

Summary
Recycling endosomen maken deel uit van het endosomale buisvormige netwerk. Hier presenteren we een methode om de dynamiek van recycling-endosomen te kwantificeren met behulp van GFP-STX13 als organelmarker.
Abstract
Recycling-endosomen (RE's) zijn tubulair-vesiculaire organellen die worden gegenereerd uit vroege / sorterende endosomen in alle celtypen. Deze organellen spelen een sleutelrol in de biogenese van melanosomen, een lysosoom-gerelateerd organel geproduceerd door melanocyten. RE's leveren de melanocyt-specifieke lading aan premature melanosomen tijdens hun vorming. Blokkering bij het genereren van RE's, waargenomen bij verschillende mutanten van het Hermansky-Pudlak-syndroom, resulteert in hypopigmentatie van huid, haar en ogen. Daarom is het bestuderen van de dynamiek (zie aantal en lengte) van RE's nuttig om de functie van deze organellen in normale en ziekteomstandigheden te begrijpen. In deze studie willen we de RE-dynamiek meten met behulp van een resident SNARE STX13.
Introduction
Biosynthese van melaninepigmenten vindt plaats in melanosomen, een melanocyt-specifiek lysosoom-gerelateerd organel (LRO) dat naast conventionele lysosomen bestaat. Het endocytische systeem speelt een sleutelrol in de biogenese van melanosomen, die nodig zijn voor huidskleur en fotoprotectie tegen ioniserende straling1,2,3. Tijdens dit proces worden de melaninesynthetiserende enzymen gesorteerd op vroege / sorteer-endosomen en vervolgens getransporteerd naar premature melanosomen via tubulaire of vesiculaire endosomen die recycling-endosomen (RE's) worden genoemd 4,5,6,7,7,8,9,10. De targeting en fusie van deze organellen reguleren de rijping van volledig functionele gepigmenteerde melanosomen7,11,12,13,14. Defecten in de vorming van deze organellen of ladingssortering naar deze organellen veroorzaken oculocutaan albinisme en andere klinische fenotypen, waargenomen bij Hermansky-Pudlak-syndroom15,16.
Hier beschrijven we een eenvoudige op microscopie gebaseerde techniek om de RE's te bestuderen en te analyseren. Bij deze methode hebben we gebruik gemaakt van een transmembraaneiwit, Qa-SNARE Syntaxin (STX)13, dat zich bevindt op het recyclen van endosomen17 en cycli tussen het sorteren van endosomen en melanosomen in melanocyten12,18. Verder zorgt het verwijderen van N-terminaal ongestructureerd regulerend domein (namelijk SynN of STX13Δ129) ervoor dat de SNARE vast komt te zitten in melanosomen, die de voorwaartse smokkelroute naar het melanosoom meet12. We hebben een bekende recycling endosomale marker Rab GTPase (Rab)11 gebruikt in onze studies14,19. Fluorescentiebeeldvorming van de eiwitten GFP-STX13WT, GFP-STX13Δ129, mCherry-Rab11 en TYRP1 in melanocyten van het wilde type, gevolgd door kwantificering van hun relatieve lokalisatie, zal de aard en dynamiek van RE's bieden naast hun targeting op melanosomen. Dit is dus een eenvoudige techniek die kan worden gebruikt om de dynamiek van RE's in melanocyten te visualiseren en te meten.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
Het protocol omvat het zaaien van melanocyten gevolgd door transfectie van de plasmiden. Verdere stappen omvatten fixatie, immunostaining, beeldvorming en analyse van de cellen om de lengte en het aantal RE's te meten. De gedetailleerde beschrijving van het protocol wordt hieronder gegeven.
