

Summary
在这个视频文章中,我们目前的分离纯化的方法
Abstract
“
Protocol
果蝇外周神经元的磁性珠排序总评(共完成“议定书”的时间:2.5-3小时)
在任何时候都保持一个干净,核糖核酸酶自由的环境标准的实验室程序必须遵守,以防止RNA降解。
当解剖果蝇幼虫角质层,并放置在细胞内分解缓冲区,外周神经元是脱离角质层的细胞之一。我们必须利用这个属性,并设计了这个协议,以去除大部分从非特异性的细胞,如肌肉和脂肪隔离DA能神经元前角质层。
通过实践,可以成功地完成了整个协议,大约2.5小时内。在实验开始前,必须完成编制抗体包被的磁珠。
1。准备装订细胞磁珠:
实验开始前,必须完成这一步。可以准备准备珠和储存在4℃直至需要。
- 100μL磁珠M - 280链霉亲和素包被的磁珠在PBS的3倍,由500μL新鲜PBS resuspending制粒在一个强大的磁场,它每次清洗。
- 最后直接在未经稀释的生物素标记的鼠抗鼠CD8a抗体(抗体浓度为100微克/毫升)100μL重悬磁珠。
- 孵育1小时偶有轻度振荡,以防止沉淀冰的混合物。 [1μL磁珠M - 280可绑定0.05-0.10微克生物素抗体]。
- 珠抗体混合物洗净三次步骤1.1中所述,以除去多余的抗体。现在涂有抗体的磁珠,并随时可以使用。商店珠抗体混合物在100μL的1X PBS在4 ° C,直到使用。
2。选择和洗涤幼虫:(10-15分钟)
- 挑选30-50年龄匹配的三龄幼虫,并放置在1.6毫升离心管1-1.2毫升1X磷酸盐缓冲液(PBS)。 (3-5分钟)
- 关闭管各1秒和涡最大设置三次。
- 使用火抛光的巴斯德吸管完全弃上清。重复洗(2.1),涡街(2.2)步骤3-4次,直到上清液明显清晰的任何食物颗粒和碎片。
- 简单重复1毫升70%的乙醇洗,弃去上清液。
- 用1毫升的ddH2O洗两次幼虫和上清液。
- 简单重复洗一次1毫升的RNase的上清液。
- 洗净的幼虫在1毫升的ddH2O三次,以确保彻底清除核糖核酸酶的。
3。解剖:(10-12分钟)
- 广场10-12幼虫上的一个Sylgard涂层35毫米的Petri板中心。位置稍微远离对方。 [ 关键的一步:放弃任何,似乎没有要在适当的发育阶段的幼虫]
- 使用所有的幼虫幼虫一双细清扫剪刀剖开前提示。
- 使用一对沉闷杜蒙反转幼虫内而外的第5号钳。幼虫角质层内插入之一forcep所有后结束。捏钳提示一起抢的角质层(图1a)(尝试按Sylgard表面的角质层,使其更容易)后结束。使用产钳的第二对,推动内而外的幼虫角质层。 [ 关键的一步:尝试练习这种方法,细胞的分离实验,然后再尝试几次。设法得到完全倒置的幼虫,以确保所有的软组织暴露容易分解的解决方案。]
- 解剖3-4幼虫后,他们立即转移到新鲜的,冰冷的PBS(管放在冰)在1.6 ml离心管。
- 重复步骤3.1至3.4,直到所有需要的幼虫收集(30-40幼虫在本议定书)。 [ 关键的一步:预计10-20%的亏损,在解剖和离解,并计划据此]
4。去除松散附着的非特异性细胞:(2-3分钟)
[这一步艾滋病松散贴壁的非特异性的组织,如脂肪机构和中枢神经系统的结算。]
- 取1.6 ml离心管中倒幼虫表皮及更换约700-800新鲜冰冷的PBS液的上清液。
- 脉冲涡流离心管5倍全速每脉冲(3秒)。
- 弃上清,约700-800μL新鲜,冰冷的PBS代替。
- 重复步骤4.2和4.3的三倍。
- RESuspend幼虫表皮在400μL新鲜的,冰冷的PBS
5。离解成单细胞悬液,组织:(18-20分钟)
[ 关键的一步:“过度分离可能导致导致不良的细胞产量和细胞存活率低的细胞表面标记的损失。幼虫组织可以分离,无论是机械分离(超声,douncing),酶解(胰蛋白酶,胶原酶等)或两者的结合。由于这些幼虫组织难以解离,我们发现,机械和酶解相结合产生了最好的结果。]
- 添加1.5μL1X Liberase Blendzyme 3(28瓦nsch单位/瓶)400的PBS液中悬浮的幼虫角质层。
