Figure 1 presents a schematic for assembling the Glytube, whereas Figure 2 shows an overview of the experimental design to measure meal size using the fluorescence-based assay described here. Figure 3 provides representative fluorescein meal size measurements from a blood-feeding experiment. Figure 4, Figure 5, and Figure 6 illustrate a sampling of biological questions that can be addressed using this protocol. Applications of the protocol are wide-ranging and include altering blood meal composition, feeding pharmacological compounds, precisely quantifying sub-optimal blood meals or smaller nectar meals, and comparing feeding behavior across mosquito genotypes.
To generate a standard curve for meal volume calculations, fluorescence readings are plotted from the designated reference wells each containing an unfed mosquito and a known volume of the meal with 0.002% fluorescein (Figure 3A). Fluorescence readings from the remaining wells, which contain mosquitoes from either the negative control group of unfed mosquitoes or the experimental group of mosquitoes offered a meal, are compared to this standard curve to quantify the meal volume (µL) consumed by each mosquito (Figure 3B). To validate the baseline readings in this assay, it should be confirmed that mosquitoes from the unfed negative control group are not assigned a positive value of µL consumed (Figure 3B, left). Although all females in the experimental group were offered the blood meal, some mosquitoes fed (Figure 3B, middle) and some did not (Figure 3B, right). This result demonstrates that two types of data can be obtained from this protocol: 1) the percentage of total females that feed on a given meal, and 2) the volume ingested by the females that feed on a given meal.
This protocol can be used to deliver and quantify meals with various protein compositions. Figure 4A,B show data collected using meals with added fluorescein. The proportion of mosquitoes that fed and the meal volume they ingested, respectively, were calculated from the fluorescence readings. These readings are highly sensitive and allow for precise quantification of µL, but have the limitation that mosquitoes cannot be used for future live experiments. Figure 4C,D show data collected from an independent experiment with mosquitoes that were scored as fed or unfed by eye after they were offered meals without fluorescein. Meal size was calculated as average weight/female from groups of 5 mosquitoes. Although these weight measurements are less sensitive than fluorescence measurements, they allow the females to be recovered and used for further live experimentation. The proportion of mosquitoes that feed can vary across different experimental days, as reflected in Figure 4A and Figure 4C.
Figure 5 shows the volume consumed of meals containing drugs that regulate mosquito host-seeking behavior. In these experiments, females were offered blood, saline + ATP, or saline + ATP meals with 100 μM of the human NPY Y2 receptor agonist, TM30338. This drug alters host-seeking behavior through activation of Ae. aegypti NPY-like receptor 7. Measuring meal sizes is critical for the interpretation of experiments to assess the effect of this drug on post-blood-feeding behavior because it allows the researcher to calculate the dose consumed by each female.
In the previous examples, females were fed either blood or substitute blood meals, all of which resulted in 3–5 µL meals (Figure 3, Figure 4, Figure 5). This fluorescence-based assay can also be used to measure smaller and/or more variable meal sizes that cannot be accurately discerned from average group weight measurements. In Figure 6, the same fluorescence quantification protocol was used to measure nectar-feeding behavior by exchanging the Glytube for a cotton ball saturated with 10% sucrose containing 0.002% fluorescein. Nectar sugars cannot be presented in the Glytube assay because females cannot detect the presence of nectar sugars with the stylet and do not initiate feeding27. These data allow the researcher to determine that sugar meals are consistently smaller than blood meals, in agreement with previous work34 (Figure 6).
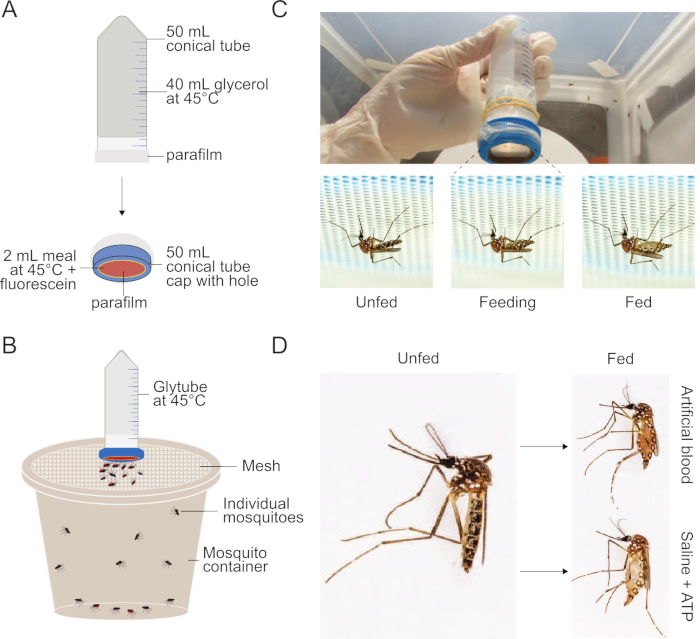
Figure 1: Setup of Glytube method used to feed meals to mosquitoes. (A) Schematic of a deconstructed Glytube used to feed blood and other meals to mosquitoes. (B) Schematic of a Glytube presented atop a container of mosquitoes with a mesh lid. Female mosquitoes can pierce through the mesh lid to feed. (C) Photographs of the Glytube (top), and female Aedes aegypti mosquitoes before, during, and after feeding (bottom, from left to right) on a Glytube-delivered meal. Mosquitoes are shown piercing through the mesh covering their container to access the membrane feeder. (D) Photographs showing the appearance of female Ae. aegypti mosquitoes that are unfed (left) and that have engorged on either an artificial blood meal (right, top) or a saline + ATP meal (right, bottom). The Glytube method was previously published in Costa-da-Silva et al. (2013)20. Photographs in (C) and (D) are courtesy of Alex Wild. Please click here to view a larger version of this figure.