1. Zaaien van muizenmelanocyten op voorbehandelde coverslips
- Bestrijk de glazen afdekplaten in een petrischaaltje (d.w.z. 4 - 5 in een schaal van 35 mm) met keldermembraanmatrixmedium (1:20 in volledig RPMI-medium: RPMI + 10% warmte-geïnactiveerd FBS + 1x Glutamine + 1x Antibioticamengsel) en droog het gedurende 15 minuten in de weefselkweekkap. Was de coverslips voor gebruik één keer met 1x PBS.
- Bewaar de wild type muismelanocyten (melan-Ink4a-Arf-1 van C57BL/6J, a/a, Ink4a-Arf-/- muizen, beschreven in 20 en verkrijgbaar bij The Welcome Trust Functional Genomics Cell Bank) in een petrischaaltje (d.w.z. 35 of 60 mm schaal) aangevuld met volledig RPMI-medium.
- Was de cellen tweemaal met 1x PBS en voeg 0,5 of 1 ml Trypsine-EDTA (0,25%) oplossing voor trypsiniserende cellen toe. Incubeer de cellen bij 37 °C gedurende 2 - 5 minuten voor het losmaken van de petrischaal.
- Voeg 1 - 2 ml volledig RPMI-medium toe, suspensie en breng de cellen vervolgens over naar een centrifugebuis.
- Centrifugeer de celsuspensie bij 4 °C, 376 x g gedurende 5 minuten en resuspend de pellet vervolgens in 1x PBS.
- Herhaal de centrifugatiestap en resuspend de cellen in 1 ml volledig RPMI-medium.
- Zaai de cellen op het keldermembraan medium-gecoate coverslips met 50-60% confluentie (ongeveer 6 x 105 cellen op een 35 mm celkweekschaal met 4 - 5 coverslips). Voeg altijd 200 nM PMA (voeg 5 μL werkvoorraad 40 μM phorbol 12-myristaat 13-acetaat toe) toe aan de vergulde celsuspensie in volledig RPMI-medium.
- Incubeer na het zaaien de plaat bij 37 °C gedurende 12- 24 uur.
2. Transfectie van cellen met de STX13 plasmiden
- Gebruik de volgende reagentia: pEGFP-C1-STX13WT en pEGFP-C1-STX13Δ129 (beschreven in12). mCherry-Rab11, was een vriendelijk geschenk van Graça Raposo, Institut Curie, Parijs (beschreven in referentie19). Anti-TYRP1 antilichaam van ATCC (TA99).
- Na 12 - 24 uur zaaien, transfecteer de cellen met plasmiden met behulp van een op lipiden gebaseerd transfectiereagens. Neem voor een schotel van 35 mm 5 μL van het transfectiereagens in 250 μL OPTI-MEM-medium in een microcentrifugebuis en neem ongeveer 200 ng van elk plasmide in 250 μL OPTI-MEM.
- Incubeer de buisjes met DNA en het transfectiereagens gedurende 5 minuten. Meng zonder herhaald pipetteren (totaal volume zal ongeveer 500 μL zijn). Incubeer gedurende 30 minuten bij kamertemperatuur. Voer elke 10 minuten gedurende 30 minuten met de hand tikken op de buis uit bij RT.
- Was de cellen tijdens de incubatie twee keer met 1x PBS, één keer met OPTI-MEM en voeg vervolgens 1 ml OPTI-MEM toe aan de cellen.
- Voeg na 30 minuten incubatie het transfectiereagens-DNA-mengsel druppelsgewijs toe aan de cellen door de schaal te bedekken.
- Incubeer de cellen bij 37 °C gedurende 6 uur. Zuig het OPTI-MEM medium aan met transfectiereagens en voeg het volledige RPMI medium aangevuld met 200 nM PMA toe.
- Incubeer de cellen bij 37 °C gedurende 48 uur.
3. Fixatie van de cellen
OPMERKING: De volgende procedure wordt uitgevoerd buiten de weefselkweekkap.
- Na 48 uur transfectie, was de cellen tweemaal met 1x PBS en fixeer de cellen vervolgens met 3% formaldehyde (vers bereid in 1x PBS) gedurende 30 minuten.
