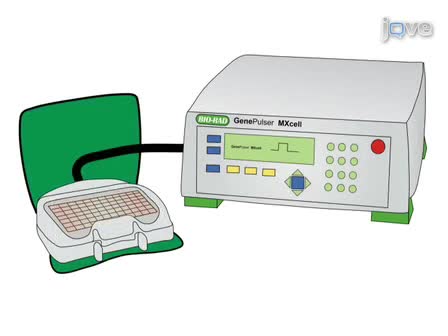
Osteoclasts are tissue-specific macrophage polykaryons derived from the monocyte-macrophage lineage of hematopoietic stem cells. This protocol describes how to isolate bone marrow cells so that large quantities of osteoclasts are obtained while reducing the risk of accidents found in traditional methods.