Figure 2: Schematic of how to quantify meal size after Glytube blood-feeding protocol. (A) Mosquitoes are offered a meal with fluorescein (top, experimental group) or no meal (bottom, unfed negative control group). (B) Individual mosquitoes are added to a 96-well plate after terminating the feeding experiment. (C) Standard curve is generated using known amounts of meal containing 0.002% fluorescein. (D) Mosquitoes are homogenized to release any consumed fluorescein, and fluorescence levels in each well are quantified using a plate reader. This fluorescence quantification method is modified from Liesch et al. (2013)34. Please click here to view a larger version of this figure.
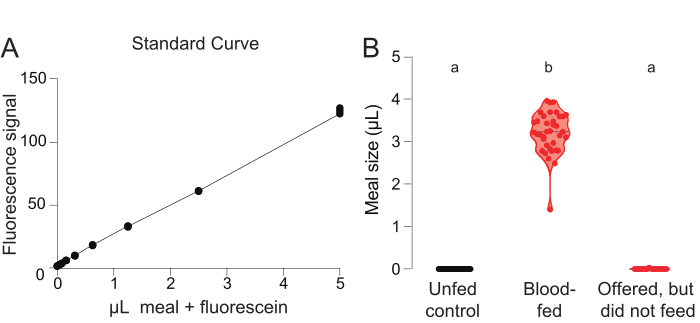
Figure 3: Glytube blood-feeding experiment with fluorescein-based quantification. (A) Standard curve measurements obtained from the wells where a mosquito from the unfed control group was added to a known quantity of meal containing 0.002% fluorescein (y-axis scale = arbitrary units). (B) Meal volume calculated using fluorescence readings for females in the unfed control group (left, black, n = 40), the experimental group that fed on blood (middle, red, n = 37), and the experimental group that did not feed on blood (right, red, n = 23). Each point represents a measurement from an individual female. Data are shown as median with range. Letters indicate statistically distinct groups, Kruskal-Wallis test with Dunn’s multiple comparison, p<0.01. These data were published in Jové et al. (2020)27. Please click here to view a larger version of this figure.
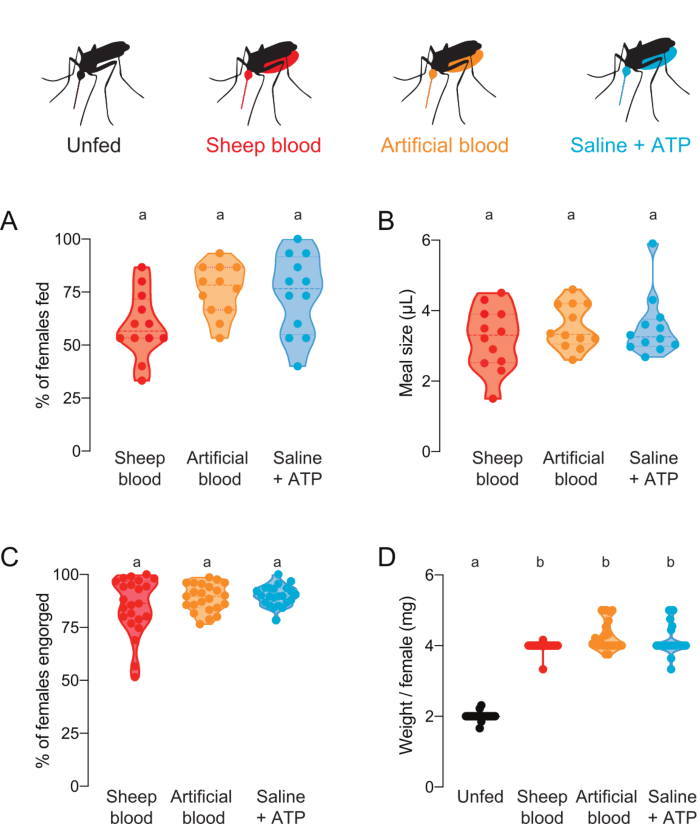
Figure 4: Quantification of meals with differing protein composition. Females were offered meals of either sheep blood (red), artificial blood with human blood proteins (Kogan (1990)22) (orange), or protein-free saline + ATP meal (aqua)7. (A) Percentage of females fed scored using fluorescence readings. Each point represents a group of 12–16 females. Data are shown as medians with ranges, n = 12. (B) Meal volume calculated using fluorescence readings. Each point represents a measurement from an individual female in a single trial from Figure 4A. Data are shown as medians with ranges, n = 12. (C) Percentage of females fully engorged after artificial membrane feeding, scored by eye. Each point represents the percent of females engorged from groups of 20–30 females. Data are shown as medians with ranges, n = 23. (D) Meal sizes scored as weight/female after feeding status was scored by eye. Weights were calculated as the average of groups of 5 mosquitoes. Data are shown as medians with ranges, n = 23. A–D: Letters indicate statistically distinct groups, Kruskal-Wallis test with Dunn’s multiple comparison, p<0.05. Please click here to view a larger version of this figure.