- Was de cellen na fixatie twee keer met 1x PBS en bewaar de coverslips in 1x PBS tot verder gebruik. Als alternatief kunnen cellen op glazen platen worden gemonteerd (zie hieronder) of worden opgeslagen bij 4 °C.
4. Immunostaining van de cellen
- Bereid een vochtige kamer voor: plaats paraffinefilm gesneden stuk op vochtig filterpapier in een petrischaal, bedekt met aluminiumfolie.
- Bereid 25 μL primaire antilichaamoplossing (0,2% saponine in 1x PBS, 0,1% BSA in 1x PBS en 0,02% natriumazide in 1x PBS). Voeg antilichaam toe bij een verdunning van 1:200. Voeg deze oplossing toe als druppel op een paraffinefilm in de vochtige kamer.
- Til de deklip voorzichtig op met een tang, keer deze om op deze druppel primaire antilichaamkleuringsoplossing en bedek vervolgens het deksel van de vochtige kamer. Incubeer bij kamertemperatuur gedurende 30 min.
- Bereid op dezelfde manier de secundaire antilichaamoplossing met een verdunning van 1:500 en plaats deze op paraffinefilm naast de dekplaat in de vochtige kamer. Voeg voor het kleuren van de kern DAPI (1:20.000 tot 1:30.000) toe aan de oplossing.
- Pak met behulp van een tang voorzichtig de deklip van de primaire antilichaamoplossing en dompel deze driemaal in 1x PBS (in een glazen bekerglas).
- Tik op de coverslip op tissuepapier om de overtollige PBS op de coverslip te verwijderen. Plaats het op secundaire antilichaamkleuringsoplossing in de vochtige kamer en stel het niet bloot aan licht vanwege de aanwezigheid van fluorescerend gelabelde antilichamen in de oplossing.
- Incubeer de coverslip opnieuw gedurende 30 minuten bij kamertemperatuur. Houd altijd een notitie aan de zijkant van de coverslip die de cellen tijdens deze stappen heeft.
- Pak na de incubatie voorzichtig de coverslip van de secundaire antilichaamoplossing en dompel deze vervolgens driemaal in 1x PBS. Tik verder op de coverslip op tissuepapier om de overtollige PBS op de coverslip te verwijderen.
- Plaats 12 μL Fluoromount-G montagereagens op een glasplaat en plaats voorzichtig de gebrandschilderde afdekkingslip (naar het glas gericht) op het montagereagens. Keer de glasplaat om op tissuepapier en druk vervolgens zachtjes aan.
5. Fluorescentiemicroscopie van de cellen
- Stel de gekleurde cellen onder Bright-field (BF) en fluorescentie (IF) filters in beeld met behulp van een omgekeerde fluorescentiemicroscoop uitgerust met een CCD-camera met behulp van 60x (olie) apochromatische objectieven of een andere microscoop met een vergelijkbare configuratie.
6. Kwantificering van de overlap tussen de RE gelokaliseerde eiwitten en melanosomen:
OPMERKING: De volgende stappen worden gevolgd voor de kwantificering van mander's overlapcoëfficiënt tussen de eiwitten met behulp van Fiji software (vrij te downloaden via de link: https://imagej.net/software/fiji/). Gebruik de TIFF-afbeelding met meerdere kanalen.
- Open de RAW-afbeelding. Ga naar de optie Afbeelding en selecteer Kleur | Splits kanalen en gebruik de twee kanalen voor analyse.
- Open de JACoP-plug-in in de optie Plug-in.
- Stel de drempel voor beide kanalen zo in dat alle lichtpuntjes worden geselecteerd en de achtergrond wordt geëlimineerd.
- Ga naar de optie Analyse, selecteer M1- en M2-coëfficiënten om de overlapcoëfficiënt van Mander te krijgen.
