scientific | 14-222-155 | |
| Borosilicate glass scintillation vials, with white polypropylene caps | Fisher Scientific | 03-337-4 | |
| Clear PS 48-well plate | Fisher Scientific | 08-772-52 | |
| Clear TCPS 96-well plate | Fisher Scientific | 08-772-2C | |
| Clear TCPS 48-well plate | Fisher Scientific | 08-772-1C | |
| Cover glasses, circles | Fisher Scientific | 12-545-81 | |
| Falcon tissue culture treated flasks, T25 | Fisher Scientific | 10-126-10 | |
| sticky-Slide 8 Well | Ibidi | 80828 | |
| Superfrost microscope slides | Fisher Scientific | 12-550-15 | |
| Tissue culture treated flasks, T150 | Fisher Scientific | 08-772-48 |
References
- Anderson, J. M., Rodriguez, A., Chang, D. T. Foreign body reaction to biomaterials. Seminars in Immunology. 20 (2), 86-100 (2008).
- Anderson, J. M., Miller, K. M.
- Collier, T. O., Anderson, J. M. Protein and surface effects on monocyte and macrophage adhesion, maturation, and survival. Journal of Biomedical Materials Research. 60 (3), 487-496 (2002).
- Bianchi, M. E. DAMPs, PAMPs and alarmins: all we need to know about danger. Journal of Leukocyte Biology. 81 (1), 1-5 (2007).
- McKiel, L. A., Fitzpatrick, L. E. Toll-like Receptor 2-Dependent NF-κB/AP-1 Activation by Damage-Associated Molecular Patterns Adsorbed on Polymeric Surfaces. ACS Biomaterials Science & Engineering. 4 (11), 3792-3801 (2018).
- Babensee, J. E. Interaction of dendritic cells with biomaterials. Seminars in Immunology. 20 (2), 101-108 (2008).
- Sintes, J., Romero, X., de Salort, J., Terhorst, C., Engel, P. Mouse CD84 is a pan-leukocyte cell-surface molecule that modulates LPS-induced cytokine secretion by macrophages. Journal of Leukocyte Biology. 88 (4), 687-697 (2010).
- Tom, J. K., Mancini, R. J., Esser-Kahn, A. P. Covalent modification of cell surfaces with TLR agonists improves and directs immune stimulation. Chemical Communications. 49 (83), 9618-9620 (2013).
- Abdulkhalek, S., et al. Neu1 sialidase and matrix metalloproteinase-9 cross-talk is essential for toll-like receptor activation and cellular signaling. Journal of Biological Chemistry. 286 (42), 36532-36549 (2011).
- Gorbet, M. B., Sefton, M. V.
- Xing, Z., Pabst, M. J., Hasty, K. A., Smith, R. A. Accumulation of LPS by polyethylene particles decreases bone attachment to implants. Journal of Orthopaedic Research. 24 (5), 959-966 (2006).
- Ding, H., et al. Comparison of the cytotoxic and inflammatory responses of titanium particles with different methods for endotoxin removal in RAW264.7 macrophages. Journal of Materials Science: Materials in Medicine. 23 (4), 1055-1062 (2012).
- Hoogenboom, R., Becer, C. R., Guerrero-Sanchez, C., Hoeppener, S., Schubert, U. S. Solubility and thermoresponsiveness of PMMA in alcohol-water solvent mixtures. Australian Journal of Chemistry. 63 (8), 1173-1178 (2010).
- Efimenko, K., Wallace, W. E., Genzer, J. Surface modification of Sylgard-184 poly(dimethyl siloxane) networks by ultraviolet and ultraviolet/ozone treatment. Journal of Colloid and Interface Science. 254 (2), 306-315 (2002).
- Godek, M. L., Sampson, J. A., Duchsherer, N. L., McElwee, Q., Grainger, D. W. Rho GTPase protein expression and activation in murine monocytes/macrophages is not modulated by model biomaterial surfaces in serum-containing in vitro cultures. Journal of Biomaterials Science. Polymer Edition. 17 (10), 1141-1158 (2006).
- Park, J. S., et al. Involvement of Toll-like Receptors 2 and 4 in Cellular Activation by High Mobility Group Box 1 Protein. Journal of Biological Chemistry. 279 (9), 7370-7377 (2004).
- Ohashi, K., Burkart, V., Flohé, S., Kolb, H. Cutting Edge: Heat Shock Protein 60 Is a Putative Endogenous Ligand of the Toll-Like Receptor-4 Complex. The Journal of Immunology. 164 (2), 558-561 (2000).
- Wong, T., McGrath, J. A., Navsaria, H. The role of fibroblasts in tissue engineering and regeneration. British Journal of Dermatology. 156 (6), 1149-1155 (2007).
- van Wachem, P. B., et al. The influence of protein adsorption on interactions of cultured human endothelial cells with polymers. Journal of Biomedical Materials Research. 21 (6), 701-718 (1987).
- Miller, K. M., Anderson, J. M. Human monocyte/macrophage activation and interleukin 1 generation by biomedical polymers. Journal of Biomedical Materials Research. 22 (8), 713-731 (1988).
- Bonfield, T. L., Colton, E., Anderson, J. M. Plasma protein adsorbed biomedical polymers: Activation of human monocytes and induction of interleukin 1. Journal of Biomedical Materials Research. 23 (6), 535-548 (1989).
- González, O., Smith, R. L., Goodman, S. B. Effect of size, concentration, surface area, and volume of polymethylmethacrylate particles on human macrophages in vitro. Journal of Biomedical Materials Research. 30 (4), 463-473 (1996).