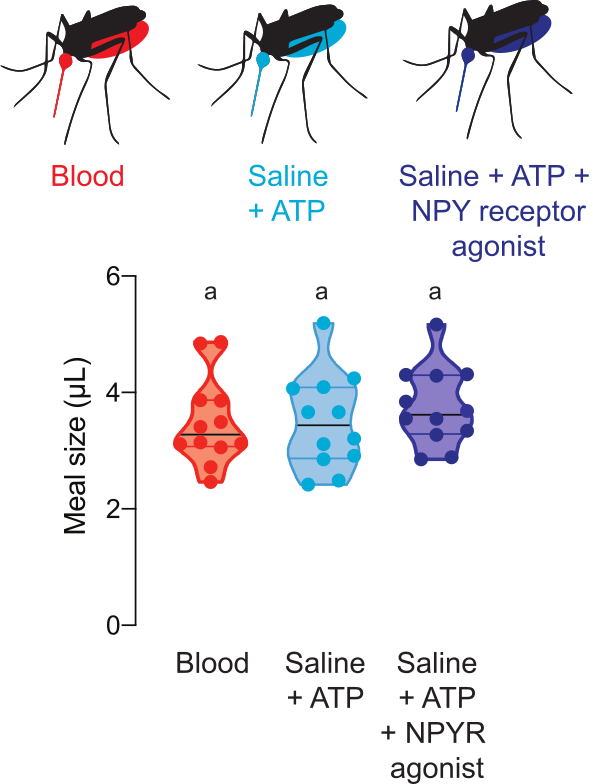
Figure 5: Quantification of meals with pharmacological compounds. Females consume meals of the same size of sheep blood (red), saline + ATP (aqua), and saline + ATP + 100 µM dose of human NPY Y2 receptor agonist TM30338 (dark blue). Meal volume calculated using fluorescence readings. Each point represents a measurement from an individual female. Data are shown as medians with ranges, n = 12. Letters indicate statistically distinct groups, Kruskal-Wallis test with Dunn’s multiple comparison, p<0.05. Please click here to view a larger version of this figure.

Figure 6: Quantification of smaller nectar meals. (A) Schematic of nectar-feeding assay. (B) Meal volume calculated using fluorescence readings for wild-type females offered meals of either water (blue, n = 36) or 10% sucrose (green, n = 53), each with 0.002% fluorescein, in the nectar-feeding assay. Each point represents a measurement from an individual female. Data are shown as medians with ranges. Letters indicate statistically distinct groups, Mann-Whitney test, p<0.05. These data were published in Jové et al. (2020)27. Please click here to view a larger version of this figure.
| Artificial Blood Meal | |||
| Concentration of Stock Solution (mg/mL) | Volume of Stock Solution in Meal (μL/mL) | Final Meal Concentration (mg/mL) | |
| Protein Components* | |||
| γ-Globulins | 50 | 300 | 15 |
| Hemoglobin | 35 | 230 | 8 |
| Albumin | 300 | 340 | 102 |
| Total Protein | – | – | 125 |
| Non-Protein Components | |||
| Concentration of Stock Solution (mM) | Volume of Stock Solution in Meal (μL/mL) | Final Meal Concentration (mM) | |
| NaCl | In γ-globulin stock | – | 5-10 |
| NaHCO3 | In γ-globulin stock | – | 120 |
| ATP | 200 | 5 | 1 |
| Water | – | 125 | – |
| *Protein components are prepared in stock solution of double-distilled water, except for γ-Globulins, which are dissolved in 400 mM NaHCO3 and include a variable amount of NaCl (2-4%) in the product. | |||
Table 1: Recipe for preparing artificial blood meals (adapted from Kogan (1990)22). Artificial blood consists of protein and non-protein components regularly found in human blood and provides the option to vary the ratios of these components. Mosquitoes can produce eggs after feeding on artificial blood7,22.
| Saline Meal | |||
| Component | Concentration of Stock Solution (mM) | Volume of Stock Solution in Meal (μL/mL) | Final Meal Concentration (mM) |
| NaCl | – | – | – |
| NaHCO3 | 400 | 300 | 120 |
| ATP | 200 | 5 | 1 |
| Water | – | 695 | – |
Table 2: Recipe for saline meal with ATP (adapted from Duvall et al. (2019)7). Protein-free saline meals can be used to deliver compounds of interest to mosquitoes while still mimicking the abdominal distension that occurs after blood-feeding, but without triggering the egg development that occurs when proteins are ingested.