- Druk op de optie Analyseren in de JACoP-plug-in en zie het resultaat dat de overlapcoëfficiënt van Mander weergeeft.
7. Kwantificering van het buisvormige aantal en de lengte van recycling-endosomen:
OPMERKING: De volgende stappen worden gevolgd voor de kwantificering van het aantal en de lengte van de tubuli met behulp van Fiji-software.
- Open de raw-afbeelding Ga naar afbeelding optie, selecteer Kleur | Splits kanalen en gebruik het gewenste kanaal voor analyse.
- Ga nogmaals naar Afbeelding , selecteer Type | Converteren naar 8-bits afbeelding.
- Ga vervolgens naar Plug-ins, selecteer | analyseren Tubeness. Stel sigmawaarde in op 0,1075 . Druk op OK.
- Ga nogmaals naar Afbeelding , selecteer Type | Converteren naar 8 bit afbeelding.
- Ga naar Afbeelding | | aanpassen Drempelwaarde (Gebruik dezelfde drempelwaarden voor alle afbeeldingen. Afbeeldingen moeten ongeveer dezelfde intensiteit hebben).
- Ga naar proces- | Binaire | Converteren naar masker.
- Ga naar Proces, selecteer Binair en selecteer vervolgens Skeletonize.
- Ga naar Analyseren, selecteer Skelet en kies Skelet analyseren. In resultaat en uitvoer selecteert | (a) Bereken het grootste kortste pad | (b) toon gedetailleerde informatie | (c) geef het gelabelde skelet weer. Druk op OK.
OPMERKING: Het resultaat wordt geopend in de tabelindeling. De kolom Gemiddelde taklengte toont de lengte van alle verschillende buisjes in de geselecteerde cel (stel schaal in voor het verkrijgen van waarden in micrometers). - Als u het aantal buisjes in de cel wilt verkrijgen, gaat u naar Analyseren en selecteert u de optie Deeltje analyseren . Druk op OK.
OPMERKING: In de verkregen resultaten toont de telkolom het aantal tubuli in die specifieke cel. - Sla de gegevens op en analyseer.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Kwantificering van STX13Δ129 mutante lokalisatie naar de melanosomen
Immunofluorescentiemicroscopie van STX13 in muiswildtype melanocyten toonde GFP-STX13WT gelokaliseerd als vesiculaire en buisvormige structuren en GFP-STX13Δ129 gelokaliseerd als ringachtige structuren naast het celoppervlak (figuur 1A). Verder toonde intracellulair ringachtig GFP-STX13Δ129 colocalisatie met het melanosoomeiwit TYRP1 (figuur 1A) en helderveldbeeldige melanosomen (gegevens niet getoond)12. Zoals eerder aangetoond, wordt een cohort van overexpressie GFP-STX13WT waargenomen in melanosomen12. Om de relatieve lokalisatie van GFP-STX13WT en GFP-STX13Δ129 naar melanosomen te meten, hebben we Fiji gebruikt en geanalyseerd met jacop plugin. De gemeten Mander's overlap coëfficiënt (MOC) tussen GFP-STX13Δ129 met TYRP1 is ongeveer 1,5 keer hoger in vergelijking met GFP-STX13WT met TYRP1 (Figuur 1B). Interessant is dat TYRP1 2,9 keer hogere MOC-waarden vertoonde met GFP-STX13Δ129 in vergelijking met GFP-STX13WT (figuur 1B). Deze gegevens geven aan dat de lokalisatie van GFP-STX13Δ129 naar melanosomen relatief hoger is in vergelijking met GFP-STX13WT bij een steady state.