- 涡的解决方案为1秒的最大设定值(这应该到解决方案中删除从角质层松散贴壁细胞)每2-3倍。
- 孵育5分钟,在室温(22-25℃)的解决方案。 [ 关键的一步:孵化时间极大地影响了最终的细胞分选效率。建议的孵化时间应足以松动组织。不要超过15分钟。]
- 脉冲涡流管,最大设置为2秒每脉冲全速20-30倍。这应该释放到溶液中的肌肉和其他组织。检查一个小样本,从一个荧光立体显微镜下,在每一步的解决方案。 (有了经验,应该能够确定的解离程度,直接启用的荧光立体显微镜下观察离心管)
- 幼虫表皮洗净,用新鲜的,冰冷的PBS的2-3倍,并终于重悬于500μL新鲜的,冰冷的PBS含1%BSA。
- 为了避免幼虫坚持2毫升Kontes组织磨床和大间隙杵,预涂层组织磨床和杵在1%BSA的PBS液和经过短暂的冲洗丢弃BSA溶液的玻璃表面的角质层。随后,使用火抛光的巴斯德吸管,转让步骤5.5角质层BSA的涂层组织磨床。 [ 关键的一步:预冷组织磨床/上放置了几分钟的冰,以防止细胞损伤/裂解杵] 。
- Dounce与缓慢而稳定的笔触的组织,避免起泡(约20-30杆)。 [ 关键的一步:Dounce缓慢和稳定,否则细胞裂解]
- 为了评估细胞分解水平,擦拭用干净的Kimwipe组织的组织磨床的外墙,和一种荧光立体显微镜下检查。应该有脱离的神经元的角质层,和解决方案中可以看出。如果发现这是很难,或者吸管出一个解决方案的小样本,和一种荧光立体显微镜下观察。细胞分解的一个很好的迹象是从幼虫角质层的神经元的情况下。但是,如果仍然观察连接到角质层的神经元,或观察不完全分离的细胞,进一步dounce,待细胞实现单细胞悬液。
- 磨碎的解决方案,用火抛光的巴斯德吸管5倍缩小到标准的尖端直径约50%,其次是用火抛光的巴斯德吸管10倍缩小到标准的尖端直径约25%。 [ 关键的一步:有力trituration可能会损坏细胞。监控步骤之间的细胞,并调整相应的程序]。
- 通过一个30微米的细胞过滤器的过滤解决方案,并在1.6 ml离心管收集细胞滤液。最终的解决方案应该由一个单细胞悬液,现在准备磁性细胞分选。
6。磁珠细胞分选:(45 - 75分钟,依赖于抗体孵育时间)
- 加入15μl抗体包被的磁珠的细胞悬液500μL(步长5.10)。余下的抗体偶联的磁珠可存储,直到随后的细胞隔离需要。
- 抗体包被的磁珠细胞孵育冰偶尔手工搅拌30-60分钟。 关键的一步:在较高的温度或较长时间的潜伏期,可能会导致非特异性抗体结合。
- 2分钟的离心管中,放置在一个强大的磁场。正选择以及未绑定的珠所有细胞区分开来管方。
- 慢慢地移液管取上清液,确保不打扰细胞沉淀。
- 新鲜的,冰冷的PBS清洗细胞3-4次,以消除任何剩余的非特异性细胞。
- 在30μL新鲜的,冰冷的PBS重悬细胞。
- 近似的细胞的纯度和产量,吸取5μL的细胞悬液ension一个血球的表面抛光和一种荧光立体显微镜下可见荧光细胞计数。同时检查的非荧光的细胞和杂质的任何迹象量。通常情况下,样本将被高度浓缩的荧光细胞。
7。磁性微球的排序细胞RNA的提取:(60 - 75分钟)
- 计数,颗粒在磁场的细胞后,弃去上清液,并添加PicoPure™RNA分离试剂盒(Molecular Devices公司)20μL提取缓冲液。根据细胞数量,可能需要更高的提取缓冲液中添加的量。
- 旋涡管的最大速度,使细胞沉淀提取液混合。
- 30分钟,42℃孵育管。
- 为了确保搬迁前柱纯化的RNA(见第7.5步)的磁珠,简要离心管是在2000(X)G为2分钟沉淀磁珠。管,然后放置在一个强大的磁场,保留沉淀,上清液转移到一个新的离心管。
- 提取和柱纯化的RNA PicoPure RNA提取试剂盒制造商的指示。 DNA酶处理是可选的,可以列上的RNA纯化过程中,根据分析要求。最后,洗脱,直到准备使用绑定在一个小体积的洗脱缓冲液并储存于-80 ° C(11-30μL)的总RNA。如果需要一个1μL等份可用于评估一个2100生物分析仪(安捷伦科技公司)的总RNA质量。