- Anderson, J. M., et al. Protein adsorption and macrophage activation on polydimethylsiloxane and silicone rubber. Journal of Biomaterials Science. Polymer Edition. 7 (2), 159-169 (1995).
- Lord, M. S., Foss, M., Besenbacher, F. Influence of nanoscale surface topography on protein adsorption and cellular response. Nano Today. 5 (1), 66-78 (2010).
- Chen, S., et al. Characterization of topographical effects on macrophage behavior in a foreign body response model. Biomaterials. 31 (13), 3479-3491 (2010).
- Shen, M., Horbett, T. A. The effects of surface chemistry and adsorbed proteins on monocyte/macrophage adhesion to chemically modified polystyrene surfaces. Journal of Biomedical Materials Research. 57 (3), 336-345 (2001).
- Love, R. J., Jones, K. S. The recognition of biomaterials: Pattern recognition of medical polymers and their adsorbed biomolecules. Journal of Biomedical Materials Research Part A. 101 (9), 2740-2752 (2013).
- McNally, A. K., Anderson, J. M. Phenotypic expression in human monocyte-derived interleukin-4-induced foreign body giant cells and macrophages in vitro: Dependence on material surface properties. Journal of Biomedical Materials Research Part A. 103 (4), 1380-1390 (2015).
- Gambhir, V., et al. The TLR2 agonists lipoteichoic acid and Pam3CSK4 induce greater pro-inflammatory responses than inactivated Mycobacterium butyricum. Cellular Immunology. 280 (1), 101-107 (2012).
- Suzuki, O., Yagishita, H., Yamazaki, M., Aoba, T. Adsorption of Bovine Serum Albumin onto Octacalcium Phosphate and its Hydrolyzates. Cells and Materials. 5 (1), 45-54 (1995).
- Johnston, R. L., Spalton, D. J., Hussain, A., Marshall, J. In vitro protein adsorption to 2 intraocular lens materials. Journal of Cataract and Refractive Surgery. 25 (8), 1109-1115 (1999).
- Jin, J., Jiang, W., Yin, J., Ji, X., Stagnaro, P. Plasma proteins adsorption mechanism on polyethylene-grafted poly(ethylene glycol) surface by quartz crystal microbalance with dissipation. Langmuir. 29 (22), 6624-6633 (2013).
- Swartzlander, M. D., et al. Linking the foreign body response and protein adsorption to PEG-based hydrogels using proteomics. Biomaterials. 41, 26-36 (2015).
- Chamberlain, M. D., et al. Unbiased phosphoproteomic method identifies the initial effects of a methacrylic acid copolymer on macrophages. Proceedings of the National Academy of Sciences. 112 (34), 10673-10678 (2015).
- Dillman, W. J., Miller, I. F. On the adsorption of serum proteins on polymer membrane surfaces. Journal of Colloid And Interface Science. 44 (2), 221-241 (1973).
- Ishihara, K., Ziats, N. P., Tierney, B. P., Nakabayashi, N., Anderson, J. M. Protein adsorption from human plasma is reduced on phospholipid polymers. Journal of Biomedical Materials Research. 25 (11), 1397-1407 (1991).
- Warkentin, P., Wälivaara, B., Lundström, I., Tengvall, P. Differential surface binding of albumin, immunoglobulin G and fibrinogen. Biomaterials. 15 (10), 786-795 (1994).
- Berghaus, L. J., et al. Innate immune responses of primary murine macrophage-lineage cells and RAW 264.7 cells to ligands of Toll-like receptors 2, 3, and 4. Comparative Immunology, Microbiology and Infectious Diseases. 33 (5), 443-454 (2010).
- Zhang, Y., Karki, R., Igwe, O. J. Toll-like receptor 4 signaling: A common pathway for interactions between prooxidants and extracellular disulfide high mobility group box 1 (HMGB1) protein-coupled activation. Biochemical Pharmacology. 98 (1), 132-143 (2015).
- Mizel, S. B., Honko, A. N., Moors, M. A., Smith, P. S., West, A. P. Induction of macrophage nitric oxide production by Gram-negative flagellin involves signaling via heteromeric Toll-like receptor 5/Toll-like receptor 4 complexes. Journal of Immunology. 170 (12), 6217-6223 (2003).
- Das, N., et al. HMGB1 Activates Proinflammatory Signaling via TLR5 Leading to Allodynia. Cell Reports. 17 (4), 1128-1140 (2016).
- Pelegrin, P., Barroso-Gutierrez, C., Surprenant, A. P2X7 Receptor Differentially Couples to Distinct Release Pathways for IL-1β in Mouse Macrophage. The Journal of Immunology. 180 (11), 7147-7157 (2008).
- Tak, P. P., Firestein, G. S. NF-κB: A key role in inflammatory diseases. Journal of Clinical Investigation. 107 (1), 7-11 (2001).
- Ashkenazi, A., Dixit, V. M. Death receptors: signaling and modulation. Science. 281 (5381), 1305-1308 (1998).
- Erridge, C. Endogenous ligands of TLR2 and TLR4: agonists or assistants. Journal of Leukocyte Biology. 87 (6), 989-999 (2010).
- Feng, Y., et al. A macrophage-activating, injectable hydrogel to sequester endogenous growth factors for in situ angiogenesis. Biomaterials. 134, 128-142 (2017).
- Lonez, C., et al. Cationic lipid nanocarriers activate Toll-like receptor 2 and NLRP3 inflammasome pathways. Nanomedicine: Nanotechnology, Biology, and Medicine. 10 (4), 775-782 (2014).