Kwantificering van STX13WT-lokalisatie naar de recycling-endosomen
Immunofluorescentiemicroscopie van GFP-STX13WT toonde colocalisatie met bekende recycling endosomale proteïne Rab11 (uitgedrukt als mCherry-Rab11) (figuur 2A,B). De gemeten MOC tussen GFP-STX13WT en mCherry-Rab11 is ongeveer 1,4 keer hoger in vergelijking met mCherry-Rab11 met GFP-STX13WT (figuur 2B). Om het aantal en de lengte van GFP-STX13WT-positieve endosomale tubuli te meten, hebben we Fiji-software gebruikt zoals beschreven in de protocolsectie. mCherry-Rab11 wordt gebruikt als positieve controle in de experimenten (figuur 2). Melanocyten getransfecteerd met GFP-STX13WT vertoonden een hoger aantal tubuli per cel in vergelijking met cellen die mCherry-Rab11 tot expressie brengen (Figuur 2C bovenste grafiek, vergelijk staaf A met staaf B). De tubulusgetallen worden echter verminderd bij co-expressie van GFP-STX13WT en mCherry-Rab11 in de cellen (figuur 2C bovenste grafiek, vergelijk staaf A met C en balk B met D). Interessant is dat de gemiddelde tubuluslengte (μm) voor zowel GFP-STX13WT als mCherry-Rab11 vergelijkbaar is met elkaar in cellen die afzonderlijk of samen tot expressie komen (figuur 2C onderste grafiek). Samen suggereren deze gegevens dat GFP-STX13WT lokaliseert naar RET's als vergelijkbaar met Rab11.
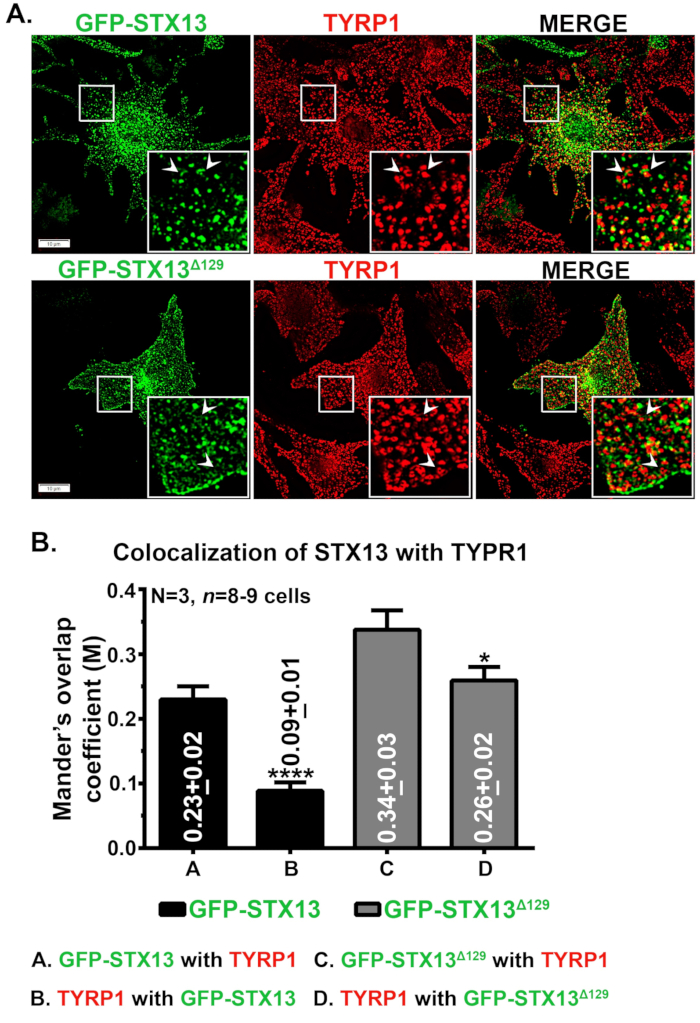
Figuur 1: Lokalisatie van GFP-STX13WT en GFP-STX13Δ129 naar melanosomen in melanocyten van het wilde type. (A) Melan-Ink4a-Arf-1 melanocyten werden getransfecteerd met GFP-STX13WT en GFP-STX13Δ129. Cellen werden gefixeerd, gekleurd met anti-TYRP1-antilichaam en vervolgens geanalyseerd door fluorescentiemicroscopie. Inzetstukken zijn een vergrote weergave van de witte kadergebieden. Pijlen wijzen naar de lokalisatie van GFP-STX13WTto RE's en GFP-STX13Δ129 naar melanosomen. Schaalbalken, 10 μm. (B) Kwantificering van colocalisatie tussen STX13 en TYRP1. De overlapcoëfficiënt van Mander (M) tussen GFP-STX13WT of GFP-STX13Δ129 met TYRP1 en vice versa wordt afzonderlijk in de plot weergegeven (gemiddelde ± S.E.M.). N=3. *p ≤0,05 en ****p ≤0,0001. Klik hier om een grotere versie van deze figuur te bekijken.