代表性的成果:
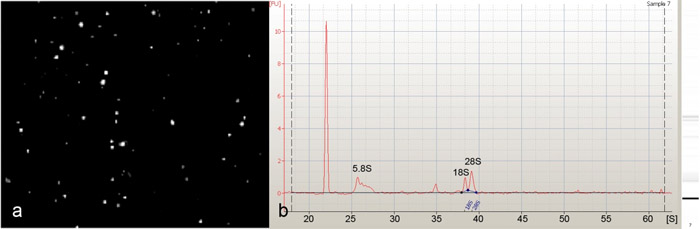
磁珠排序是用来隔离果蝇 DA能神经元(图1)。从这些孤立的DA能神经元(图2a)纯化的RNA被发现的优秀品质急剧5.8S,18S和28S核糖体RNA峰的存在表明,在Agilent 2100生物分析仪(安捷伦科技公司)进行分析时,(图2b)。 30-40第三龄幼虫开始,我们平均300-500类IV DA 使用PPK - GAL4驱动神经元和1500-2000 DA能神经元(I类,II,III及IV)使用隔离的泛DA神经元的具体GAL4 21-7驱动程序。为了评估特定的浓缩,分离的细胞神经元,我们进行了定量逆转录聚合酶链反应(QRT - PCR),使用两个神经元的基因特异性标记( elav和futsch)。这些分析显示显着的两个标记基因相比,通过使用我们的协议(图3)流动表明DA能神经元的一个非常具体的浓缩倍浓缩。最后,来自泛DA能神经元和第四类DA能神经元的分离出的RNA是用于执行安捷伦果蝇全基因组寡核苷酸芯片(4 × 44K)(图4)的转录表达谱。这些分析确定了另外一个以前未知的分子和公认的信号转导通路可能DA神经元的发展发挥重要的职能作用的广谱的DA神经元枝晶形态发生许多以前牵连的监管机构。目前正在进行研究,设计,评估的潜在作用,这些以前未知的分子(S)在调解DA神经元的发展,特别是枝晶的形态发生。

图1: 果蝇 DA能神经元的磁性珠排序示意图(一 )年龄匹配的第三龄幼虫轴承的DA神经元特异性GAL4,记者UAS - mCD8 - GFP转基因是由反相幼虫角质层内而外,揭露解剖PNS的解离缓冲区,并存储在冰冷的PBS(二 )进行酶解加入Liberase Blendzyme 3的溶液中含有幼虫的角质层(三)幼虫组织进一步震荡,trituration分离和douncing除去非特异性标记的组织,如脂肪组织,内脏和中枢神经系统(D,E)细胞,然后使用30微米的细胞过滤器过滤。该解决方案包含了不同类型的细胞,包括上皮细胞,肌肉和神经元的单细胞悬液。,(F)抗鼠CD8a抗体涂层磁珠M - 280加入细胞悬液,冰浴30-60分钟。( G)的磁珠结合的DA能神经元表达的标记GFP融合蛋白(H,I)的磁珠涂层细胞是由放置在一个强大的磁场解决方案分离的CD8 +鼠标。上清液丢弃,细胞洗3次,以除去残留的非特异性的细胞,从而导致( J)Highly纯化DA能神经元的数量。请点击这里看到图1的放大版本。

图2:(一)积极选择,绿色荧光蛋白荧光类IV DA神经元细胞分离和磁珠排序隔离的形象代表。导致神经元的人口被确定为高度富集类IV DA能神经元很少或没有污染的细胞杂质。隔离磁珠排序的大神经元(B),安捷伦2100生物分析仪(安捷伦科技公司)的总RNA电泳,表现出一个优秀的总RNA质量存在5.8S,18S,28S rRNAs。请点击这里看大图图2。

图3:QRT - PCR分析神经元的标记基因的表达在孤立的DA能神经元(GAL421 - 7,UAS - mCD8 - GFP),并通过一小部分的流量是一式三份。 QRT - PCR的两个特定神经元的标记基因(elav和futsch)的表达水平进行了评估。从这些分析中得到的值进行标准化的内对照(rp49),和相对流量的一小部分通过观察这些水平计算ΔΔCτ 方法 6 。 elav和futsch都极大地丰富DA神经元中分离出来的人口相比,流动通过分数。

图4:代表类第四大神经元特异性Cy3标记,标记的微阵列图像文件。这里展示的是安捷伦的果蝇全基因组寡核苷酸芯片(4 × 44K)与Cy3标记的labeld类第四大经磁珠排序纯化的神经元分离的总RNA杂交。图4看到一个更大的版本, 请点击这里。
Subscription Required. Please recommend JoVE to your librarian.