Figuur 2: Lokalisatie van GFP-STX13WT naar recycling van endosomen in melanocyten van het wilde type. (A) Melan-Ink4a-Arf-1-melanocyten werden getransfecteerd met GFP-STX13WT en mCherry-Rab11. Cellen werden gefixeerd en geanalyseerd door fluorescentiemicroscopie. Inzetstukken zijn een vergrote weergave van de witte kadergebieden. Pijlen wijzen naar de lokalisatie van GFP-STX13WT naar mCherry-Rab11-positieve compartimenten. Schaalbalken, 10 μm. (B) Kwantificering van colocalisatie tussen STX13 en Rab11. Manders overlapcoëfficiënt (M) tussen GFP-STX13WT met mCherry-Rab11 en vice versa wordt (gemiddelde ± S.E.M.) afzonderlijk in de plot weergegeven. N=3. p ≤0,001. C. Kwantificering van aantal en lengte (in μm) van STX13- of Rab11-positieve RE's. Het gemiddelde aantal tubuli/cel en de gemiddelde tubuluslengte van GFP-STX13WT en mCherry-Rab11 worden (gemiddelde ± S.E.M.) afzonderlijk in het waarnemingspunt weergegeven. N=3. Merk op dat de cellen zijn getransfecteerd met GFP-STX13WT en mCherry-Rab11 in (C) en (D). Klik hier om een grotere versie van deze figuur te bekijken.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Recycling-endosomen zijn een cohort van endocytische organellen en ze bemiddelen de recycling van lading naar het celoppervlak in alle celtypen21,22,23,24,25. In gespecialiseerde celtypen zoals melanocyten leiden deze organellen hun smokkelroute gedeeltelijk om naar de melanosomen voor hun biogenese3,16,26. Verder worden RET's gegenereerd na het sorteren van lading op vroege / sorteerdosomen en morfologisch verschijnen als buisvormige vesiculaire structuren. Hun dynamiek (aantal en lengte) is afhankelijk van verschillende cellulaire factoren, waaronder motoreiwitten21,22,23,24,25,27,28. Weinig studies gebruikten Rab11 als marker om de RE's in melanocyten te labelen14,19. Terwijl in niet-melanocyten naast Rab11 ook STX13 en KIF13A zijn gebruikt om ES13,17,19 te visualiseren en te karakteriseren. In deze studie hebben we STX13 gebruikt als marker om de RE's in melanocyten te labelen, gevolgd door een vergelijking met Rab11-positieve tubuli (figuur 2). Interessant is dat N-terminale deletie in STX13 (GFP-STX13Δ129) resulteert in een verkeerde lokalisatie van de SNARE naar melanosomen (figuur 1). Zo visualiseert de lokalisatie van GFP-STX13WT en GFP-STX13Δ129 in melanocyten respectievelijk de RE's en melanosomen. Deze eiwitten kunnen worden gebruikt als respectievelijke organellenmarkers voor de steady-state lokalisatiestudies.
Studies hebben aangetoond dat verschillende eiwitten zoals TYRP1, TYR (tyrosinase), VAMP7, OCA2, Rab32/38 lokaliseren naar de melanosomen naast andere intracellulaire organellen3,29. Onze studies toonden aan dat de N-terminale verwijderde STX13-mutant (GFP-STX13Δ129) lokaliseert naar melanosomen en plasmamembraan. We veronderstellen dat GFP-STX13Δ129 mogelijk kan worden gebruikt als een verslaggever om de handel van PE's naar het celoppervlak en LRO te bestuderen. GFP-STX13WT daarentegen lokaliseert naar RET's als vergelijkbaar met Rab11. Onze studies illustreerden dat GFP-STX13WT ook kan worden gebruikt voor het markeren van RE's in melanocyten. We voorspellen dat GFP-STX13WT een betere RE-marker kan zijn dan Rab11, omdat Rab11-overexpressie de endosomale dynamiek verandert. Al met al fungeert GFP-STX13WT als een potentiële RE-marker om hun dynamiek bij steady-state conditie te bestuderen.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
De auteurs verklaren geen belangenverstrengeling te hebben.
Acknowledgments
Dit werk werd ondersteund door het Department of Biotechnology (BT/PR32489/BRB/10/1786/2019 aan SRGS); Onderzoeksraad voor Wetenschap en Techniek (CRG/2019/000281 SRGS); DBT-NBACD (BT/HRD-NBA-NWB/38/2019-20 naar SRGS) en IISc-DBT partnerschapsprogramma (naar SRGS). De infrastructuur op de afdeling werd ondersteund door DST-FIST, DBT en UGC. AMB werd ondersteund door DBT-JRF (DBT/2015/IISc/NJ-02).
Materials
| Name | Company | Catalog Number | Comments |
| anti-TYRP1 antibody (TA99) | ATCC | HB-8704 | |
| Fluoromount-G | Southern Biotech | 0100-01 | |
| Lipofectamine 2000 | ThermoFisher Scientific | 11668-500 | |
| Matrigel matrix | BD Biosciences | 356231 | |
| OPTI-MEM | ThermoFisher Scientific | 022600-050 | |
| Phorbol 12-myristate 13-acetate | Sigma-Aldrich | P8139 | |
| RPMI Medium 1640 | ThermoFisher Scientific | 31800-022 |
References
- Dell'Angelica, E. C. The building BLOC(k)s of lysosomes and related organelles. Current Opinion in Cell Biology. 16 (4), 458-464 (2004).
- Raposo, G., Marks, M. S. Melanosomes--dark organelles enlighten endosomal membrane transport. Nature Reviews in Molecular Cell Biology. 8 (10), 786-797 (2007).
- Ohbayashi, N., Fukuda, M. Recent advances in understanding the molecular basis of melanogenesis in melanocytes. F1000Research. 9, F1000 Faculty Rev-608 (2020).
- Theos, A. C., et al. Functions of adaptor protein (AP)-3 and AP-1 in tyrosinase sorting from endosomes to melanosomes. Molecular Biology of the Cell. 16 (11), 5356-5372 (2005).
- Di Pietro, S. M., et al. BLOC-1 interacts with BLOC-2 and the AP-3 complex to facilitate protein trafficking on endosomes. Molecular Biology of the Cell. 17 (9), 4027-4038 (2006).
- Setty, S. R., et al. BLOC-1 is required for cargo-specific sorting from vacuolar early endosomes toward lysosome-related organelles. Molecular Biology of the Cell. 18 (3), 768-780 (2007).
- Delevoye, C., et al. AP-1 and KIF13A coordinate endosomal sorting and positioning during melanosome biogenesis. Journal of Cell Biology. 187 (2), 247-264 (2009).
- Bultema, J. J., Ambrosio, A. L., Burek, C. L., Di Pietro, S. M. BLOC-2, AP-3, and AP-1 proteins function in concert with Rab38 and Rab32 proteins to mediate protein trafficking to lysosome-related organelles. Journal of Biological Chemistry. 287 (23), 19550-19563 (2012).