Discussion
这里介绍的协议优化,坚持紧紧地使用磁珠细胞分选策略果蝇三龄幼虫角质层内表面的外周神经元的分离和纯化。虽然我们已经使用了此协议专门隔离果蝇DA能神经元,此协议的应用到其他类型的细胞,角质层中的幼虫或蛹的发展阶段,坚持以隔离(如上皮细胞,肌肉,其他的外周神经元)可适应由不同的几个参数,并使用不同的GAL4,记者UAS - mCD8 - GFP转基因标签的细胞类型或类型的利益。此外,该协议可用于损俱损的功能和增益功能的方法,有兴趣的基因可能会变成一个UAS mCD8 - GFP的转基因可以与GAL4转基因加上直接或者克隆的基因具体亏损的功能( 如无人机的RNAi)或增益功能利益的细胞类型。例如,在一个转录因子的情况下,不妨查明后可能向上或向下调节基因功能丧失或增益功能利益的细胞类型中的表达。分离总RNA纯化的细胞类型的兴趣,通过此协议,并利用这种RNA进行微阵列表达分析可以找出差异调控基因,可能代表下游的转录调控目标,发挥的作用,在介导细胞内的表型变化。
对于成功的细胞分选,它是必不可少的,给予认真的关注上述协议中所强调的关键步骤。常见的问题领域,可能需要一些进一步的故障诊断和优化的例子,取决于细胞类型,包括:(1)低格的产量和(2)细胞聚集在磁珠隔离。在第一种情况,可以尝试Liberase Blendzyme 3浓度的降低,通过增加通过douncing机械分离和补偿。在第二种情况下,人们可以尝试减少应用胶粘剂实验室带一个或多个层以上的磁铁的磁场强度。
Subscription Required. Please recommend JoVE to your librarian.
Acknowledgments
我们感谢博士。裕农一月和韦斯Grueber提供在这项研究中所使用的飞股票。作者承认托马斯F和支持这项研究的凯特米勒Jeffress纪念信托(DNC)和乔治梅森大学教务办公室(EPRI)。
Materials
| Name | Company | Catalog Number | Comments |
| 10X Phosphate Buffered Saline (PBS) | MP Biomedicals | PBS10X02 | Diluted to 1X working solution |
| 10X Liberase Blendzyme 3 | Roche Group | 11814176001 | Diluted to 1X working solution (28 Wünsch units/vial) |
| RNase-AWAY | Sigma-Aldrich | 83931 | |
| Biotinylated Rat anti-Mouse-CD8a antibody | Invitrogen | MCD0815 | 100 μg/ml stock concentration |
| BSA (Bovine Serum Albumin), Fraction V | GIBCO, by Life Technologies | 11018-017 | Prepare a 1% BSA solution in PBS |
| Dynabeads M-280 Streptavidin | Invitrogen | 11205D | 1 μl can bind 0.05-0.10 μg of biotinylated antibody |
| PicoPure RNA Isolation Kit | Molecular Devices | KIT0204 | Follow manufacturer’s instructions |
Equipment
|
|||
References
- Corty, M. M., Matthews, B. J., Grueber, W. B. Molecules and mechanisms of dendrite development in Drosophila. Development. 136, 1049-1061 (2009).
- Parrish, J. Z., Emoto, K., Kim,, Jan, Y. N. Mechanisms that regulate establishment, maintenance, and remodeling of dendritic fields. Ann. Rev. Neurosci. 30, 399-423 (2007).
- Lee, T., Luo, L. Mosaic analysis with a repressible cell marker for studies of gene function in neuronal morphogenesis. Neuron. 22, 451-461 (1999).
- Grueber, W. B. Projections of Drosophila multidendritic neurons in the central nervous system: links with peripheral dendrite morphology. Development. 134, 55-64 (2007).
- Song, W., Onishi, M., Jan, L. Y., Jan, Y. N. Peripheral multidendritic sensory neurons are necessary for rhythmic locomotion behavior in Drosophila larvae. Proc Natl Acad Sci USA. 104, 5199-5204 (2007).
- Livak, K. J., Schmittgen, T. D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) methods. Methods. 25, 402-408 (2001).

{kind=link}
{kind=link}
{kind=link}