- Sitaram, A., et al. Differential recognition of a dileucine-based sorting signal by AP-1 and AP-3 reveals a requirement for both BLOC-1 and AP-3 in delivery of OCA2 to melanosomes. Molecular Biology of the Cell. 23 (16), 3178-3192 (2012).
- Nag, S., et al. Rab4A organizes endosomal domains for sorting cargo to lysosome-related organelles. Journal of Cell Science. 131 (18), (2018).
- Dennis, M. K., et al. BLOC-2 targets recycling endosomal tubules to melanosomes for cargo delivery. Journal of Cell Biology. 209 (4), 563-577 (2015).
- Jani, R. A., Purushothaman, L. K., Rani, S., Bergam, P., Setty, S. R. STX13 regulates cargo delivery from recycling endosomes during melanosome biogenesis. Journal Cell Science. 128 (17), 3263-3276 (2015).
- Shakya, S., et al. Rab22A recruits BLOC-1 and BLOC-2 to promote the biogenesis of recycling endosomes. EMBO Reports. 19 (12), 45918 (2018).
- Bowman, S. L., et al. A BLOC-1-AP-3 super-complex sorts a cis-SNARE complex into endosome-derived tubular transport carriers. Journal of Cell Biology. 220 (7), 202005173 (2021).
- Wei, M. L. Hermansky-Pudlak syndrome: a disease of protein trafficking and organelle function. Pigment Cell Research. 19 (1), 19-42 (2006).
- Bowman, S. L., Bi-Karchin, J., Le, L., Marks, M. S. The road to lysosome-related organelles: Insights from Hermansky-Pudlak syndrome and other rare diseases. Traffic. 20 (6), 404-435 (2019).
- Prekeris, R., Klumperman, J., Chen, Y. A., Scheller, R. H. Syntaxin 13 mediates cycling of plasma membrane proteins via tubulovesicular recycling endosomes. Journal of Cell Biology. 143 (4), 957-971 (1998).
- Mahanty, S., et al. Rab9A is required for delivery of cargo from recycling endosomes to melanosomes. Pigment Cell Melanoma Research. 29 (1), 43-59 (2016).
- Delevoye, C., et al. Recycling endosome tubule morphogenesis from sorting endosomes requires the kinesin motor KIF13A. Cell Reports. 6 (3), 445-454 (2014).
- Ha, L., et al. ARF functions as a melanoma tumor suppressor by inducing p53-independent senescence. Proceedings of the National Academy of Science U. S. A. 104 (26), 10968-10973 (2007).
- Soldati, T., Schliwa, M. Powering membrane traffic in endocytosis and recycling. Nature Reviews Molecular Cell Biology. 7 (12), 897-908 (2006).
- Grant, B. D., Donaldson, J. G. Pathways and mechanisms of endocytic recycling. Nature Reviews Molecular Cell Biology. 10 (9), 597-608 (2009).
- Hsu, V. W., Prekeris, R.
- Taguchi, T.
- Goldenring, J. R.
- Delevoye, C., Marks, M. S., Raposo, G. Lysosome-related organelles as functional adaptations of the endolysosomal system. Current Opinion in Cell Biology. 59, 147-158 (2019).
- Hsu, V. W., Bai, M., Li, J. Getting active: protein sorting in endocytic recycling. Nature Reviews in Molecular Cell Biology. 13 (5), 323-328 (2012).
- Desfougeres, Y., D'Agostino, M., Mayer, A. A modular tethering complex for endosomal recycling. Nature Cell Biology. 17 (5), 540-541 (2015).
- Le, L., Sires-Campos, J., Raposo, G., Delevoye, C., Marks, M. S. Melanosome biogenesis in the pigmentation of mammalian skin. Integrated Computational Biology. 61 (4), 1517-1545 (2021).
